Comparative Osteology of Two Far Eastern Species of Ratsnakes (Serpentes:Colubridae),Elaphe dione (Pallas,1773) and E.schrenckii (Strauch,1873),for the Purpose of Palaeontological Studies
2022-03-26ViatcheslavRATNIKOV
Viatcheslav RATNIKOV
Voronezh State University,Universitetskaya Pl.,1,Voronezh,Voronezhskaya Oblast 394018,Russian Federation
Abstract The taxonomic identification of Quaternary remains requires a comparison of fossil bones with the skeletons of existing taxa based on the morphological criteria.However,the osteology of most modern snake species has not yet been studied.This work is aimed at finding differences in the bone morphology of Amur and Dione’s ratsnakes and the criteria for their identification.The variety of bone morphology depends on several parameters.First,these are species differences which determine the systematic position of the animal.Second,it is individual variability.Third,these are progressive changes in the vertebrae morphology along the vertebral column from the first vertebra to the last.Fourth,it is age-related variability.20 skeletonised specimens:12 specimens of Elaphe dione and 8 specimens of Elaphe schrenckii were used to define diagnostic osteological characters.The bones of the two studied species are very similar and their variability overlaps.However,differences and identification criteria were found for 13 skull bones:nasal,frontal,parietal,supraoccipital,maxilla,quadrate,palatine,pterygoid,ectopterygoid,basiparasphenoid,basioccipital,dentary and the compound bone.In addition,the variability and differences of the cervical,trunk,cloacal,and caudal vertebrae were examined.
Keywords bones morphology,identification criteria,interspecific differences,intraspecific variation
1.Introduction
The taxonomic identification of Quaternary remains requires a comparison of fossil bones with skeletons of existing taxa based on the morphological criteria.This approach is often affected by a lack of works describing the osteology of modern taxa with due consideration of intraspecific and interspecific morphological variability (Bochatonet al.,2019).
The fossil remains of amphibians and reptiles collected over the past decades in a number of caves in the Russian Far East need to be identified.This will provide information on the taxonomic composition and diversity of ectothermic tetrapods at different stratigraphic levels in different locations of the region.These data can be used to reconstruct the environment (Ratnikov,1996,2016) and its changes over time.As fossil materials contain scattered bones,each cranial and postcranial element has to be determined separately.The study of the collection from the Medvezhyi Clyk Cave in the Partizansky District of Primorsky Krai (about 100,000 bones) revealed the predominance of two snake species that currently live in the area around the cave,namely Dione’s (Elaphe dione) and Amur (Elaphe schrenckii) ratsnakes (Ananjevaet al.,2006;Maslova,2016).Their remains include both skull elements and vertebrae from different parts of the vetebral column.It is obvious that they need to be differentiated to identify their quantitative relationship in the location.
Modern materials have not yet been used to study the osteology ofElaphe dioneandElaphe schrenckii.The exception is their vertebrae (Helfenberger,2001;Ratnikov,2004).The aim of the first study was to estimate the phylogenetic relationships between ratsnake species in the Old World by using,among other things,vertebra features.The information about the vertebrae was given in a table which provided average values for few indices without any descriptions or illustrations.Therefore,this cannot help identify fossil remains.The second publication specified the characteristics of the trunk vertebrae of eight ratsnake species,includingElaphe dioneandElaphe schrenckii.Unfortunately,these descriptions relate to the basic morphology of the trunk vertebrae and do not cover the possible variants for their variability.
Recently,the fossil vertebrae ofElaphecf.dioneandElaphecf.schrenckiihave been described from the Pleistocene deposits of Shanyangzhai Cave,Hebei,China (Chenet al.,2019;Chen,2020).No one else has reported the fossil remains of the Amur ratsnake.On the contrary,information about the findings regarding the Dione’s ratsnake,which now has a large area,is available in publications,but only few of them contain a description of their bone morphology.The vertebrae ofElaphe dionewere described from the early Middle Pleistocene localities of Berezovka (Ratnikov,1998) and Korotoyak-4 (Ratnikov,2020) in Eastern Europe.The vertebrae ofElaphecf.dionewere recorded from the early Middle Pleistocene locality of Kozii Ovrag (Ratnikov,2002a) in Eastern Europe.A pterygoid fragment from the late Pleistocene of the Andronovo locality in Eastern Europe may also belong toElaphe dione(Ratnikov,2018).Vertebrae identified as belonging toElapheaff.dione,were described from the late Middle Miocene localities of Baikadam and Malyi Kalkaman-1 in north-eastern Kazakhstan (Ivanovet al.,2018),and the late Pliocene locality of Kotlovina in Ukraine (Ratnikov,2002b).Dione’s ratsnake was found in a number of caves in Crimea (Zerova and Chkhikvadze,1984),the Volga region (Yakovlevet al.,2013,2016),Southern Urals (Yakovlevet al.,2004;Yakovleva,2004;Yakovleva and Yakovlev,2017),and Altai (Gutieva and Chkhikvadze,1990),as well as on the late Paleolithic site of Listvenka in Krasnoyarsk Krai (Ratnikov,2009;Ratnikovet al.,2009).Due to the absence of comparative characteristics for the skull bones and the large intraspecific variability of the vertebrae,especially inElaphe dione,there is a need for an additional search to determine the criteria for their diagnostics.
The purpose of this work was to find differences in the bone morphology of Amur and Dione’s ratsnakes and the criteria for their identification.
2.Materials and Methods
20 skeletonised specimens (Table 1):12 specimens ofElaphe dioneand 8 specimens ofElaphe schrenckii,were used to define diagnostic osteological characters.The author had been collecting the modern comparative material,including the described specimens for several dozens of years.Some untagged preserved and dried specimens originated from museum collections.The corpses of other specimens were found by herpetologists during fieldwork and were sent to the author ina dried form.The skeletons were obtained from individuals of varying ages and sizes representing the intraspecific variability ofElaphe dioneandElaphe schrenckii.Sexual dimorphism was not taken into consideration,because the sex had only been recorded for some of the specimens.All bones were boiled,separated,and cleaned from the muscles and sinews with a scalpel.To retain their position,the vertebrae were threaded on a string in sequence.Specimens were studied under a binocular microscope.They were included in the author’s comparative osteological collection located at the Faculty of Geology of Voronezh State University.
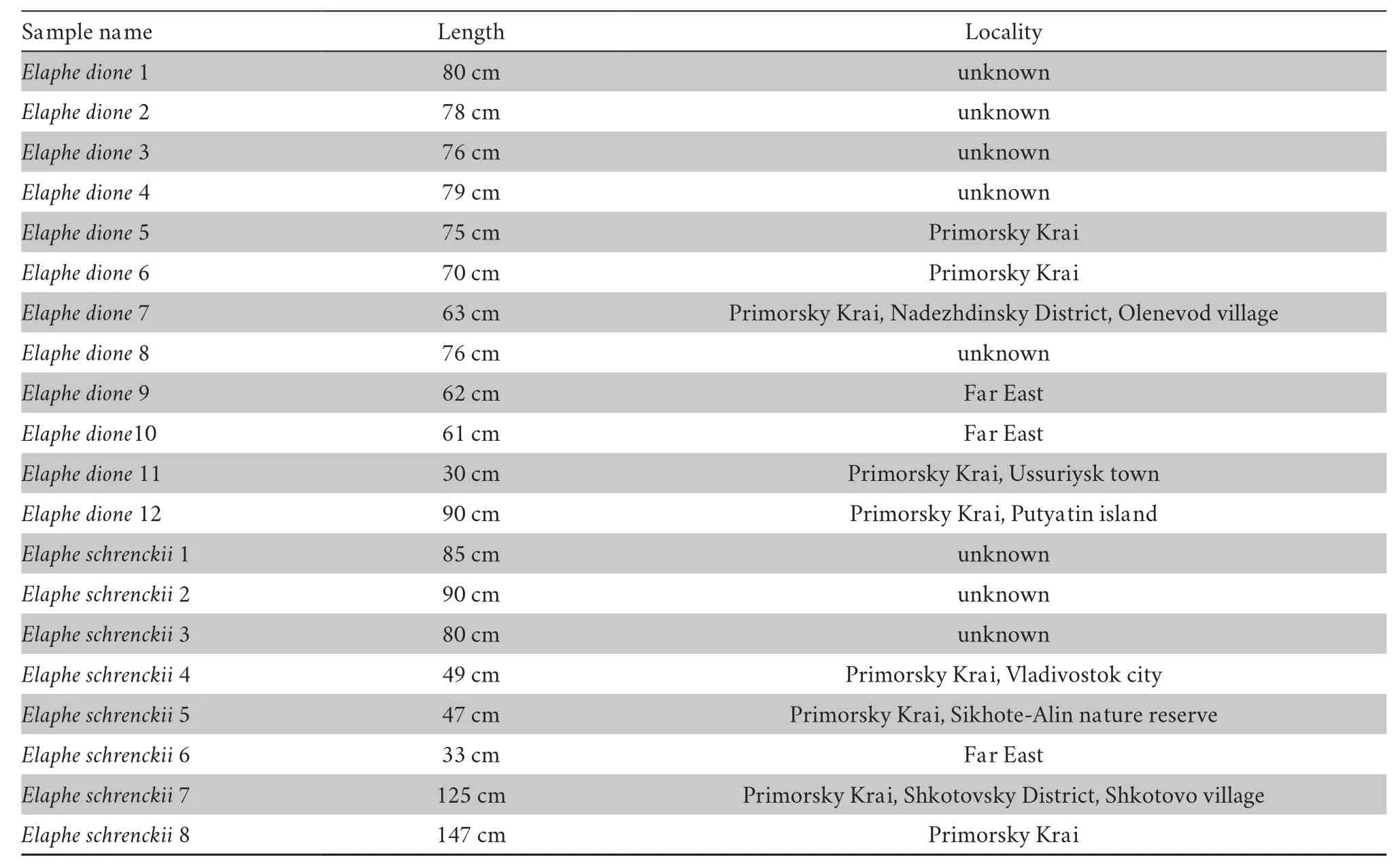
Table 1 Information about the studied specimens of Elaphe dione and Elaphe schrenckii.
