3个桃品种对淹水胁迫的生理响应及耐涝性评价
2022-03-22王玉玲周晨浩肖金平古咸彬张慧琴李南羿张岚岚
王玉玲,周晨浩,肖金平,古咸彬,张慧琴,李南羿 ,张岚岚
(1. 浙江农林大学园艺科学学院/浙江省山区农业高效绿色生产协同创新中心,浙江 杭州 311300;2. 浙江省农业科学院园艺研究所,浙江 杭州 310021)
0 引言
【研究意义】桃(Prunus persicaL.)是核果类果树中最重要的树种,也是近年来发展最快的果树之一,桃产业在农民增收和乡村振兴中发挥了重要作用。桃由于根系较浅、呼吸作用旺盛,生产中易受到涝害影响。全球气候变暖导致异常天气增多,其中洪涝对农业生产构成极大威胁[1]。浙江等地的南方桃产区受梅雨和台风影响极易发生桃园内涝,不仅影响当年产量而且加速了树势的衰败。桃涝害防御除综合技术的研发集成外,同时应加强抗性砧木和品种的评价筛选,以及对不同类型桃品种的涝害生理特性的研究,这对于深入了解桃抗涝机制及有针对性地提出抗涝技术措施具有重要的理论价值和实践意义。【前人研究进展】广义的涝害包括渍害和涝害,土壤表层存在明显积水为涝害,土壤含水量过饱和但未见积水为渍害。渍害和涝害都会对植物造成淹水胁迫,从而降低土壤中O2溶解度,形成缺氧或无氧的根系环境[2-4]。植物对淹水胁迫生理响应的研究主要集中在植株光合特性及抗氧化保护酶等方面,在大田和蔬菜作物上研究较多如玉米[5]、黄瓜[6],而对果树特别是栽培品种方面研究相对较少。果树对淹水胁迫的形态响应与其他作物表现基本一致,苹果砧木幼苗在涝害低氧胁迫下根长、株高、生物量较对照明显降低[7]。淹水导致桃光合作用受阻,但不同品种光合参数指标变化幅度并不一致,耐涝性较好的品种往往能够表现出更好的光能利用效率[8]。淹水还会加速活性氧(ROS)的产生,影响细胞稳态。短期淹水胁迫时植物依靠抗氧化酶系统维持ROS平衡,减缓氧化损伤程度[9-10],如淹水处理下桃幼树叶片SOD、POD和过氧化氢酶(CAT)酶活性呈现前期升高响应胁迫,但到后期由于植株受损严重出现降低趋势[11]。耐涝植物如欧白英、杨树可在乙烯参与下形成通气组织和不定根以适应淹水胁迫[12-13]。古咸彬等[14]也发现猕猴桃耐涝砧木浙猕砧1号在淹水22 d后茎基部皮孔增大,并伴有根状组织发生,且随着淹水时间延长气生根数量增多。【本研究切入点】模拟涝害条件下不同成熟期的主栽桃品种植株形态及生理特性的响应变化,以及桃耐涝性综合评价指标有待深入探讨。【拟解决的关键问题】通过模拟涝害环境,揭示3个不同成熟期桃品种在淹水胁迫下植株形态、渗透调节、光合特性和抗氧化酶活性等方面的差异,并评价其耐涝性,以期为桃耐涝机制研究及耐涝品种的评价筛选提供一定的理论依据和参考。
1 材料与方法
1.1 材料及处理方法
本试验在浙江农林大学平山试验基地进行。供试品种为早熟硬质桃品种春红、中熟软溶质桃品种新川中岛和晚熟硬溶质桃品种锦绣,均为1 年生嫁接苗,砧木为实生播种的普通毛桃苗。
2019年3月下旬选取大小长势基本一致的3个品种嫁接苗,定植于直径和高度分别为35 cm和30 cm的无纺布美植袋中。袋土为m(园土):m(基质):m(有机肥)=1∶1∶1,进行常规管理。选择生长一致的健壮苗木于8月6日开始进行0、2、4、6、8 和10 d淹水处理,共持续10 d;将植株放入长、宽、高分别为120 cm×90 cm×34 cm的塑料水箱中,注水至没过土壤表面2~3 cm,上方搭建避雨棚,保证棚内通风透光;处理期间及时补水,保持水面高度。每处理5株,重复3次。
