胡桃楸种源家系幼龄期生长变异及选择研究
2022-03-26芦贤博徐连峰庞忠义刘继锋温宝阳裴晓娜王学刚赵曦阳
芦贤博,徐连峰,庞忠义,刘继锋,温宝阳,裴晓娜,赵 岭,王学刚,赵曦阳*
(1.林木遗传育种国家重点实验室东北林业大学,黑龙江 哈尔滨 150040;2.黑龙江省林科院齐齐哈尔分院,黑龙江 齐齐哈尔 161005;3.国有新民市机械林场,辽宁 沈阳 110300;4.宾县万人欢林场,黑龙江 哈尔滨 150411;5.国家林业和草原局哈尔滨林业机械研究所,黑龙江 哈尔滨,150086;6.中国林科院寒温带林业研究中心,黑龙江 哈尔滨,150086)
胡桃楸(Juglans mandshuricaMaxim.)为胡桃科(Juglandaceae)胡桃属(Juglans)落叶乔木。在我国主要分布于东北地区的小兴安岭、完达山脉、长白山区和辽宁东部[1],是我国Ⅱ 级珍稀树种和中国珍稀濒危树种的三级保护植物[2],与黄菠萝、水曲柳并称为“东北三大硬阔”。胡桃楸树干通直圆满,材质坚硬致密,木材弹性好、易加工,被广泛用于军工、船舶、家具、居室装修及乐器制造等方面,是优良的用材树种[3];其果实果仁营养丰富,含油量高,可制作油料和保健食品,也可作为一些特色食品的添加剂原料;胡桃楸还具有一定的药用价值,其青果、枝皮及种仁均可入药[4-5];并且,其新鲜根皮、枝皮、青果皮中均含有药用价值较高的化学物质——胡桃醌[6],具有极强的抗菌[7-8]及抗肿瘤作用[9],目前在临床医学及医药化学领域中得到广泛应用。
林木幼龄期选择是根据林木优良性状在早、晚龄间存在的正相关关系,针对其幼龄期的性状表现,进行优良个体选择[10]。自20 世纪80 年代以来,国内外学者已对多个树种进行幼龄期选择研究,并取得重大进展[11]。在胡桃楸方面,刘桂丰等[12]和颜廷武等[13]的研究表明,幼龄期的胡桃楸已具有代表性,因此,对胡桃楸进行幼龄期选择是可行且有效的。育种专家已经对胡桃楸进行种源区划,并对少数种源、家系进行初步评价[14]。但是存在选育的良种少,繁殖系数低,很难满足东北林区造林需求等问题,并且由于胡桃楸用途广泛,导致人为的过度采伐,野生胡桃楸资源遭到严重破坏,近年来在林业生产上虽然对胡桃楸进行了大面积人工栽培,但多利用现有的普通胡桃楸造林,在生长和木材产量方面仅表现出较低的增益,因此,胡桃楸的良种选育工作意义重大,不同生态区、不同用途良种选育迫在眉睫。本研究以黑龙江省不同种源内的28 个胡桃楸半同胞家系为材料,对其6 年生生长性状的遗传变异进行分析,初步筛选优良亲本、优良家系和单株,为胡桃楸用材林的良种选育提供基础。
1 材料与方法
1.1 试验地点与试验材料
试验地位于黑龙江省宾县万人欢林场(128°04′29″~128°19′13″ E,45°44′20″~45°57′18″ N),该地平均海拔340 m,年平均温度2.4℃,年降水量600~800 mm,无霜期130 d 左右,植物生长期140 d 左右,年日照时数2 560 h。
试验材料包括黑龙江省万人欢、七台河、牡丹江、宝清等4 个种源的28 个胡桃楸半同胞家系,其中,万人欢种源有14 个家系,七台河种源有7 个家系,牡丹江种源有4 个家系,宝清种源有3 个家系。不同种源的地理气象因子概况见表1。2015 年春季育苗,2016 年定植。采用随机完全区组试验设计,3次重复,12株小区,株行距为3 m × 4 m,周围设置保护行。