First,homologous bones were compared in two randomly selected specimens of roughly equal length,oneElaphe dioneand oneElaphe schrenckii.Then,the revealed difference was checked against the other specimens.It is obvious that not all the differences that were originally found appeared to be species criteria.Many of them only demonstrated variants of intraspecific variability.These variations were shown in the descriptions of the bones.
Digital pictures were taken with a DCM-300 digital camera attached to binoculars.Each bone was photographed several times with different depths of field.The use of the HeliconFocus program made it possible to obtain the resulting sharp images of the bone.Further image processing was performed with the help of PhotoShop and CorelDRAW.
In the figures of vertebrae,different capital letters indicate different specimens,while identical capital letters with different numbers (i.e.,A1,A2,A3) indicate the same specimen,but from different views.In the figures of cranial bones,only capital letters were used.The morphological elements indicated in the figures correspond to the elements mentioned in the text.
The nomenclature given by Szyndlar (1984) and Raccaet al.(2020) was used for descriptions.Some terms have been added by the author.
3.Results
3.1.Description of the cranial bonesThe cranial bones of the two species under consideration are very similar,since they belong to the same genus.The variability of various elements made it difficult to find identification criteria.However,these criteria were found for many (but not for all) bones of the skull.Below,first a description of the general features for each bone will be provided,and then the differences found.
Nasal(Figure 1) The bone consists of two plates perpendicular to each other and a medial nasal flange.The vertical ventromedial plate is called medial nasal lamina.The horizontal dorsal plate is called the dorsolateral nasal lamina.In dorsal view,the dorsolateral nasal lamina consists of two parts:a small anterior nasal lobe and a large lateral nasal lobe.Szyndlar (1984) considered the possibility to correctly identify species by their nasals to be dubious.However,these bones have some differences for the studied species.The anterior nasal lobe inElaphe dioneis comparatively large,while in the case ofElaphe schrenckii,it is very small.The lateral nasal lobe is wider and shorter inElaphe dione.InElaphe dione,its lateral edge is rounded,whereas inElaphe schrenckii,it is comparatively straight.The medial nasal flange inElaphe dioneis narrower.
Frontal(Figure 2) This bone is of cylindrical shape and of moderate length.Its anterior opening (frontal aperture) is oval and is horizontally oriented.The walls of the bone around the anterior opening have three processes:the septomaxillary process ventrally and the internal and external premaxillary processes laterally.The trabecular ridge runs along the medial edge of the ventral side of the bone.In dorsal view,the frontal bone is rectangular in shape with a markedly concave lateral margin.The dorsal surface is convex turning laterally into a relatively wide lateral crest.It starts from the posterior base of the external premaxillary process and is of the same width in ventral view or is slightly wider posteriorly.The posterior edge of the dorsal surface is straight and bevelled forward at an angle of about 15°.
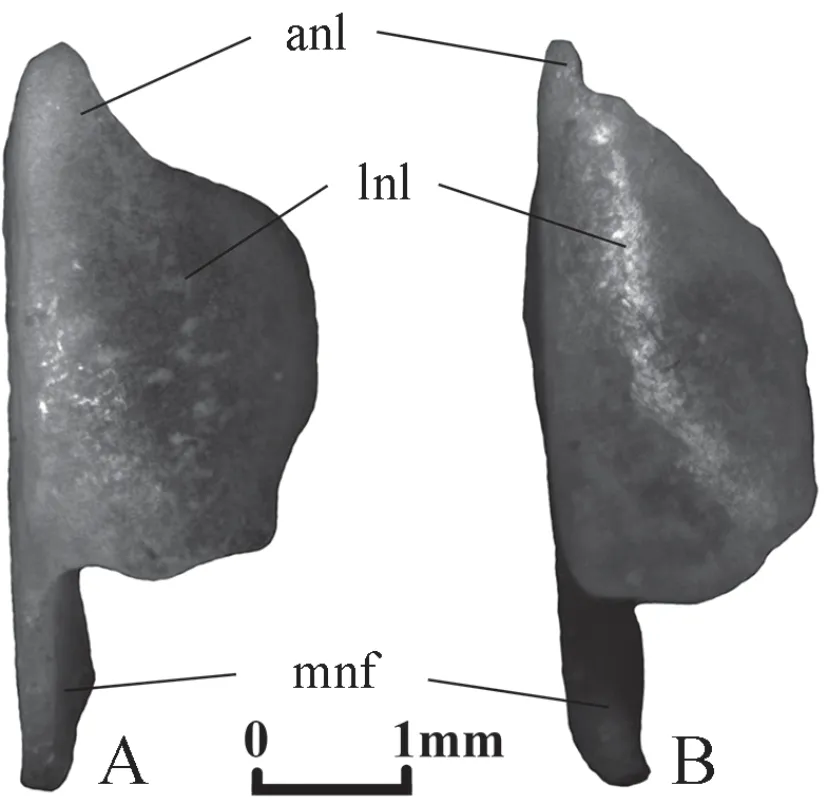
Figure 1 Right nasal in dorsal view.A:Elaphe dione,specimen No.8;B:Elaphe schrenckii,specimen No.3.anl:anterior nasal lobe of dorsolateral nasal lamina;lnl:lateral nasal lobe of dorsolateral nasal lamina;mnf:medial nasal flange.
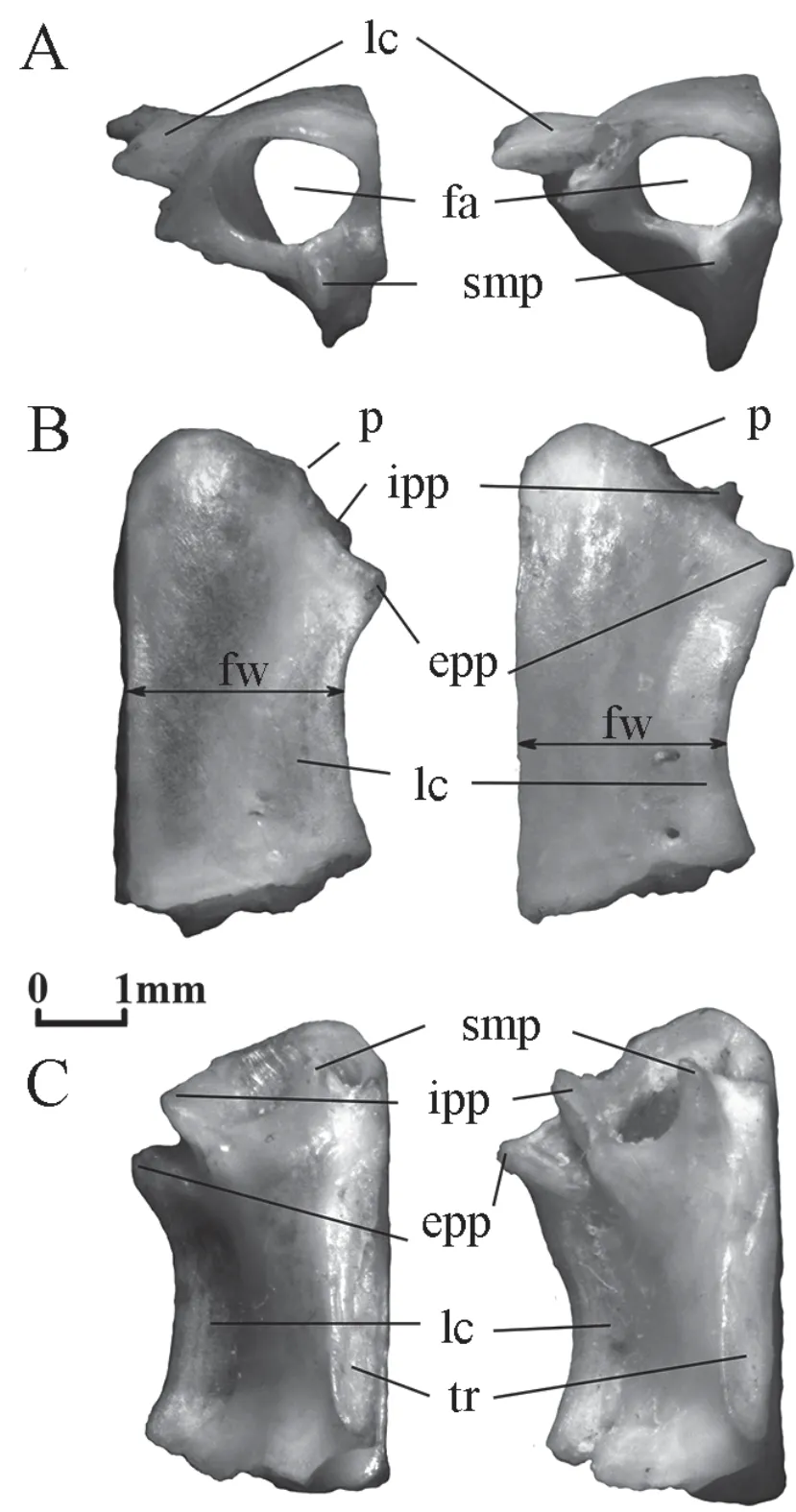
Figure 2 Right frontal of Elaphe dione,specimen No.8 from the left and Elaphe schrenckii,specimen No.3 from the right.A:anterior view;B:dorsal view;C:ventral view.epp:external premaxillary process;fa:frontal aperture;fw:minimal frontal width;ipp:internal premaxillary process;lc:lateral crest;p:protrusion,which divides the anterior edge into the medial (curved) and lateral (straight) parts;smp:septomaxillary process;tr:trabecular ridge.
The width of the bone varies in dorsal view.InElaphe dione,it is the shortest in the front half or in the middle of the bone,while inElaphe schrenckii,it is the shortest in the posterior half or almost in the middle of its length.
Differences between the two species can also be found in the structure of the anterior edge of the bone.A small protrusion,from which the internal premaxillary process begins,divides the anterior edge into the medial (curved) and lateral (straight) parts.The length of both parts inElaphe dioneis the same,while inElaphe schrenckiithe medial part is shorter than the lateral.The anterior point of the bone is located in the middle of the medial part inElaphe schrenckii,and is slightly displaced medially from its centre inElaphe dione.The termination of the external premaxillary process protrudes somewhat more laterally than the termination of the internal process in both species.However,the terminations of both processes can be in level only inElaphe dione.