1.2 根系细胞学分析
分别取淹水处理后0 、4 、8 、10 d植株细根系,参照关超[15]的方法将根小心剪成0.5 cm左右根段,放入FAA固定液中固定12 h,于4 ℃条件下保存。经过洗涤、脱水、透明、透蜡、包埋后用全自动切片机(Leica255,德国)进行切片,用番红-固绿双染色法进行染色,封片后用显微镜(OLYMPUS BH-2,日本)观察细胞结构。
1.3 植株形态观察
每天观察植株形态变化,记录不同品种的植株生长状态。对植株涝害情况划分等级[16-17]:1级:芽叶受害率小于5%;2级:芽叶受害率6%~25%,个别枝梢萎蔫干枯,受害较轻;3级:芽叶受害率26%~50%,少数枝梢萎蔫干枯,受害中等;4级:芽叶受害率51%~75%,较多枝梢受害、萎蔫枯死,受害较重;5级:芽叶受害率76%以上,大部分枝梢受害、萎蔫干枯,甚至整株死亡,受害重。
根据统计数据,按以下公式计算涝害指数:

1.4 叶片光合指标测定
分别在淹水后0 、2 、4 、6 、8 、10 d用便携式光合仪(Li-6400,美国)在晴天的上午9:00~11:00测定各处理植株功能叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci),选择新枝条中部成熟叶片(自顶芽向下第5~8片),每株重复测定3次,每品种测定5株。
1.5 叶片生理生化指标测定
分别取各处理植株中上部成熟叶片,用去离子水清洗后擦干,参照刘新等[18]的方法测定叶片生理生化指标,其中电导率仪法叶片相对电导率(REC),蒽酮比色法测定可溶性糖(SS)含量,硫代巴比妥酸比色法测定丙二醛(MDA)含量,氮蓝四唑法测定SOD活性,愈创木酚法测定POD活性。
计算公式为:相对电导率/%=(初电导率-蒸馏水电导率) /(终电导率-蒸馏水电导率)×100, 伤害率/%=[1-(1-处理组相对电导率)/(1-对照组相对电导率)] ×100
1.6 数据统计分析
数据采用Excel 2010和SPSS 20进行处理与分析,显著性分析采用Duncan多重比较,利用Origin 2018作图。耐涝性综合评价参照周广生等[19]、白丹凤等[20]的方法,利用主成分分析和隶属函数分析比较品种间耐涝性差异,具体方法如下:

U(Xj)为每种指标的隶属函数值,式中Xj为第j个指标的测定值,Xmin、Xmax表示第j个指标的最小测量值和最大测量值,Wj为各指标权重,式中pj为各品种第j个主成分的贡献率;D值为涝害条件下各品种用隶属函数分析所得的耐涝性综合评价值,D值越大,说明该品种耐涝性越强。
2 结果与分析
2.1 淹水胁迫对植株形态的影响
淹水会造成植株叶片萎蔫黄化,甚至脱落。本试验中,淹水后3个品种叶片均受到影响,锦绣、新川中岛、春红分别在第4天、第5天、第7天出现萎蔫症状。淹水第10天锦绣植株水渍斑明显,叶片脱落严重;新川中岛中部和基部叶片黄化、脱落明显;春红叶片出现黄化并有少量脱落。3个品种植株均是老叶首先出现形态变化,说明老叶对淹水胁迫更敏感。根据形态统计涝害指数得到表1,可知随着淹水时间的延长,各品种涝害指数逐渐升高,淹水4 d各品种涝害指数上升较快,春红、新川中岛、锦绣淹水较前一天分别增加128.47%、112.08%和223.31%。淹水10 d,新川中岛、锦绣涝害指数分别达到70.67%、78.22%,春红仅为52.79%,显著低于另外2个品种。