表1 胡桃楸不同种源地理气象因子Table 1 The geographical and meteorological factors of different J.mandshurica provenances
1.2 试验方法
于2020 年11 月对6 年生的胡桃楸半同胞家系的树高、地径、冠幅、分枝角、通直度、侧枝数等性状进行测量。利用塔尺测量树高;利用围尺测量地径;利用钢卷尺测量冠幅(分别测量东西和南北两个方向后取平均值);利用量角器测量分枝角;利用分级法确定通直度[15],具体见表2;利用地径和树高比值计算尖削度计算单株尖削度[16];连续测定近3 年树高生长量,利用平均值计算树高年均生长量。

表2 通直度调查评判标准及分值Table 2 The research criteria and score of stem straightness degree
1.3 统计分析方法
本研究所有数据均使用SPSS 22.0 软件和EXCEL 软件进行分析。方差分析线性模型为[17]:

式中:μ 为总体平均值,Bi为 区组效应,P/F为种源/家系效应,Piji/Fiji为区组与种源/家系的交互作用,Mei ji为机误。
家系遗传力(H2)计算公式[18]:

式中:g为 亲本的一般配合力,x为亲本的某个交配组合在某个性状的子代平均值,μ为这个性状所有组合的子代总平均值。
主成分值和综合得分计算公式分别为[25]:

式中:Yi为第i主成分值,αij为主成分i内性状j的特征值,Xj为 性状j的平均值,W为综合得分值,ωi为 第i主成分贡献率,n为性状个数,p为提取主成分个数。
2 结果与分析
2.1 各性状方差分析
对胡桃楸各种源及各家系不同生长性状进行方差分析,结果见表3、4。除侧枝数外,其余各性状在种源间的差异均达到显著水平(p< 0.05);通直度、侧枝数和尖削度在区组间以及通直度在区组 × 种源间的差异未达显著水平(p> 0.05);分枝角在种源间和侧枝数、尖削度在区组 × 种源间的差异显著(0.01 <p< 0.05);除此外,各性状在其他变异来源间的差异均达到极显著水平(p<0.01)。不同家系各性状方差分析表明:除通直度、侧枝数及尖削度等性状在区组间的差异不显著外(p> 0.05),各性状在各变异来源间的差异均达显著水平(p< 0.05)。因此,胡桃楸的生长性状在种源及家系间具有较大遗传差异,对优良种源及优良家系的选择具有较大改良潜力。

表3 不同种源各性状方差分析表Table 3 Variance analysis of different traits among provenances

表4 不同家系各性状方差分析表Table 4 Variance analysis of different traits among families
2.2 各性状遗传变异分析
胡桃楸28 个家系各生长性状遗传变异参数见表5。所有家系树高平均值为2.565 m,变幅为2.042~3.046 m;树高年增长量的平均值为0.770 m,变幅为0.598~0.931 m;地径的平均值为6.135 cm,变幅为4.772~7.439 cm;冠幅的平均值为1.776 m,变幅为1.206~2.263 m;分枝角平均值为50.129°,变幅为44.792°~57.833°;通直度平均值为3.816,变幅为2.972~4.278;侧枝数平均值为2.356 个,变幅为1.810~2.704 个;尖削度平均值为2.411,变幅为2.209~2.667。

表5 不同家系各性状变异参数Table 5 Variation parameters of different traits families
各性状表型变异系数变化范围为14.049%~41.519%,除尖削度外,其余各性状表型变异系数均超过20%。各性状遗传变异系数变化范围为8.754%~23.873%,出尖削度外,其余各性状遗传变异系数均超过10%。从家系遗传力来看,除通直度外,其余各性状的家系遗传力较高;从单株遗传力来看,除冠幅、分枝角、侧枝数和尖削度外,其余各性状的单株遗传力较高。
2.3 各性状相关性分析
对所有家系生长性状进行相关性分析,结果见表6。各性状中,除尖削度与分枝角(-0.056)、尖削度与侧枝数(0.049)以及通直度与冠幅(-0.009)之间的相关性不显著外(p> 0.05),其余各性状之间均达显著水平(p< 0.05),其中,树高与树高年增长量以及树高与地径之间相关性较高,相关系数均在0.80 以上。树高与尖削度之间呈极显著负相关,相关系数为-0.243。此外,尖削度与树高年增长量、通直度之间均达极显著负相关,相关系数分别为-0.292、-0.131。