Parietal(Figure 3) It is a large bone covering the brain dorsally and laterally.The lateral descending parts are strongly convex.The dorsal surface has a shallow sagittal groove,which divides it into two slightly convex halves.The dorsal surface is limited laterally and posteriorly by parietal crests,which approach each other in the posterior direction.The development of the ridges ranges from very sharp to blurred or to almost invisible.Bachmayer and Szyndlar (1987) noted that convergence of the parietal crests before reaching the posterior border of the bone is characteristic ofElaphe.However,the result of this convergence may be different.In some individuals,the parietal crests reach the posterior edge of the bone and remain separated (Figure 3A,D).In others,they merge and form a single crest (Figure 3B,C,E).There may be present a posterior process that bears the posterior terminations of the parietal crests (Figure 3C,E).The front edge of the bone is concave in dorsal view and is formed by two large lateral lobes separated medially by a smaller convex third lobe.Small foramina in various numbers are located behind the lateral parts of the lateral lobes.Paired parietal openings,well or poorly visible,sometimes doubled,are located approximately in the middle of the sagittal length (including the posterior process).
The sagittal groove inElaphe schrenckiibifurcates not far from the anterior margin of the bone.This element inElaphe dioneusually bifurcates in the anterior quarter and rarely near the anterior margin of the bone.
The parietal crests inElaphe schrenckiiconverge at a small distance from the posterior border of the bone and form a narrow crest on the relatively long posterior process (Figure 3E).Such crests inElaphe dioneapproach each other at the most at the posterior margin of the bone (Figure 3A,D) or at a small distance from it;in the latter case,they can diverge to the sides (Figure 3B) or form a crest on the short posterior process (Figure 3C).
Supraoccipital(Figure 4) The bone issub-pentagonal in shape,it is wider than long.The dorsal surface shows three crests:the sagittal crest running along the sagittal plane; two occipital crests located symmetrically at an angle to the sagittal crest.The occipital crests are inclined posteromedially and divide each half of the bone into supraoccipital and posterior areas.InElaphe dione,these three crests are moderately high or high.They converge before the anterior margin of the bone or reach it separately.The sagittal crest usually reaches the posterior margin of the bone;its height in this case is constant over the entire length.Less often,it does not reach the posterior margin,and its height gradually decreases posteriorly.The right and left branches of the occipital crest are directed posterolaterally and usually reach the margin of the bone;their height is almost constant over their entire length or decreases towards their ends.The supraoccipital orifices may be present in the supraoccipital area.
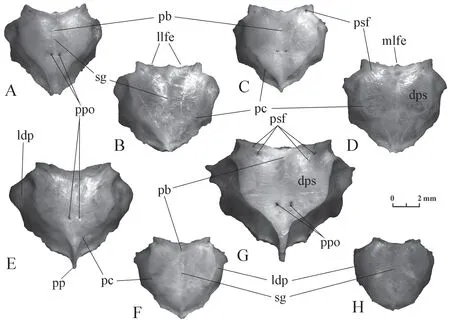
Figure 3 Parietal in dorsal view.A-D:morphological variants of Elaphe dione (A:specimen No.3;B:specimen No.4;C:specimen No.7; D:specimen No.8);E-H:morphological variants of Elaphe schrenckii (E:specimen No.2;F:specimen No.4;G:specimen No.7;H:specimen No.6).dps:dorsal parietal surface;ldp:lateral descending part;llfe:lateral lobe of front edge;mlfe:medial lobe of front edge;pb:point of sagittal groove bifurcation;pc:parietal crest;pp:posterior process;ppo:paired parietal openings;psf:parietal small foramina;sg:sagittal groove.

Figure 4 Supraoccipital in dorsal view.A-E:Elaphe dione (A:specimen No.3;B:specimen No.4;C:specimen No.7;D:specimen No.9;E:specimen No.10);F-I:Elaphe schrenckii (F:specimen No.1;G:specimen No.2;H:specimen No.3;I:specimen No.4.) asc:anterior sagittal crest;oc:occipital crest;pa:posterior area;psc:posterior sagittal crest;sc:sagittal crest;soa:supraoccipital area;soo:supraoccipital orifices.
The sagittal and occipital crests inElaphe schrenckiiare high and even very high.They usually converge in the anterior quarter of the bone length in the sagittal plane.As a result,the short anterior section of the sagittal crest (anterior sagittal crest) may be present before the confluence of the three ridges.The posterior section of the sagittal crest (posterior sagittal crest) usually does not reach the posterior edge of the bone.It has the same height for its short portion but posteriorly it reduces in size.The right and left branches of the occipital crest are directed almost laterally and usually do not reach the edge of the bone;they have the same height for their short portions but posteriorly they reduce in size.There are supraoccipital orifices in the supraoccipital area.
Maxilla(Figure 5) The bone is comparatively thin,either arcuate in dorsal view or has a straight lateral edge with the front and rear ends bent inward.Teeth are isodont,large,strongly bent backwards,and are of the same length throughout the continuous row.There are from 16 to 18 tooth positions,usually 17,inElaphe schrenckiiand from 15 to 18,usually 16 or 17,inElaphe dione.The number of tooth positions may differ in the right and left maxilla even in one individual.The ectopterygoid process is located in the back of the bone mesially.It is 2-3 times longer than wider and its length is equivalent to three or four tooth positions.One or two tooth positions are present behind the process (more often two inElaphe dioneand one inElaphe schrenckii).Prefrontal process is shorter (shorter than three teeth),but it protrudes more medially and narrows towards its distal termination.The middle of its base inElaphe dioneis usually opposite the 7th tooth positions,but can also be opposite the 7th and the 8th or the 7th and the 6th tooth positions.ForElaphe schrenckii,it is usually located opposite the 8th tooth positions and rarely opposite the 9th or opposite the 8th and 9th tooth positions.
Quadrate(Figure 6) This is an elongated bone with a joint (trochlea quadrati) for attaching to the compound bone of the lower jaw.Trochlea quadrati has two asymmetric lobes:a quadrate lateral knob,which is convex on the lateral side,and a quadrate medial knob flattened on the medial side.The quadrate bone widens proximally and its dorsal edge forms the dorsal crest.The quadrate crest extends more than half of the bone length along its lateral edge.The stapedial process is clearly visible on the medial side of the quadrate.
The proximal expansion of the bone inElaphe dionebegins almost from the trochlea quadrati.As a result,the general shape of the quadrate is almost triangular in most specimens.InElaphe schrenckii,the expansion begins closer to the middle of the bone and reaches a relatively lower value at the proximal edge.Therefore,the general shape of the quadrate bone is rather elongated than triangular.
The edge of the quadrate crest in mostElaphe dionespecimens is noticeably convex over most of the length,while inElaphe schrenckiiit is straight or even concave.
Palatine(Figure 7) The bone is long with isodont teeth of similar length strongly bent backwards.Their number is 9-10 inElaphe schrenckiiand 7-10 inElaphe dione.The bone has several processes.The largest vomerine process in the middle of the bone is trapezoidal in shape,its distal termination bends medially.Two palatopterygoid processes separated by a concavity are located at the posterior border of the bone.The palatopterygoid dorsal process is longer than the palatopterygoid ventral process.The maxillary (=lateral) process is located in the anterior half of the bone at the level between the second and the fifth (less often the sixth) tooth positions;the length of its base is equivalent to three or four tooth positions;the width of the process increases posteriorly;its posterior edge deviates slightly backwards (is rarely perpendicular to the axis of the bone).The maxillary process ends with a rounded tip.Two orifices for the maxillary nerve are located at the base of the maxillary process.The relative distance between them is smaller forElaphe schrenckiiand is equivalent to one tooth position.The posterior maxillary nerve orifice is located almost in the middle of the bone length.The posterior maxillary nerve orifice is noticeably shifted inElaphe dionefrom the middle of the bone forwards and the distance between the two foramina is 1.5-2 tooth positions.
Pterygoid(Figure 8) A long bone in ventral or dorsal view is slightly curved and has a series of isodont teeth.The anterior third of the bone is narrow,gradually expanding to the broad posterior part.The anterior end is divided into two processes,and the anterior lateral process is longer than the anterior medial one.An almost straight tooth row starts from the anterior end of the bone and extends approximately to the middle of its length.It contains 10-14 tooth positions inElaphe dioneand 11-13 tooth positions inElaphe schrenckii.Several posterior teeth are located on a wide part.The pterygoid flange is located on the lateral side at the transition of the narrow part of the bone to the wide part.A pointed ectopterygoid process is usually present inElaphe dione,while the ectopterygoid process inElaphe schrenckiiis wide,round,or completely absent.InElaphe schrenckii,the inner edge of the bone after the tooth row forms a downward medial kink.Elaphe dioneusually does not have such a kink.
Ectopterygoid(Figure 9)
The bone is Y-shaped,the relief of its ventral surface is indistinct.The long posterior branch is from straight to markedly curved,and has almost parallel or slightly diverging edges and a pointed end.The inner branch is pointed.The outer branch is wide;its distal edge is oriented at different angles to the posterior branch.The relative length of anterior branches of ectopterygoids differs in the studied species.The front angle of the outer branch inElaphe schrenckiiprotrudes slightly ahead of the inner branch end.The front angles of both branches are approximately at the same level or the inner (thin) branch protrudes slightly forward inElaphe dione.
Basiparasphenoid(Figure 10) The bone is long and can be divided into two parts:the posterior broad basisphenoid part and the anterior narrow parasphenoid process.