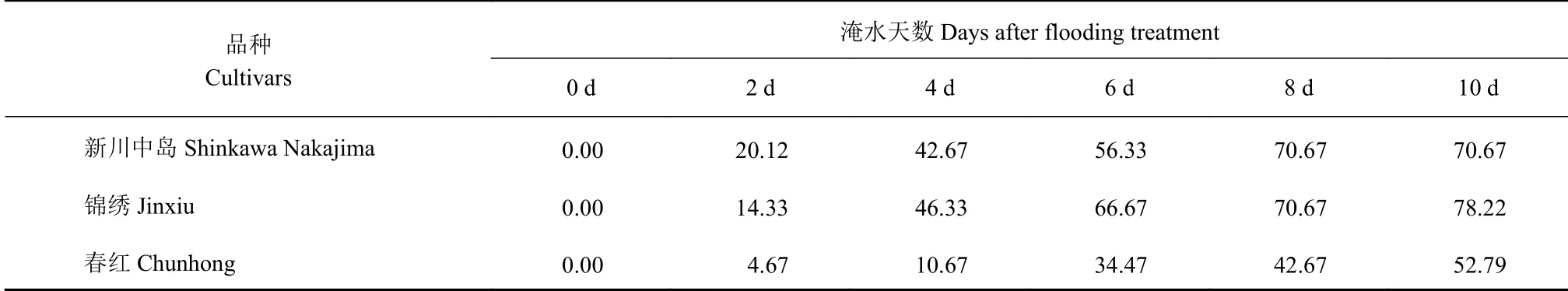
表 1 淹水胁迫下3个桃品种涝害指数Table 1 Waterlogging indices of 3 peach cultivars under flooding stress (单位:%)
2.2 淹水胁迫对砧木根系细胞结构的影响
根是淹水状态下植株最直接受到影响的器官,根系发育状况很大程度上会影响植株在涝害情况下的表现。通过砧木根系形态及细胞学观察,调查淹水对3个桃品种砧木根系的损伤程度以及砧穗互作对根系耐涝性可能的影响。淹水10 d所有品种根系根尖腐烂变黑,并无不定根产生。如图1所示,正常水分管理的植株根系细胞结构清晰,皮层细胞排列规则紧密(图1-A、E、I);随着淹水时间延长,皮层薄壁细胞扩大变形,皮层细胞与初生木质部细胞发生分离。淹水4 d,皮层细胞部分发生裂解,产生不规则气腔(图1-B、F、J),但细胞形状依然清晰。淹水8 d,皮层细胞增大,初生木质部与皮层薄壁细胞发生分离,并发生不同程度裂解,各品种间砧木根系细胞结构无明显差异(图1-C、G、K)。淹水10 d,根系横切结构较未淹水植株变化明显,皮层薄壁细胞离解融合,部分气腔发生断裂相互贯通,形成大的空腔(图1-D、H、L)。结果表明,淹水对3个品种砧木根系的伤害程度基本一致,砧木根系细胞形态结构的变化没有明显差异。

图 1 淹水胁迫下3个桃品种根系横切解剖结构变化Fig. 1 Changes on root cross-section structure of 3 peach cultivars under flooding stress
2.3 淹水胁迫对叶片光合指标的影响
光合作用为植物生长提供能量,淹水胁迫抑制植物光合能力,降低植物光合速率。评价光合能力的主要指标有净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等。
由图2-A可见,淹水处理导致各品种叶片净光合速率降低,第4 天净光合速率下降最快,新川中岛、锦绣和春红分别下降72.38%、72.23% 和66.88%。第6天开始春红Pn显著高于新川中岛和锦绣,第10天锦绣Pn降至负值,说明此时有机物消耗量大于积累量。图2-B结果显示,淹水条件下3个桃品种叶片蒸腾速率与净光合速率变化趋势基本一致,处理第4天下降较快,春红和锦绣与对照相比均达到显著差异水平(P<0.05)。淹水第10天春红叶片Tr和锦绣接近,显著高于新川中岛。
气孔导度是淹水胁迫最为敏感的指标,也是影响植物光合作用、呼吸作用及蒸腾作用的主要因素。由图2-C可知,随淹水进行Gs呈下降趋势,气孔开闭受阻。淹水4 d,各品种蒸腾速率下降明显,春红、锦绣和新川中岛分别较对照下降了72.12%、81.85%和67.47%,新川中岛显著高于其他品种。如图2-D所示,淹水条件下叶片胞间CO2变化与Pn、Tr等变化趋势相反,随淹水进行总体呈上升趋势。0~4 d各品种植株Ci较对照变化不明显,品种间无显著差异;第6 天Ci明显上升,春红达到374.55 μmol·mol-1;第8 天锦绣达到446.04 μmol·mol-1,明显高于其他品种。