表6 不同家系各性状相关性分析Table 6 Correlation coefficients among different traits among families
2.4 各性状一般配合力分析
28 个胡桃楸半同胞家系各性状一般配合力见表7、8。树高一般配合力变化范围为-0.523~0.481,其中,较高的家系为WRH2、WRH3 和QTH8;树高年增长量一般配合力变化范围为-0.173~0.161,其中,较高的家系为WRH2、WRH3和WRH13;地径一般配合力变化范围为-1.363~1.304,其中,较高的家系为QTH2、WRH3 和WRH6;冠幅一般配合力变化范围为 -0.569~0.487,其中,较高的家系为QTH2、BQ3 和QTH8;分枝角一般配合力变化范围为 -5.239~7.803,其中,较高的家系为WRH5、WRH14和WRH3;通直度一般配合力变化范围为-0.844~0.461,其中,较高的家系为WRH8、WRH5 和QTH1;侧枝数一般配合力变化范围为-0.546~0.348,其中,较高的家系WRH2、WRH13 和MDJ3;尖削度一般配合力变化范围为-0.202~0.256,其中,较高的家系为MDJ7、WRH6和QTH10。

表7 树高、树高年增长量、地径、冠幅的一般配合力Table 7 General combining ability values of tree height,annual height increase,basal diameter and crown diameter
2.5 各性状主成分分析
对胡桃楸的各性状进行主成分分析,分析结果见表9。以特征值大于1 为标准提取出3 个主成分,累计贡献率达到73.264%,说明选出来的3 个主成分可以涵盖所有性状73.264%的信息,具有高度概括的效果,故选取前3 个主成分进行分析。
由表9 可知:主成分Ⅰ 中,树高(0.956)、树高年增长量(0.878)、地径(0.892)及冠幅(0.824)分别在3 个主成分中的特征值最高,故主成分Ⅰ 可以代表这4 个性状。主成分Ⅱ 中,尖削度(0.879)在3 个主成分中的特征值最高,故主成分Ⅱ 可以代表该性状。在主成分Ⅲ 中,分枝角(0.622)、通直度(0.482)、侧枝数(0.526)在3 个主成分中的特征值最高,故主成分Ⅲ 可以代表这3 个性状。
由表9 中各性状的载荷量可以得到主成分的线性方程,用X1、X2、X3、X4、X5、X6、X7、X8分别表示树高、树高年增长量、地径、冠幅、分枝角、通直度、侧枝数、尖削度;用Y1、Y2、Y3分别表示主成分Ⅰ、Ⅱ、Ⅲ,得到如下线性方程:

2.6 各种源综合得分
由于各主成分的贡献率不同,因此,选取它们的贡献率作为计算综合得分的权重,由表9 可知:这3 个主成分的权重分别为:42.398%、17.219%、13.646%。综合评价得分计算公式为:
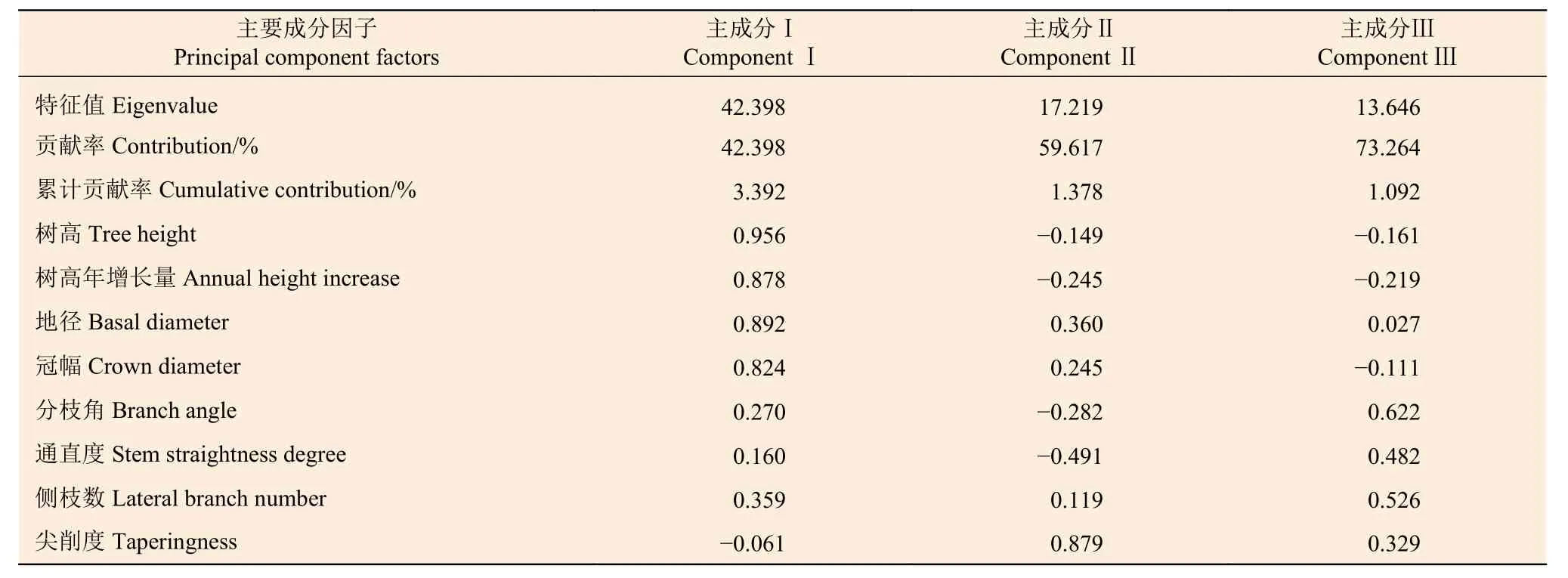
表9 各性状主成分分析Table 9 Principal component analysis of different traits