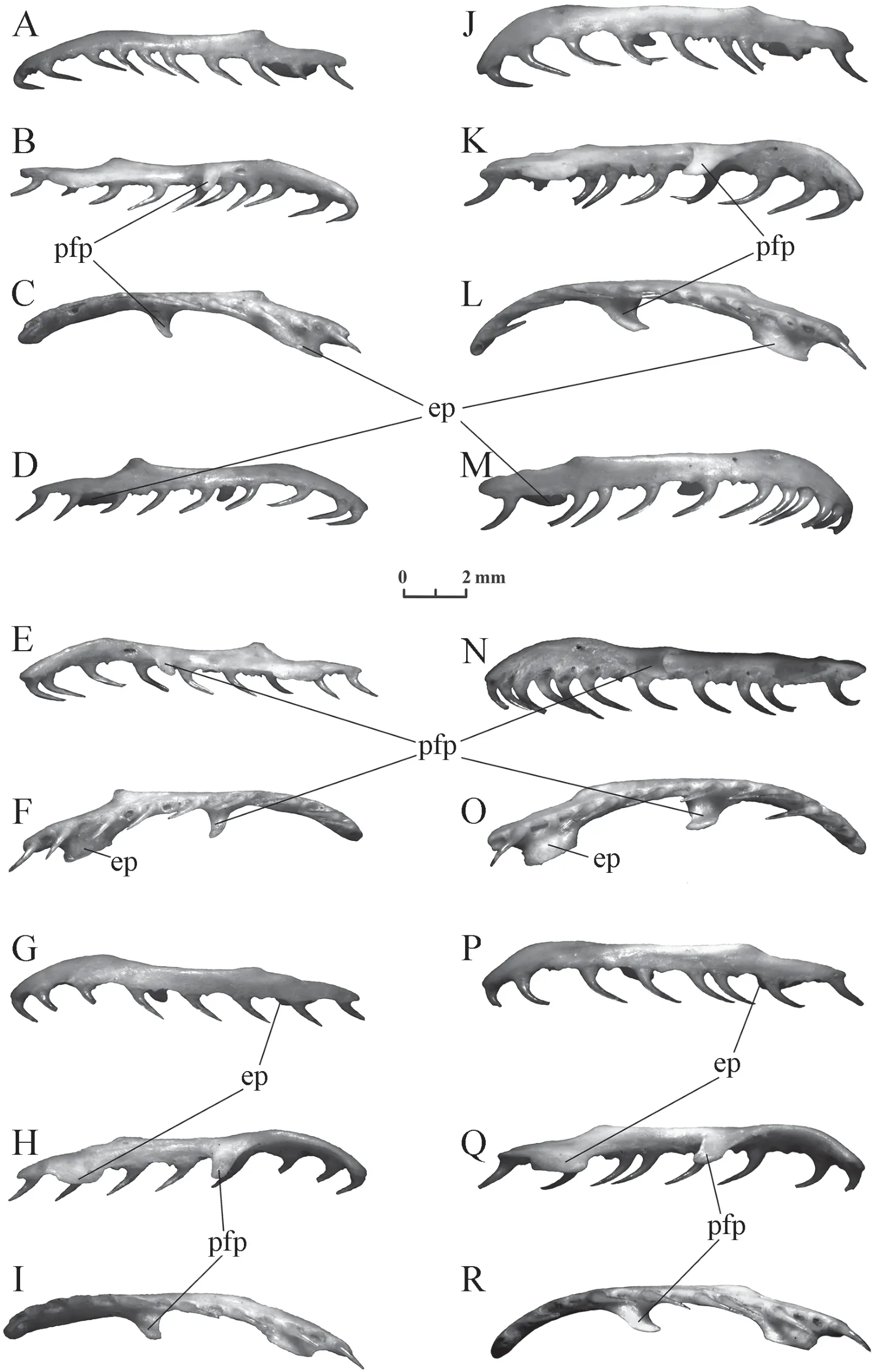
Figure 5 Maxilla of Elaphe dione (A-C:specimen No.4,sinister;D-F:specimen No.4,dexter;G-I:specimen No.8,sinister) and Elaphe schrenckii (J-L:specimen No.2,sinister;M-O:specimen No.2,dexter;P-R:specimen No.3,sinister).A,D,G,J,M,P:lateral view;B,E,H,K,N,Q:medial view;C,F,I,L,O,R:ventral view.ep:ectopterygoid process;pfp:prefrontal process.
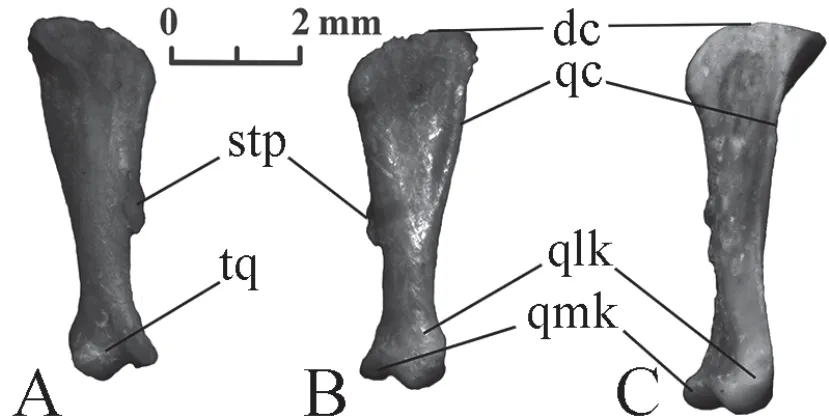
Figure 6 Right quadrate of Elaphe dione,specimen No.8 (A,B) and Elaphe schrenckii,specimen No.3 (C).A:anteromedial view;B,C:posterolateral view.dc:dorsal crest;qc:quadrate crest;qlk:quadrate lateral knob;qmk:quadrate medial knob;stp:stapedial process;tq:trochlea quadrati.
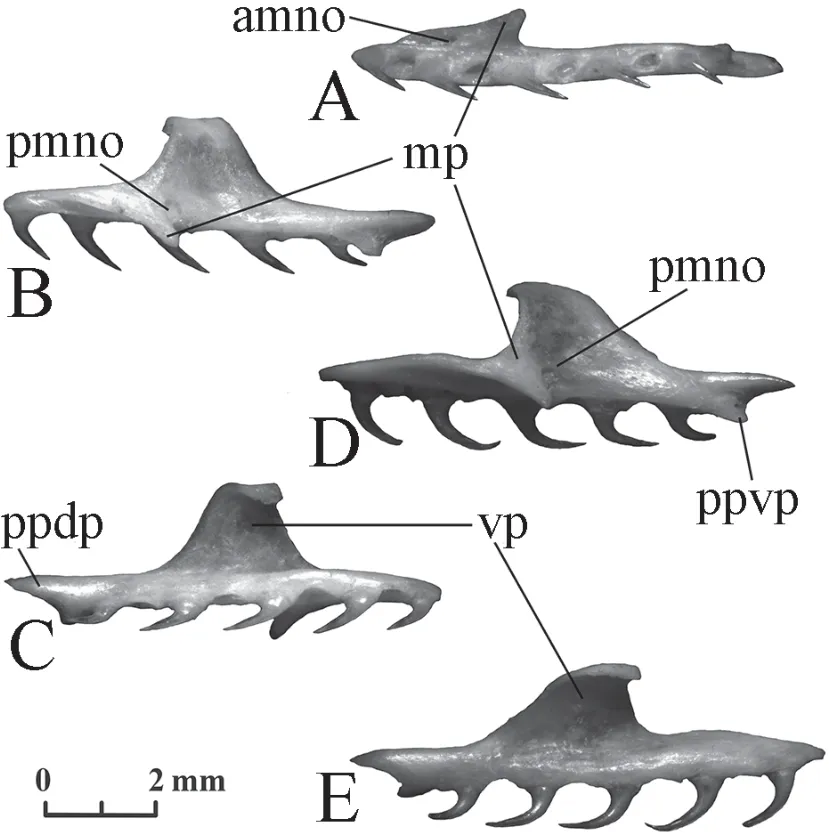
Figure 7 Left palatine of Elaphe dione,specimen No.8 (A-C) and Elaphe schrenckii,specimen No.3 (D,E).A:ventral view;B,D:lateral view;C,E:medial view.amno:anterior maxillary nerve orifice;mp:maxillary process (lateral process);pmno:posterior maxillary nerve orifice;ppdp:palatopterygoid dorsal process;ppvp:palatopterygoid ventral process;vp:vomerine process.
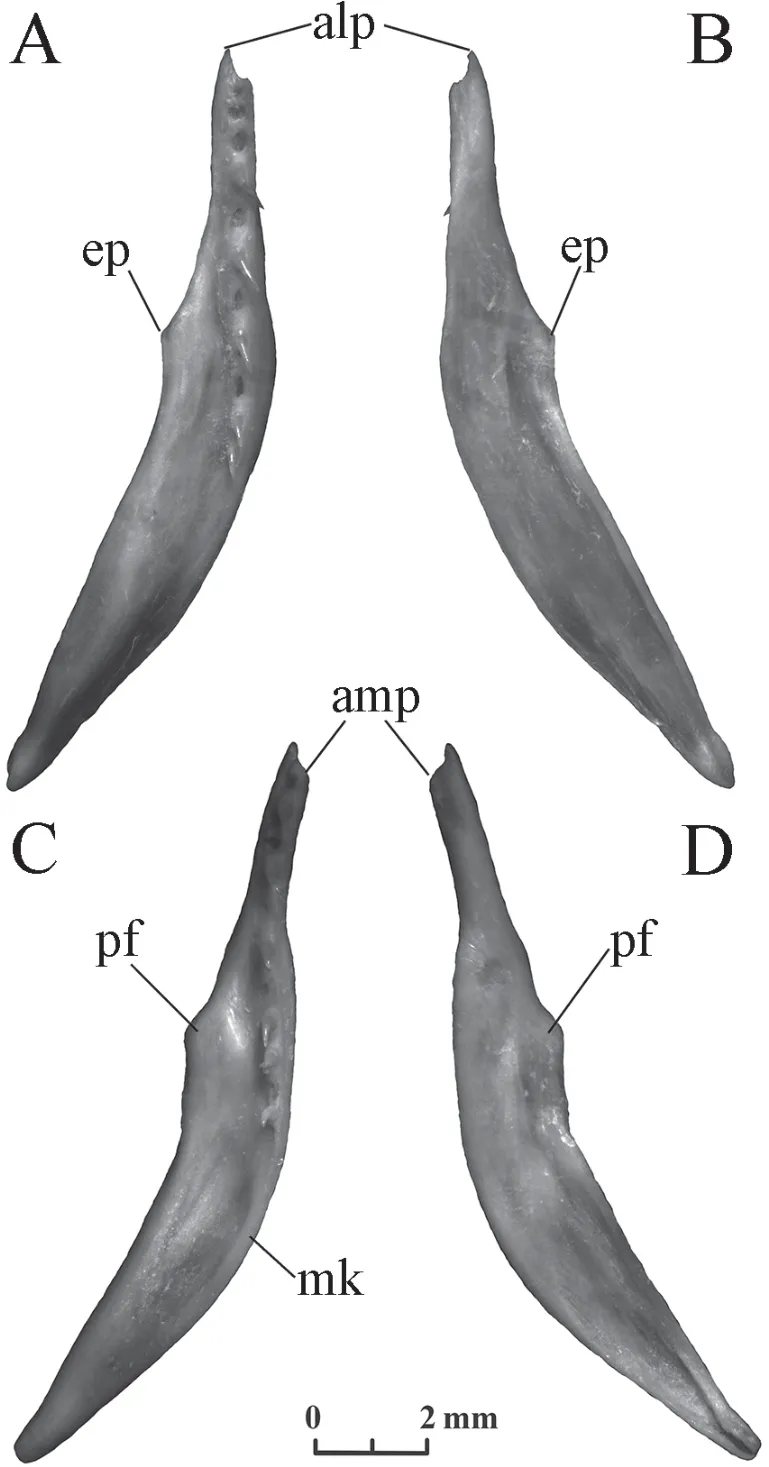
Figure 8 Right pterygoid of Elaphe dione,specimen No.8 (A,B) and Elaphe schrenckii,specimen No.3 (C,D).A,C:ventral view;B,D:dorsal view.alp:anterior lateral process;amp:anterior medial process;ep:ectopterygoid process;mk:medial kink;pf:pterygoid flange.