综上所述,淹水胁迫明显抑制桃植株光合能力,不同光合指标变化略有不同,各品种叶片Pn、Tr、Gs随淹水进行呈下降趋势,Ci则上升。淹水6 d,各品种间Pn、Tr、Ci出现显著差异,其中Pn尤为明显,淹水10 d极显著低于正常水分条件下植株,其中春红净光合速率最高,锦绣最低。总体来看,淹水胁迫下春红光合能力强于新川中岛和锦绣。
2.4 淹水胁迫对叶片相对电导率、可溶性糖和丙二醛含量的影响
淹水对植物细胞造成不同程度损伤,破坏细胞平衡状态,影响植物正常的生长发育。植物细胞膜对维持细胞的微环境和正常的代谢起着重要作用。由图3可见,淹水条件下,各品种植株叶片相对电导率(REC)较对照组明显升高,且随处理时间延长REC呈上升趋势。淹水6 d,锦绣叶片REC显著高于其他品种;第10 天,春红、锦绣和新川中岛分别是对照的4.55倍、3.60倍和2.92倍。
伤害率是指植物受到逆境胁迫之后相较于正常植株细胞膜受到损伤的程度,伤害率越大越容易对植物造成不可逆的损伤。由图3可知,随着淹水时间延长,不同品种叶片伤害率呈上升趋势,处理第10天,新川中岛、锦绣、春红叶片伤害率分别为63.92%、71.10%、59.98%,锦绣显著高于新川中岛和春红。
植物在逆境胁迫下,可通过调节渗透物质含量维持细胞内部稳定,减轻对植物的伤害。如图4,淹水胁迫进程中,各桃品种功能叶中可溶性糖(SS)含量呈波动变化趋势,其中新川中岛、锦绣呈先上升后下降趋势,春红呈先下降再上升后下降趋势;淹水第4 天,锦绣可溶性糖含量上升至2.09倍,达到300.40 μmol·g-1,显著高于其他品种;第10 天春红叶片SS含量显著低于其他品种,但高于初始值(P<0.05)。新川中岛、锦绣叶片丙二醛(MDA)含量呈先上升后下降再上升趋势,春红则呈先上升后下降,各品种MDA含量均在第2 天上升最快,分别上升162.68%、51.42%和52.93%,均显著高于各品种对照组(P<0.05)。春红在淹水4~6 d叶片中MDA含量显著高于新川中岛和锦绣,其他时间各品种间无显著差异。

图 2 淹水胁迫下3个桃品种叶片光合指标变化Fig. 2 Changes on leaf photosynthetic indices of 3 peach cultivars under flooding stress

图 3 淹水胁迫下3个桃品种叶片REC和伤害率变化Fig. 3 Changes on relative electric conductivity and damage degree on leaves of 3 peach cultivars under flooding stress

图 4 淹水胁迫下3个桃品种叶片SS含量和MDA含量变化Fig. 4 Changes on leaf SS and MDA contents of 3 peach cultivars under flooding stress
2.5 淹水胁迫对叶片保护酶活性的影响
超氧化物歧化酶(SOD)能保护生物体免受活性氧伤害,其活性与植物抗逆性及衰老有密切关系。植物受到逆境胁迫时,SOD酶活性会上升以保护植物细胞。本试验中,3个品种叶片SOD酶活性淹水条件总体呈现先上升后下降的趋势,但均显著高于初始值(P<0.05)。由图5可知,不同品种SOD酶活性变化速率不同,0~2 d锦绣叶片SOD酶活性增长约2.32倍;2~4 d春红上升较快,达到2.96倍。淹水6 d各品种SOD酶活性达到最大,约为170 U·g-1,但品种间无显著差异。