经过计算后得到胡桃楸各种源的综合得分以及排名情况见表10。各种源中,综合得分变化范围为12.362~13.319,按树高、树高年增长量和地径现实增益在2%以上,兼顾其他性状对种源进行评价选择,万人欢种源入选为优良种源,其树高、树高年均增长量、地径及冠幅现实增益分别为3.109%、5.014%、2.193%和1.040%。
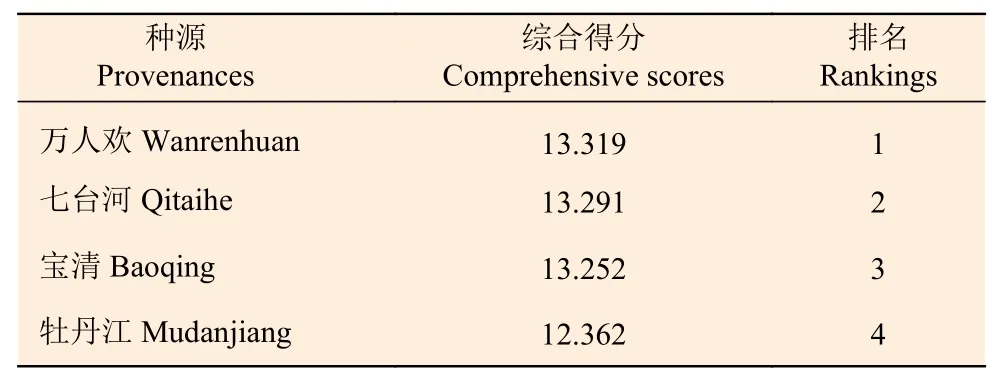
表10 各种源综合得分和排名Table 10 Comprehensive score and ranking of provenances

表8 分枝角、通直度、侧枝数、尖削度的一般配合力Table 8 General combining ability values of branch angle,stem straightness degree,lateral branch number and taperingness
2.7 优良家系及单株综合得分
根据主成分分析的结果,对所有家系及单株进行综合得分计算,根据各家系及各单株综合得分情况,以10% 和1% 的入选率,分别对家系与单株进行筛选,共获得3 个优良家系(WRH3、WRH5、QTH2)与10 株优良单株,入选家系及单株综合得分及排名情况见表11、12。入选家系树高、树高年增长量、地径及冠幅的平均值分别为2.835 m、0.854 m、7.033 cm 和1.938 m,遗传增益分别为5.835%、5.410%、7.908%和6.069%;入选优良单株树高、树高年增长量、地径及冠幅平均值比总体平均值分别高1.068 m、0.250 m、2.225 cm 和1.312 m,单株遗传增益分别为26.74%、24.11%、23.91%和26.53%。