Figure 9 Right ectopterygoid of Elaphe dione (A:specimen No.3;B:specimen No.4) and Elaphe schrenckii,specimen No.3 (C),dorsal view.er:external ramus;ir:internal ramus.
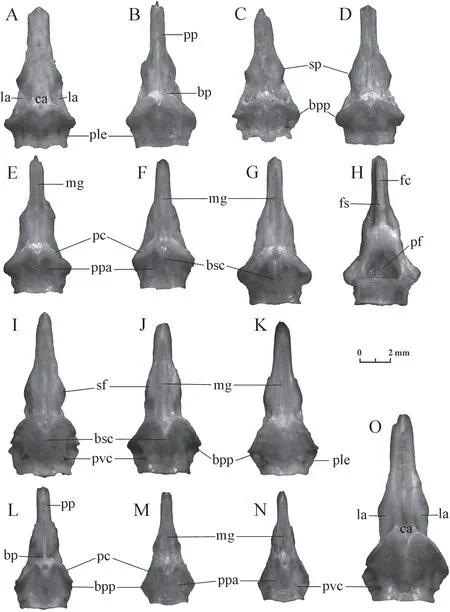
Figure 10 Basiparasphenoid of Elaphe dione (A:specimen No.1;B:specimen No.2;C:specimen No.3;D:specimen No.5;E:specimen No.6;F:specimen No.7;G,H:specimen No.8) and Elaphe schrenckii (I:specimen No.1;J:specimen No.2;K:specimen No.3;L:specimen No.4;M:specimen No.5;N:specimen No.6;O:specimen No.7).A-G,I-O:ventral view;H:dorsal view.bp:basisphenoid part;bpp:basipterygoid process;bsc:basisphenoid crest;ca:central area;fc:frontal crest;fs:frontal step;la:lateral area;mg:median groove;pc:pterygoid crest;pf:pituitary fossa (fossa hypophyseos);ple:posterolateral edge;pp:parasphenoid process;ppa:postpterygoid area;pvc:posterior orifice of the Vidian canal;sf:suborbital flange;sp:suborbital process.
The basisphenoid part of the bone is about 2/3 of its length.The anterior half of the basisphenoid part is much narrower than the posterior half.Its lateral edges at this point form more or less pronounced suborbital flanges.The central area is located on the ventral side in the centre of the basisphenoid part.Well-developed pterygoid crests diverge from the central area posterolaterally.They are inclined anterolaterally and close the anterior orifices of the Vidian canals opening in front of them.The posterior orifices of the Vidian canals are located more or less symmetrically in the postpterigoid area.Lateral areas are located in the hollows of the ventral surface on both sides from the central area.They are limited medially by pterygoid crests,as well as by low ledges or eroded ridges contouring the central area laterally.The lateral areas can be slightly wider or narrower than the central area.The basisphenoid crest extends backwards from the central area along the medial line.The degree of its development varies from clearly visible to almost indistinguishable.The length also varies,but it never reaches the posterior edge of the bone.The basipterygoid processes are located at the front ends of the right and left posterolateral edges of the basiparasphenoid.The posterior edge of the bone is usually tripartite,with more or less convex lobes.The deep pituitary fossa,usually of a round-triangular shape,is located on the dorsal side of the basisphenoid part of the bone.
The width of the parasphenoid process is constant or slightly varies.The ventral surface of this element has a median groove,whose length varies in different samples.The frontal crest is located on the dorsal side.It has parallel edges and a pointed or rounded end.The frontal step on the frontal crest is located approximately at the level of the base of the parasphenoid process and is developed differently (from a weak inflection to a distinct ledge).
The study revealed several differences in the morphology of this bone betweenElaphe dioneandElaphe schrenckii.The most reliable difference is the shape of the pterygoid ridges.Their height in the first species decreases laterally and their lateral ends merge with the lateral edges of the bone.The basipterygoid processes are rounded and rather broad.The height of pterygoid ridges in the second species does not change laterally;their lateral ends rise above the lateral edge of the bone.Basipterygoid processes are relatively acute.
Suborbital flanges in the Amur ratsnake are blunt or undeveloped,while in the Dione’s ratsnake they are usually sharp with a suborbital process,however,they can be undeveloped.
The length of the median groove inElaphe dionevaries widely.However,a short median groove extending only in the limits of the parasphenoid process was not found in the studied specimens ofElaphe schrenckii.
The lateral areas inElaphe schrenckiiare narrower than the central area,while inElaphe dionethey may be both wider or narrower.
The posterolateral margin of the bone is concave inElaphe dioneand almost straight inElaphe schrenckii.
InElaphe dione,there is a smooth narrowing of the basisphenoid part of the bone in its middle,while in most individuals ofElaphe schrenckii,with the exception of small specimens,the narrowing is rather sharp.
The ratio of the bone length to the distance between the basipterygoid processes inElaphe dione(1.8-2.09) is usually slightly smaller than inElaphe schrenckii(2.05-2.43).That is why the bone ofElaphe schrenckiilooks longer.
Basioccipital(Figure 11) In ventral view,the bone outline is almost pentagonal,it is slightly or noticeably wider than long.The basioccipital crest extends in the anterior half of the bone in the transverse direction and almost reaches the lateral edges.As a rule,it forms three bends called lobes:one medial and two lateral.Lateral lobes are usually better developed (they form a sharper relief on the bone surface) than the medial lobe (basioccipital process),especially in large specimens.The medial lobe merges with the medial crest,which posteriorly divides into two diverging branches and is separated by a groove from the occipitocondylar tubercle.Basioccipital tubercles are located on the lateral corners of the pentagon.The degree of development for all these structures is very unstable (from almost indistinguishable to clearly visible) and is not characteristic of the species.The anterior edge of the bone is straight or more often concave.

Figure 11 Basioccipital of Elaphe dione (A:specimen No.1;B:specimen No.4;C:specimen No.5;D:specimen No.10) and Elaphe schrenckii (E:specimen No.1;F:specimen No.4;G:specimen No.6;H:specimen No.7),ventral view.boc:basioccipital crest;bop:basioccipital process;bot:basioccipital tubercle;ll:lateral lobe;mc:medial crest;ot:occipitocondylar tubercle (condylus basioccipitalis).
The difference between the species lies in the lateral lobes of the basioccipital ridge.They are more strongly pulled back inElaphe schrenckii;their lateral edges form an angle of about 30° to the longitudinal axis of the bone,while inElaphe dionethis angle is closer to 60°.
It should be noted that the lateral lobes are not always detected.In the materials,one small specimen ofElaphe schrenckiiand one specimen ofElaphe dionedid not have pronounced lateral lobes.Apparently,in this case the identification of this species based on the bone is impossible.
Dentary(Figure 12) The bone is long,slightly curved,and carries a number of isodont teeth.Their height gradually decreases from front to back.The back of the bone forms three processes of different lengths:posterodorsal process (the longest process bears the posterior portion of the tooth row),posteroventral process (of an intermediate length),and posteromedial process (the shortest).Meckel's groove on the medial side of the bone is usually completely closed at the level of the 4-6th tooth positions.The mental foramen of elongated or more rarely rounded form,as well as a compound notch,is located on the lateral side of the bone.
Elaphe dionehas 17-20 tooth positions.The mental foramen lies at the level of 7-8th,8th,or 8-9th tooth positions.The compound notch usually approaches the 10-11th tooth positions.
Elaphe schrenckiihas 19-22 tooth positions.The mental foramen lies at the level of 9-10th,or more rarely 8-9th tooth positions.The compound notch usually approaches the 10-12th tooth positions.
Compound bone(Figure 13) The bone is long,slightly curved,with a posteriorly situated mandibular joint for attaching to the quadrate bone.This joint is composed of two knobs (the rostral and caudal knob) separated by a sigmoid fossa.Posteriorly from the joint,there is a retroarticular process,which curves in the medioventral direction.Anteriorly from the joint,there is a long mandibular pit bounded by the medial and lateral flanges and located in the posterior half of the bone.The medial flange merging into the coronoid process is curved in the dorsal direction and is 1.5-2 times higher than the lateral (=labial) flange,which has an almost straight dorsal edge.A subcoronoid groove may be present on the medial flange below the coronoid process.