图 5 淹水胁迫下3个桃品种叶片SOD和POD酶活性变化Fig. 5 Changes on leaf SOD and POD activities of 3 peach cultivars under flooding stress
叶片过氧化物酶(POD)酶活性淹水胁迫下总体呈现先上升后下降趋势,淹水4 d各品种叶片POD酶活性达到最高,新川中岛、锦绣和春红分别为84.70 U·g-1·min-1、67.83 U·g-1·min-1和105.36 U·g-1·min-1,春红显著高于其他品种(P<0.05)。淹水6 d,POD酶活性开始降低,新川中岛、锦绣、春红分别下降39.43%、34.77%、57.67%,各品种间差异不显著。
淹水过程中,桃叶片细胞膜透性增大,REC和伤害率上升,MDA和SS含量呈现先上升后下降趋势。处理初期通过SOD、POD等保护酶活性显著升高调节ROS平衡,后期随着淹水时间的延长,植株受损加重导致调节功能失灵,保护酶活性降低。
2.6 3个桃品种耐涝性综合评价
在试验中发现淹水对桃植株造成不同程度损害,但各品种单一指标变化存在差异,因此不能通过单个指标判断3个不同品种耐涝性强弱。根据光合参数、REC等9个单项指标计算耐涝系数。由表2可知,同一品种不同指标或者不同品种同一指标α值大小各不相同,认为α>100%为上升,α<100%为下降,其中各品种REC、Ci、MDA、SS、SOD为上升,Pn、Tr、Gs下降,POD在新川中岛、锦绣表现下降,春红表现上升,不同指标上升或下降幅度不同。对以上9个指标进行主成分分析,如表3,得出F1、F2贡献率为70.69%和29.30%,累计贡献率99.95%,说明F1、F2包含以上9个指标几乎全部信息,可用这2个指标作为主成分对3个不同桃品种进行耐涝性评价。

表 2 淹水胁迫下3个桃品种各单项指标耐涝系数Table 2 Waterlogging tolerance coefficients on various indices of 3 peach cultivars under flooding stress (单位:%)

表 3 淹水胁迫下3个桃品种各指标系数及贡献率Table 3 Coefficients and contribution rates of various indices of 3 peach cultivars under flooding stress
以所有指标为原始数据,对其进行隶属函数分析,求得综合指标值F1、F2,隶属函数值U(F1)、U(F2),耐涝性综合评价值D值。由表4可知,对F1来说,U(F1)最大值为1;对F2来说,U(F2)最大为1,结合耐涝性综合评价值D, 3个品种中春红耐涝性最高,新川中岛耐涝性中等,锦绣耐涝性最差。

表 4 淹水胁迫下3个桃品种耐涝性综合评价Table 4 Comprehensive evaluation on waterlogging tolerance of 3 peach cultivars under flooding stress
3 讨论与结论