表11 优良家系综合得分和排名Table 11 Comprehensive score and ranking of excellent families
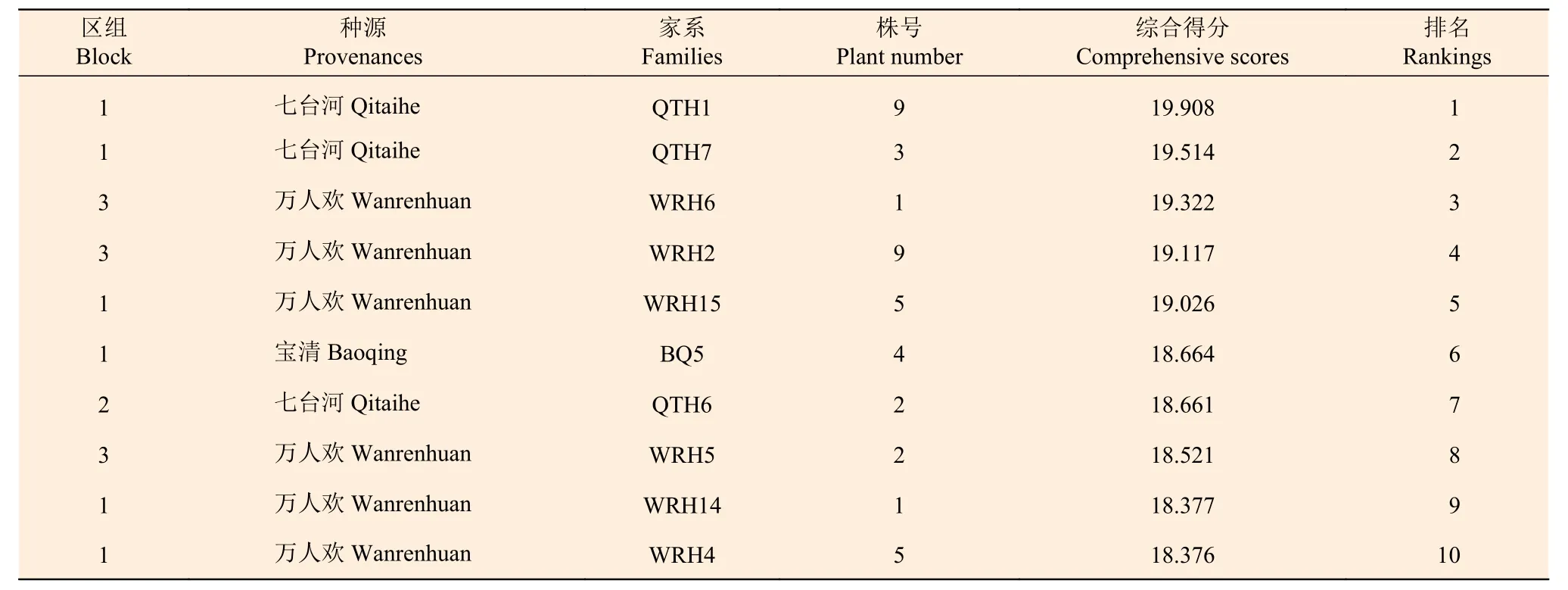
表12 优良单株综合得分和排名Table 12 Comprehensive score and ranking of excellent individual plants
3 讨论
遗传和变异是林木育种工作的主要研究内容[26],对变异来源、特点以及规律的认识是进行林木良种选择工作的重要基础[27]。本研究通过对不同性状种源间及家系间差异分析发现,各性状在胡桃楸种源间以及家系间均存在丰富的遗传变异,这与褚宪丽等[28]对胡桃楸3 个种源、45 个家系的分析结果一致,表明不同种源、半同胞家系间各生长指标间存在较大差异,有利于优良家系与单株的选择。
变异系数的大小反映植物的变异程度,它能体现林木各性状的遗传变异能力,包括遗传变异系数和表型变异系数[21]。遗传变异系数越大,表明该家系在该性状中可被利用的改良潜力越大[29]。本研究中,各性状的表型变异系数变化范围为14.049%~41.519%,与袁显磊等[30]、周彦超[31]对胡桃楸的研究结果相近,变化幅度较大,说明胡桃楸各表型性状间存在丰富的变异。此外,冠幅与侧枝数的表型变异系数最高,表明以冠幅与侧枝数作为选优指标的潜力更大。树高、树高年增长量以及地径3 个性状的遗传变异系数占表型变异系数的比例较大,均超过了70%,与解懿妮等[32]对桉树(Eucalyptus S.T.Blake)的研究结果相似,说明胡桃楸家系间的变异受遗传因素的影响较大。遗传力表示亲本某一性状遗传给子代的能力,遗传力的高低表明该性状稳定遗传给后代受到环境影响的大小[33]。本研究中,28 个胡桃楸半同胞家系各性状遗传力变化范围为0.284~0.797,除通直度外,各性状遗传力均超过0.45,属于中等遗传力水平[34],与翟文继等[35]对楸树(Catalpa bungeiC.A.Mey.)的研究结果相似,表明不同家系的胡桃楸生长性状受到了环境与遗传的综合控制;其中,侧枝数与分枝角的遗传力最高,说明侧枝数与分枝角在胡桃楸家系间的优良表现受环境影响较小,能够稳定遗传给子代。
相关性分析可以反映各性状之间存在的联系,对了解不同性状之间的关系具有重要作用[36-37]。本研究中,从各性状的相关性分析可以看出,胡桃楸各性状之间的相关性较高,其中,树高、树高年增长量、地径、冠幅以及分枝角等性状间均达极显著正相关;而树高与地径决定树木长势情况,树高与地径之间达极显著正相关,且相关系数较高,为0.827,与赵曦阳等[38]对白杨派杨树无性系的研究结果相似,研究结果为长势优良的胡桃楸综合评价选择提供依据。