The ventral supraangular crest in the compound bone ofColuber jugularis(Szyndlar,1984,Figure 3,IV),which is now calledDolichophis caspius,is marked under the joint for attachment to the quadratum at the posterior end.Raccaet al.(2020) observe this well-developed crest inHierophis viridiflavusalong the entire bone.In the studied specimens ofElaphe dioneandElaphe schrenckii,a pronounced ventral crest with a pointed edge is only present in the anterior part of the bone where there is contact with the angulare.A lateral compression is only observed under the retroarticular process and the joint,but is absent in front of it on the entire posterior half of the bone.The dorsal supraangular crest,which is in contact with the angulare from the other edge,is developed on the dorsal side on the anterior half of the compound bone.It extends to the most distal point of the bone.The third,medial supraangular,crest is located on the medial side of the anterior half of the compound bone.The supraangular foramen is located on the line of the upper lateral flange edge,a small distance from the level of the anterior end of the coronoid process.
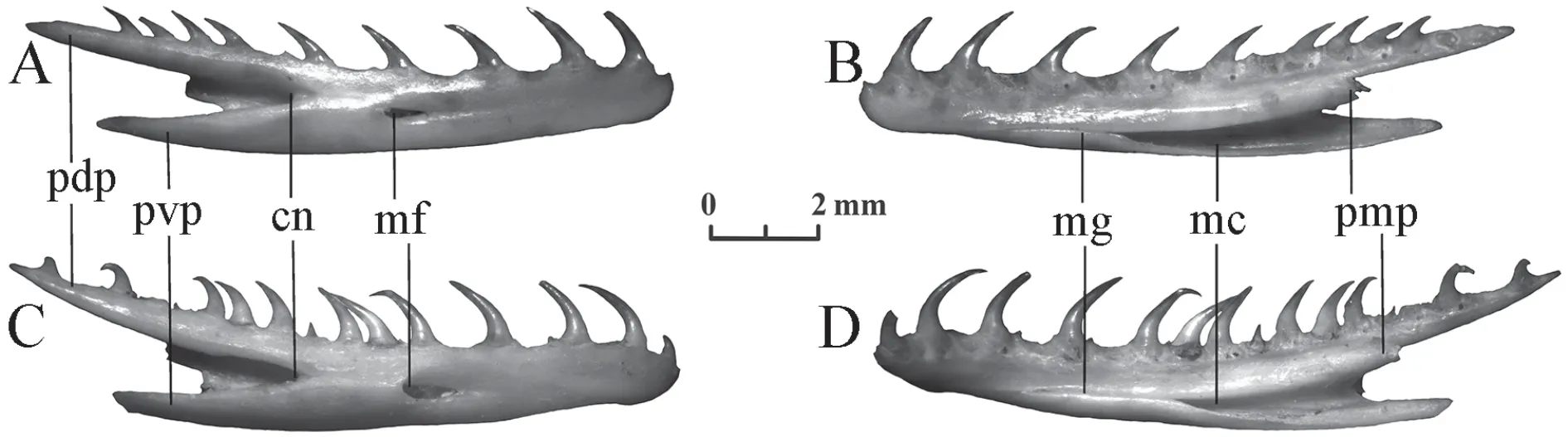
Figure 12 Dentary of Elaphe dione,specimen No.8 (A,B) and Elaphe schrenckii,specimen No.3 (C,D).A,C:ventral view;B,D:dorsal view.cn:compound notch;mf:mental foramen (dental foramen);mc:Meckel’s canal;mg:Meckel’s groove;pdp:posterodorsal process;pmp:posteromedial process;pvp:posteroventral process.
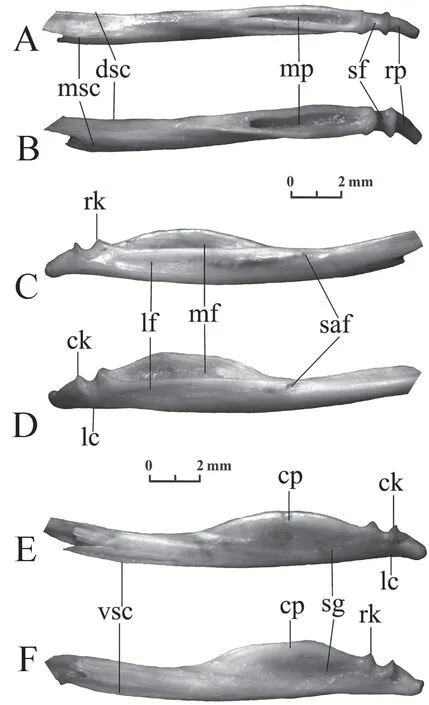
Figure 13 Compound bone of Elaphe dione,specimen No.8 (A,C) and Elaphe schrenckii,specimen No.3 (B,E).A,E:ventral view;B,C:dorsal view.ck:caudal knob of the joint;cp:coronoid process;dsc:dorsal supraangular crest;lc:lateral compression;lf:lateral flange (labial flange or process);mf:medial flange (lingual flange or process);mp:mandibular pit;msc:medial supraangular crest;rk:rostral knob of the joint;rp:retroarticular process;saf:supraangular foramen; sg:subcoronoid groove;sf:sigmoid fossa;vsc:ventral supraangular crest.
The coronoid process inElaphe schrenckiiis medially inclined;the subcoronoid groove is well developed and is usually clearly visible from the anterior to the posterior terminations of the coronoid process,especially in the posterior half of the bone.
The coronoid process inElaphe dioneis not inclined or is slightly inclined medially;the subcoronoid groove is poorly developed and is usually visible under the posterior part of the coronoid process.
3.2.Description of the vertebraeSzyndlar (1984) subdivided the ophidian vertebral column of snakes into four basic regions:cervical,trunk,cloacal,and caudal vertebrae.The trunk vertebrae are most numerous in the skeletons of snakes,they do not change much along the vertebral column and are therefore most convenient for species identification.The cervical vertebrae change in morphology very quickly from front to back and are very diverse within one species.On the contrary,cloacal and caudal vertebrae do not have any characteristic features (apart from erycine snakes) and representatives of both different species and larger taxa can have cloacal and caudal vertebrae of a similar shape.The identification of species from these vertebrae is possible if they retain some expressive features inherent in the trunk vertebrae.
Trunk vertebraeAs representatives of“Colubrinae”,Elaphehave trunk vertebrae without hypapophyses (Szyndlar,1991).In addition,their trunk vertebrae do not have parapophyseal processes.The progressive changes in the structure of the vertebrae are observed from front to back:the relief of the ventral surface is more distinct,the subcentral grooves deepen,the subcentral ridges increase;the width of the hemal keel decreases and its height increases,and the height of neural spines and laminae may also decrease.Several presacral vertebrae show a rapid antero-posterior shortening.In addition,random changes in morphological elements add variants in their variability.In general,there is a significant overlap of features in the vertebrae ofElaphe dioneandElaphe schrenckii,mainly due to the very strong variability ofElaphe dione.Since a description of the trunk vertebrae has already been published (Ratnikov,2004),I will focus on the main differences and the detected variability.
Elaphe dione(Figure 14).Neural spines are low or moderately high (about 2-4 times longer than height).The posterior end of the neural spine always protrudes posteriorly above its base.In addition,the anterior end can rarely overhang and protrude.Occasionally,a thickening is observed along the upper edge of the neural spine (Figure 14F).Laminae are more or less convex in posterior view,from fairly high to low.The anterior edge of the zygosphene in anterior view is straight or slightly convex.The width of the zygosphene ranges from relatively narrow to wide.In dorsal view,the zygosphene lobes are clearly expressed,equally developed,or the median lobe is more (sometimes considerably) or less developed;vertebrae with an undeveloped median lobe are occasionally observed;a medial notch in the median lobe is observed even less often (Figure 14 J2).The width of the zygosphenal lobes varies:most often the median lobe is wider than the lateral lobes;however,the width of the lateral lobes can also be almost equal to the width of the median lobe.Prezygapophyseal processes are usually narrow,of moderate length,directed antero-laterally (more laterally) with pointed or rounded distal tips.However,the processes can also be short,long,wide or directed to the side.Interzygapophyseal ridges are sometimes developed so strongly that the vertebra from above appears wide (Figure 14 F2).The centrum is relatively long and narrow.The lateral edges of the hemal keel in the anterior trunk vertebrae may merge with the vertebral body.The posterior end of the hemal keel can be chopped off,rounded or pointed,but it does not hang over the surface of the centrum and gradually decreases in height.Subcentral ridges appear on the posterior trunk vertebrae.They are separated from the hemal keel by subcentral grooves.

Figure 14 Trunk vertebrae of Elaphe dione.A-D:specimen No.6,A:anterior trunk vertebra,B:middle trunk vertebra,C:posterior trunk vertebra,D:predcloacal trunk vertebra;E:specimen No.1,posterior trunk vertebra;F:specimen No.12,anterior trunk vertebra;G-I:specimen No.8,G:anterior trunk vertebra,H:middle trunk vertebra,I:posterior trunk vertebra;J:specimen No.9,predcloacal trunk vertebra;K:specimen No.7,posterior trunk vertebra;L:specimen No.10,anterior trunk vertebra;M:specimen No.2,posterior trunk vertebra.1:anterior view;2:dorsal view;3:lateral view;4:posterior view;5:ventral view.c:centrum;con:condylus;cot:cotylus; hk:hemal keel;ir:interzygapophyseal ridge;l:lamina;llz:lateral lobe of zygosphene;mlz:median lobe of zygosphene;mn:medial notch;nc:neural canal;ns:neural spine;prp:presigapophyseal processes;sg:subcentral grooves;sr:subcentral ridges;z:zygosphene.