淹水胁迫影响植物的生长发育,根系是植物淹水最直接受到影响的器官[21]。耐涝植物往往通过形成气腔和不定根来维持缺氧环境中根系O2、CO2等气体扩散,缓解淹水对植物的损伤[22-23],如耐淹猕猴桃砧木具有更发达的通气组织和不定根[14,24]。本研究中,淹水导致不同品种桃砧木根系均腐烂受损,皮层细胞裂解,形成气腔,但未观察到不定根的发生,各处理间差异不显著。然而,植株形态和光合生理特性结果表明,相同砧木条件下3个不同熟期桃品种之间存在耐涝性差异。有报道认为接穗对砧木的作用也会影响植株抗逆能力,如Sun等[25]以平邑甜茶为砧木分别嫁接粉红女士和秦冠,结果粉红女士表现对水分胁迫更加敏感,秦冠则表现较强耐旱性,这与本研究结果相似。郭学民等[26]用毛桃嫁接21世纪,发现毛桃根系导管分子的类型与直径发生改变,植株表现不同的抗逆性及生长状态。更多的研究表明接穗和砧木之间的亲和性、细胞识别和信号交流等会影响植株根茎中导管直径、初生木质部大小,进而影响植株抗逆能力和生理状态[27-28]。虽然有报道接穗在一定程度上能够调控砧木根系的生长发育并产生反馈作用,但目前关于接穗对砧木抗性影响的机制尚存争议,其发生机理还需进一步深入研究。
叶片是植物最敏感的器官,逆境胁迫下叶片往往最先表现症状[29]。淹水造成的缺氧环境影响叶片气孔开闭,阻碍CO2的扩散运输,降低光合速率[30-31],这在桃砧木品种上也得到体现[32]。本研究中,淹水处理导致3个桃品种光合性能严重受损,甚至淹水10 d后锦绣的Pn降为负值。叶片保护酶系统可抵御逆境胁迫时活性氧自由基的毒害作用,是植物适应淹水胁迫的重要指标,研究表明耐淹植物保护酶活性显著高于涝敏植物[33]。本研究中,淹水处理下桃叶片抗氧化酶活性总体呈现先升后降趋势,表明受涝初期植株启动抗氧化系统清除自由基,调节细胞ROS系统,但随着处理时间延长,涝害造成细胞严重损伤,导致保护酶系统被破坏,3个桃品种保护酶活性均出现下降,这与张慧琴等[34]的研究结果基本一致。
植物的耐涝性主要体现为淹水后植株的适应性和抵抗能力。适应性主要有形态学水平和生理生化水平的适应,包括不定根和通气组织的产生,以及光合呼吸代谢、渗透调节和抗氧化机制的改变等。由于自身抗性机制的差异,不同品种在逆境条件下对具体指标响应强度不一,利用单一指标和统一标准难以对抗逆性做出正确评价。综合多指标的隶属函数法能克服单一指标评价的缺点,已被广泛应用在桃[8]、猕猴桃[20]、番茄[35]及牡丹[36]等多种植物上进行耐涝性评价。本研究中通过主成分及隶属函数分析得到的耐涝性综合评价值D大小顺序与3个品种在植株形态、净光合速率、涝害指数等方面的表现基本吻合,但与其他测定指标数值不尽一致。这与马瑞娟等[8]在不同桃砧木品种耐涝性评价研究中的结果类似,推测植株在淹水胁迫下生理机能的调节与表观适应性的变化不同步所致。
目前南方桃产区普遍以毛桃作为砧木嫁接不同栽培品种,但毛桃砧木的耐涝性表现并不突出,生产上需采取相应的技术措施来减轻洪涝危害,从而增加了人力和物力成本。桃涝害防御除了综合技术的集成外,选择抗性砧木和品种也非常重要,可以通过综合评价筛选出合适的材料供生产上推广利用。因此从品种源头考虑,桃涝害的防御除了加强耐涝砧木的选育外,也应根据不同区域的栽培条件和市场需求选择耐涝性强、砧穗亲和力好的优良桃栽培品种,以增强桃植株的抗涝能力、提高桃经济寿命和产业效益。
综上所述,相同砧木的早熟桃春红、中熟桃新川中岛和晚熟桃锦绣3个品种间耐涝性存在差异,其中春红耐涝性表现最好。在应对涝害胁迫时,耐涝桃品种具有较高的光合效率、渗透调节能力和抗氧化酶活性,来缓解低氧伤害,适应淹水环境。生产中遇强降雨等涝害天气时,应及时排涝,易受涝产区应选择耐涝桃品种和砧木。