一般配合力指在一个杂交群体中,某个亲本杂交组合的子代平均值与子代总平均值的离差[39]。一般配合力通常被认为是选择理想亲本和发展优良杂交的先决条件[40]。本研究对28 个半同胞家系各性状的一般配合力进行了分析,以20% 的入选率,各性状均筛选出6 个亲本,根据树高、树高年增长量与地径作为木材产量的指标,综合筛选出4 个优良亲本(WRH2、WRH3、WRH15 和QTH2);根据冠幅、分枝角以及侧枝数(注,各指标取所得参数相反数进行筛选)作为种植密度的指标,综合筛选出2 个优良亲本(WRH、QTH3);根据通直度与尖削度作为干形的指标,综合筛选出3 个优良亲本(QTH1、QTH2 和WRH6)。所选家系的亲本可以作为未来不同育种目标的杂交育种的首选亲本。
主成分分析法既能把握植株的综合性状表现,又能简化选优指标,简化选择程序,克服人为主观误差,选择结果准确、客观[41]。本研究对胡桃楸8 个生长性状进行了主成分分析,提取出3 个主成分,主成分Ⅰ 中树高、树高年均增长量、地径和冠幅特征值较高,可代表生长量相关信息;主成分Ⅱ 中尖削度特征值较高,可代表干形相关信息;主成分Ⅲ 中分枝角和侧枝数平均值特征值较高,可代表分枝相关信息。依据主成分分析结果,筛选出1 个优良种源(万人欢),其树高、树高年均增长量及冠幅现实增益分别为3.109%、5.014%、2.193%和1.040%;筛选出3 个优良家系(WRH3、WRH5、QTH2),入选家系树高、树高年增长量、地径及冠幅等性状遗传增益分别为5.835%、5.410%、7.908%和6.069%。与胡文杰等[42]对9 年生枫香(Liquidambar formosanaHance)的研究结果相似。其中,QTH2 既是优良家系所选材料,又是优良亲本所选材料,表明QTH2 性状表现优良,可进一步作为胡桃楸优良材料用于推广应用。在优良家系选择的同时,还筛选出优良单株,以1%的入选率,共获得10 株优良单株,入选优良单株在树高、树高年增长量、地径及冠幅的平均值比总体平均值分别高1.068 m、0.250 m、2.225 cm 和1.312 m,单株遗传增益分别为26.74%、24.11%、23.91%和26.53%。
4 结论
育种目标决定育种方法,胡桃楸主要作为果材兼用树种,对胡桃楸的评价应从多方面、多角度进行。但因其有性繁殖周期长、扦插和嫁接等无性繁殖成活率低等问题,使得胡桃楸研究进展仍较为缓慢。本研究以胡桃楸良种选育为目的,综合多个性状,结合主成分分析法,对胡桃楸种源、家系及单株进行评价与选择,共筛选出1 个优良种源、3 个优良家系和10 个优良单株,入选材料各性状之间相关性强,遗传力高,种源间以及家系间有较大的选择潜力,所选材料优势明显,在胡桃楸良种选育与推广应用中可重点考虑。