Elaphe schrenckii(Figure 15).Neural spines are high or moderately high (about 1.5-3 times longer than height).Prominences of the front and rear ends above the base of the neural spine are small.A thickening along the upper edge of the neural spine is rarely observed.Laminae are more often high,slightly convex,almost straight or even concave in posterior view,similar to a gable roof;they smoothly turn into the neural spine (Chenet al.,2019;Figure 4).Less commonly,especially in small individuals,laminae can be compressed dorsoventrally,more or less convex,but at the same time quite high,very similar to the vertebrae ofElaphe dione.The front edge of the zygosphene in anterior view is more or less convex.The median lobe of the zygosphene is usually less developed than the lateral lobes,it is often completely absent,especially in large individuals.In this case,the front edge of the zygosphene in dorsal view is more often concave.However,vertebrae with equally developed lobes and even with a more developed median lobe are sometimes found.Prezygapophyseal processes are short,pointed,directed to the sides and slightly forwards,but can reach a moderate length and become blunt.The centrum looks short and wide.Its ventral surface is flattened and laterally bounded by a clear bend.The lateral margins of the hemal keel in the anterior trunk vertebrae are usually clearly outlined.Hemal keels in most vertebrae are pointed at the end;their tips in the anterior and middle trunk vertebrae in the form of a step rise above the surface of the centrum,while the keels of posterior trunk vertebrae themselves become high.Subcentral ridges appear on the posterior trunk vertebrae.They merge with the bend limiting the ventral surface of the centrum.
Cervical vertebraeIn representatives ofElaphe,short parapophyseal processes are only found in the cervical vertebrae.The structure of the atlas does not allow species diagnostics.Moreover,this vertebra consisting of several bone elements connected by ligaments,usually falls apart during burial.The epistropheus has a structural plan that is different from all other vertebrae and can be used for species identification.The vertebrae,starting from the third,have a similar structural plan,which undergoes gradual changes in the caudal direction:vertebrae lengthen and increase in size;the neural spine lengthens to a normal length for the species;the hypapophysis changes its shape and is replaced by a hemal keel at the end of the cervical part of the vertebral column.In addition,there are changes in the zygosphene which continue in the subsequent vertebrae.
Elaphe dione(Figure 16).The median lobe of the zygosphene in the front vertebrae is less developed than the lateral ones and may even be absent (in this case,the front edge of the zygosphene in dorsal view can be concave).However,then it increases in size towards the tail.Prezygapophyseal processes can be from short to rather long.Laminae can be high,very similar to the laminae ofElaphe schrenckii.The hypapophyses in the anterior cervical vertebrae are bent and directed backwards,almost like inNatrix;they usually have a rounded end (however,in our comparative collection there is a specimen with a pointed hypapophysis).Gradually,the hypapophyses straighten (but at the same time they are relatively shorter than those ofElaphe schrenckii),and then bend forwards;their distal termination becomes narrower and sharper.The posterior edge of the straight hypapophyses approaches the condylus in a smooth arc.The end of the bent-forward hypapophysis in some vertebrae of adults resembles a long narrow excrescence directed forwards.The hypapophyses in the posterior cervical vertebrae gradually decrease until they turn into a hemal keel.
The neural spine of the epistropheus is low;usually it does not overhang or weakly overhangs at the front and strongly overhangs at the back.The front end of its base can be slightly displaced away from the front end of the neural arc (up to 1/6 of its length).The hypapophysis is directed backwards and downwards at an angle of 30°-45°.
Elaphe schrenckii(Figure 17).The median lobe of the zygosphene is less developed than the lateral lobes and may even be absent (in this case the anterior edge of the zygosphene in dorsal view may be concave).Its size gradually increases from the anterior cervical vertebrae to the caudal vertebrae and it very rarely exceeds the size of the lateral lobes.Prezygapophyseal processes can be from short to rather long.Laminae can be convex,very similar to the laminae ofElaphe dione.The hypapophyses in the anterior cervical vertebrae are straight,relatively longer than in theElaphe dione;they have a narrowed rounded end and are directed downwards and slightly backwards.The posterior edge of such hypapophyses is usually quite close to the condylus.Gradually,the hypapophyses deviate forwards;their rounded end is displaced from the posterior edge of the hypapophysis to the anterior edge and becomes acute.The end of the bent-forward hypapophysis in some vertebrae of large individuals resembles a long narrow excrescence directed forwards as inElaphe dione.The hypapophyses in the posterior cervical vertebrae gradually decrease until they turn into a hemal keel.
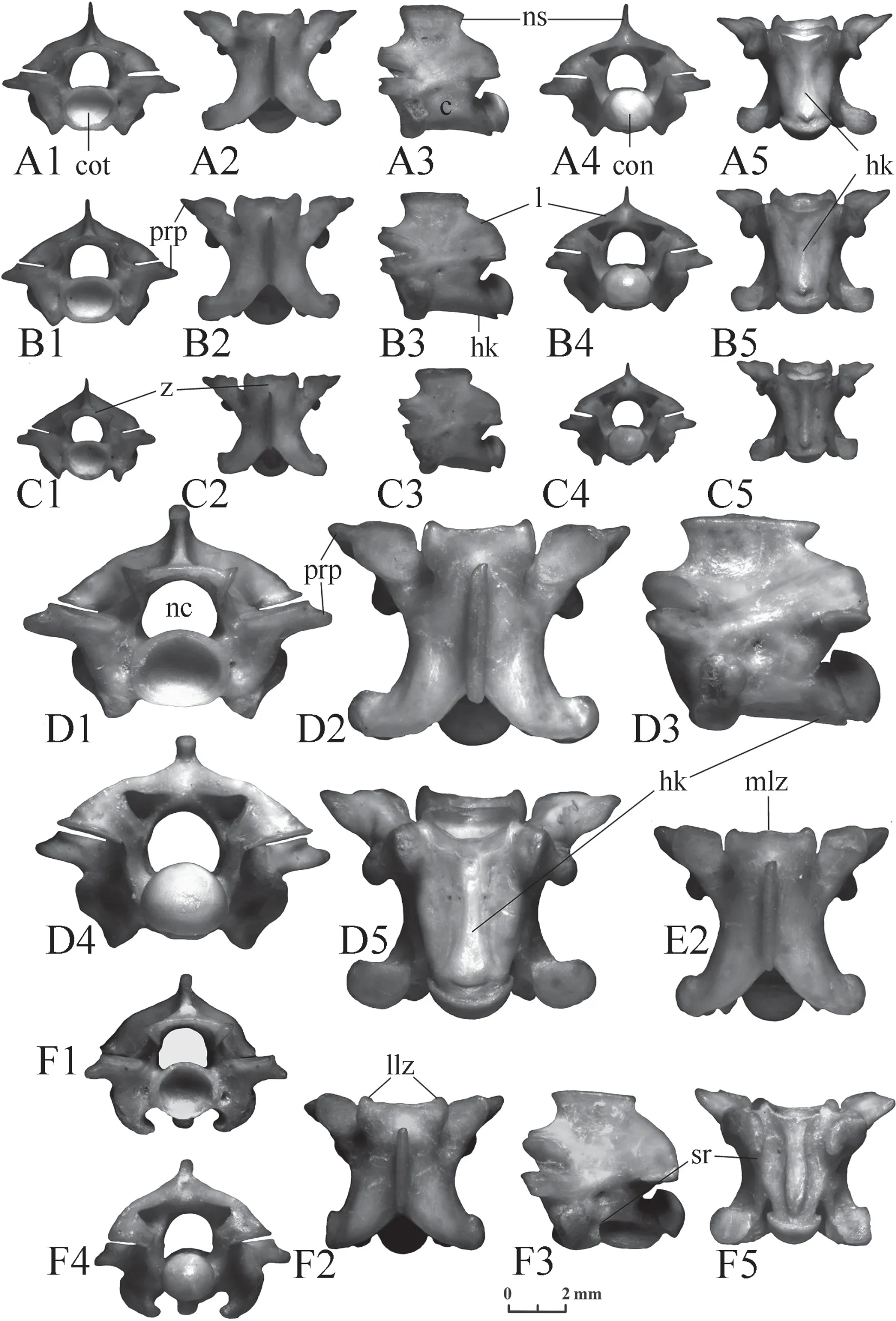
Figure 15 Trunk vertebrae of Elaphe schrenckii.A-C:specimen No.3,A:anterior trunk vertebra,B:middle trunk vertebra,C:posterior trunk vertebra;D,F:specimen No.8,D:middle trunk vertebra,F:predcloacal trunk vertebra;E:specimen No.7,middle trunk vertebra.1:anterior view;2:dorsal view;3:lateral view;4:posterior view;5:ventral view.c:centrum;con:condylus;cot:cotylus;hk:hemal keel;l:lamina;llz:lateral lobe of zygosphene;mlz:median lobe of zygosphene;nc:neural canal;ns:neural spine;prp:presigapophyseal processes;sr:subcentral ridges;z:zygosphene.

Figure 16 Cervical vertebrae of Elaphe dione,specimen No.8.A-G:changes in morphology from the third cervical vertebra (A) to the last (G);H:epistropheum.1:anterior view;2:dorsal view;3:lateral view;4:posterior view;5:ventral view.c:centrum;con:condylus;cot:cotylus;h:hypapophyses;l:lamina;llz:lateral lobe of zygosphene;mlz:median lobe of zygosphene;nc:neural canal;ns:neural spine;pap:parapophyseal processes;prp:presigapophyseal processes;z:zygosphene.

Figure 17 Cervical vertebrae of Elaphe schrenckii.A-E:changes in morphology from the third cervical vertebra (A) to the posterior (E),specimen No.3;F,G:middle cervical vertebrae,specimen No.7;H,I:middle cervical vertebrae,specimen No.8;J:epistropheum.1:anterior view;2:dorsal view;3:lateral view;4:posterior view;5:ventral view.c:centrum;con:condylus;cot:cotylus;h:hypapophyses;l:lamina;llz:lateral lobe of zygosphene;mlz:median lobe of zygosphene;nc:neural canal;ns:neural spine;pap:parapophyseal processes; prp:presigapophyseal processes;z:zygosphene.
The neural spine of the epistropheus is high;its ends strongly with an overhang at the front and at the back.The front end of its base is strongly retracted from the front end of the neural arc (from 1/3 to 1/2 of its length).The hypapophysis is directed more downwards and slightly backwards at an angle of about 60°.
Cloacal and caudal vertebrae(Figures 18,19) As has been mentioned above,these vertebrae are similar to each other.The cloacal vertebrae are short,while the caudal vertebrae gradually lengthen from the anterior to the posterior.A large variation interval for the size and proportions does not allow us to suggest the location of a separate vertebra:it can either come from the front of the tail of a small individual or from the back of the tail of a large one.The paradiapophyses,hypapophysis,and hemal keel are replaced by lymphapophyses,hemapophyses,and pleurapophyses.These elements do not have any clear species differences,although they vary greatly in size and shape even in a single individual.Thus,there are fewer morphological features in comparison with precloacal vertebrae.This makes them very difficult to identify.It is almost impossible to distinguish between the small vertebrae ofElaphe dione(Figure 18) andElaphe schrenckii(Figure 19).However,it is possible to determine the vertebrae of large individuals ofElaphe schrenckii(Figure 19) with specific features of the species.Such features include a high neural spine,a convex zygosphene in anterior view with an undeveloped median lobe,and high and weakly arched laminae.The large vertebrae are also massive,with a thick neural arch.
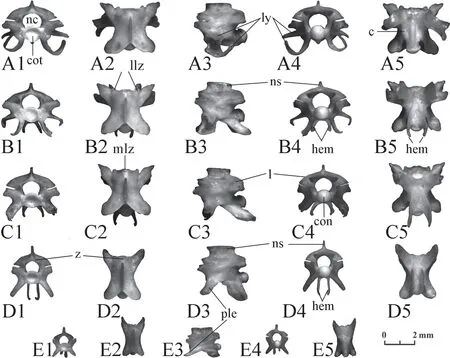
Figure 18 Cloacal (A,B) and caudal (C-E) vertebrae of Elaphe dione,specimen No.8.1:anterior view;2:dorsal view;3:lateral view;4:posterior view;5:ventral view.c:centrum;con:condylus;cot:cotylus;hem:hemapophyses;l:lamina;llz:lateral lobe of zygosphene;ly:lymfapophisis;mlz:median lobe of zygosphene;nc:neural canal;ns:neural spine;ple:pleurapophyses;z:zygosphene.

Figure 19 Cloacal (A,F) and caudal (B-E,G-H) vertebrae of Elaphe schrenckii.A-E:changes in morphology,specimen No.8; F-H:changes in morphology,specimen No.3.1:anterior view;2:dorsal view;3:lateral view;4:posterior view;5:ventral view.c:centrum;con:condylus;cot:cotylus;hem:hemapophyses;l:lamina;llz:lateral lobe of zygosphene;ly:lymfapophisis;mlz:median lobe of zygosphene;ns:neural spine;ple:pleurapophyses;z:zygosphene.
4.Discussion and Conclusions
The variability in the bone morphology depends on several parameters.First,these are species differences (as well as generic,family,etc.),which determine the systematic position of the animal.The detection of these differences constitutes the basis for determining fossil bones.Second,it is individual variability.It makes identification difficult if there is an overlap of variability intervals of species characteristics.Third,(this only applies to vertebrae),these are progressive changes in the morphology of the vertebrae along the vertebral column from the first vertebra to the last.They are observed in all vertebrates.Identifying the position of the vertebra in the vertebral column helps to identify it correctly.Fourth,it is age-related variability.As a rule,the bone morphology of large individuals reflects the typical features of the species better and includes more small structural details.
Despite the strong similarities between the osteology ofElaphe dioneandElaphe schrenckii,a careful examination of the skull bones allows us to identify the species for most of them.The revealed differences are shown in Table 2.

Table 2 Distinctive features in the skull bones of Elaphe dione and Elaphe schrenckii.
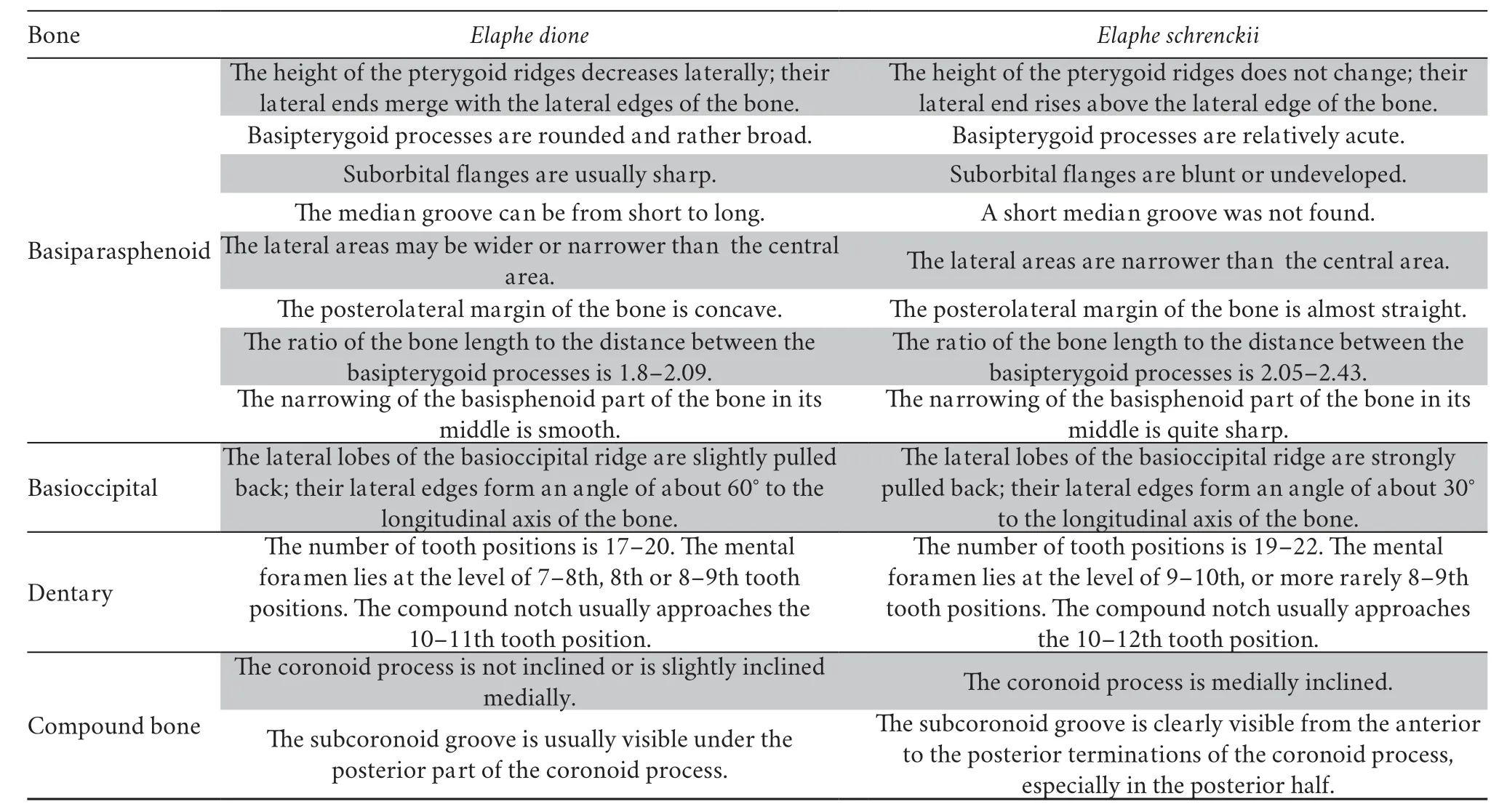
Continued Table 2
As we can see,both species of snakes have a certain variability.This variability is especially pronounced inElaphe dione.As a result,the features unique to only one species are quite rare.More often,there is an overlap in the variability intervals of features.In other words,the species can be identified unambiguously if the value of the feature does not fall into the overlapping zone.Otherwise,if the bones do not have other differences,it is only possible to identify a higher taxon.
The structure of snake vertebrae is very complex.They have a large number of changing elements.Choosing a single criterion to distinguish between the fossil vertebrae of twospecies will not always give the correct result.They need to be compared by a set of criteria.
The typical combination of features distinguishing the vertebrae ofElaphe dionefromElaphe schrenckiiare as follows.(1) The centrum is relatively long and narrow.(2) The neural spine is low and can strongly overhang both posteriorly and anteriorly.(3) The laminae in posterior view are convex and comparatively low.(4) The front edge of the zygosphene in anterior view is straight.(5) The zygosphene lobes in dorsal view are clearly expressed,equally developed,or the median lobe is developed more strongly (sometimes very strongly).(6) Prezygapophyseal processes are of moderate length,directed antero-laterally.(7) The posterior end of the hemal keel does not hang over the surface of the centrum and gradually decreases in height.
The typical combination of features distinguishing the vertebrae ofElaphe schrenckiifromElaphe dioneare as follows.(1) The centrum is relatively short and wide.(2) The neural spine is high;the anterior and posterior ends overhang slightly over the base of the neurapophysis.(3) The laminae are high,almost straight or even concave in posterior view,they smoothly merge into the neural spine.(4) The front edge of the zygosphene in anterior view is convex.(5) The median lobe of the zygosphene is not developed.(6) Prezygapophyseal processes are comparatively short,directed to the sides and slightly forwards.(7) The posterior end of the hemal keel in the form of a step rises above the surface of the centrum.
Unfortunately,typical morphology is not always observed.Many vertebrae have features found in both species.In these cases,species identification is carried out by the prevailing number of traits or the identification is limited to a higher taxon.
AcknowledgementsThe author thanks M.Tiunov,I.Maslova and V.Orlova for providing materials for a comparative osteological collection and I.Charychanskaya and S.Cox for the translation into English.The author is also grateful to H.Blain and two anonymous reviewers for the constructive comments that improved the quality of the manuscript.
杂志排行

Asian Herpetological Research的其它文章
- Offspring Sex Is Not Determined by Gestation Temperature in a Viviparous Lizard (Eremias multiocellata) from the Desert Steppe of Inner Mongolia
- Sex But Not Altitude,Modulates Phenotypic Covariations Between Growth and Physiological Traits in Adult Asiatic Toads
- Circadian Rhythm and Intersexual Differences in Auditory Frequency Sensitivity in Emei Music Frogs
- High-elevation Adaptation of Motion Visual Display Modifications in the Toad-Headed Agamid Lizards (Phrynocephalus)
- An Annotated List of Lizards (Sauria:Squamata) Recorded from the People’s Republic of China
- Appendix 1