应用环境筛与生殖力和种群增长模型研究钟萼木致濒因子
2022-03-08郭祥泉洪伟涂年旺郑经池吴炜程志强
郭祥泉,洪伟,涂年旺,郑经池,吴炜,程志强
(1福建省三明市林业局,福建 三明 365000;2福建农林大学 林学院,福建 福州 350002;3福建省永安龙头国家湿地公园管理中心,福建 永安 366000; 4福建省永安市林业局,福建 永安 366000;5福建省永安林业(集团)股份有限公司,福建 永安 366000)
生物多样性能增强生态系统稳定性,森林物种多样性可提高森林生态系统的生态效益,有效减少或消除自然灾害,如干旱、洪涝灾害、局部小气候危害等,保护森林物种多样性能更好的使森林生态系统进行良性循环。由于人们对大量木质材料及其衍生物的应用,造成许多树种的灭绝或处于濒危状态,森林多样性减少,使森林生态系统功能减弱,增加了许多自然灾害,给社会带来巨大的损失。研究濒危树种是为了更好地保护生物多样性,以提高森林生态系统的生态功能,改善人居环境的生态安全。对濒危树种的保护不应是简单禁止人为使用或减少人为干扰,使物种去濒危的最有效方法是通过人为措施促进物种生殖,增加种群数量。利用林木种群的生态学研究物种濒危因子,可为人工促进物种生殖提供更好的研究方向,加速对物种的保护与利用[1-4]。钟萼木(BretschneiderasinensisHemsl.)原分布于长江流域及以南等省的海拔500~2 000 m中、低山地带,因具有木材利用、药用等多用途特性,以及人为干扰与该树种生长习性,导致该树种处于濒危状态[5-14]。目前已有一些学者对该树种人工繁育等进行研究,但该树种的种质资源缺乏、生长习性的遗传多样性分化,较大程度限制了人工培育的发展,对该树种濒危生态学与致濒危主要原因的研究,能为该树种的去濒危与保护提供更多可实施办法[9-17]。本研究试图通过对该树种的濒危趋势与主要引起该物种致濒因子调查与分析,以揭示该树种在天然更新过程的生态习性,为科学制定保护措施与人工栽培提供理论依据。
1 研究区域自然条件
试验地分布于福建省永安天宝岩自然保护区、福建省闽江源自然保护区。福建省永安天宝岩自然保护区地处于戴云山西北延伸,为中亚热带南部季风湿润气候,土壤类型为红黄壤,多数土层由中厚到深厚,垂直分布为175~1 700 m,多年年均降雨量为1 578 mm,年平均气温16 ℃,1月平均气温8 ℃,7月平均气温28 ℃。福建省闽江源自然保护区处于武夷山脉西南,属中亚热带北部季风湿润气候区,气候温暖,雨量充沛,四季分明,冬季常有积雪与结冰。全县年平均11~17 ℃,日均温≥10 ℃的年积温4 800~5 100 ℃,无霜期260~265 d。全县年平均降雨量1 720~1 910 mm,年蒸发量1 347.4 mm。
2 研究方法
2.1 试验调查
本试验采用设置方块标准地进行调查,样地为25.82 m×25.82 m,永安市天宝岩自然保护区设置18个样地,建宁闽江源自然保护区设置21个样地。钟萼木的种子较大,自然传播距离较短,主要分布在种子掉落区,该树种种子是许多动物的食物,动物搬运种子扩大种子传播距离,加大分布范围。样地选择以钟萼木结果母树为调查中心点,在结果母树的种子掉落区设置调查样地,每株母树坡上方设置1个样地,在母树坡下方设置2个样地,坡上、下方相邻样地部分重复边界线。在样地内调查冬季果实掉落数量与分布、次年钟萼木新萌芽的幼苗以及多年生长的每棵小树高、胸径(或地径)、年龄,根据该树种芽痕计算年龄。调查所在样地内群落的乔木层、灌木层与草本层的主要种类,估算样地郁闭度、各层物种的盖度、平均高,按建群种划分群落类型。样地调查选择在该树种种子成熟期种子刚刚脱落不久时,调查样地内种子的密度,另捡取一定数量种子,测定种子千粒重与发芽力。6月份调查种子萌发后小苗生长数量,11月上旬调查当年新萌芽小苗与年末小幼苗保存数量。分别于2017年11月上旬调查种子密度与多年生小树生长,2018年的6月中旬与2018年11月上旬调查当年新萌芽小苗与年末小幼苗保存数量。
2.2 研究方法
本研究应用分株编制种群静态生命表,应用环境筛分析种群在群落竞争中的生存情况;应用种子更新编制生殖力表,通过种子生殖与生殖生态分析种群增长力与影响种群增长的主要因素,应用因子变量的线性回归模型分析影响种群增长过程的影响因子,筛选环境对该树种种群增长影响的主要因子。
3 结果与分析
3.1 钟萼木种子生态与生殖生态分析
根据对该树种的野外调查与人工培育结果,人工培育见参考文献12、13,该树种种群天然更新趋于濒危的主要原因在于种子安全保存、种子的深休眠和当年生幼苗生长保存率。该树种种子的千粒重达700~900 g,属于大粒种子,富含淀粉,是许多动物的越冬食物;种子成熟脱落后在较短时间段内易被多种动物作为食物所掠取,被储藏于某些动物的巢穴中,不能被用于天然更新[12-13]。野外调查结果表明,天然脱落的种子经过一段时间后,大部分种子消失,少部分种子被落叶等凋落物覆盖而被保存下来。该树种种子外壳坚硬,具有蜡质层,不易透水,属于生理后熟的深休眠类型,种子需经休眠处理才能萌芽或部分萌芽,所以,天然条件下林下自然更新的幼树分布量较少[9,12]。利用种子更新生殖编制生殖力表见表1,表内的X、lx、mx、lxmx、xlxmx中的X代表存活小树年龄,lx代表X年龄时个体存活率,mx代表X年龄时平均每一母树林下更新小树的存活株数,R0为种群净增殖力,rm为内禀增长率,T为世代平均周期,λ为周期增长率[1-4]。

表1 钟萼木种群种子生殖力表Table 1 The seed fecundity schedule of Bretschneidera sinensis
由表1可知,该树种结实有明显的大小年,1次丰年结果后会出现2~4年的小年,该树种母树结实的大小年现象特别显著,影响种群增长;种群净增殖力为5.898 4,内禀增长力为0.849 6,表明净增殖率很小,种群依靠种子进行自我更新,个体在自然竞争中种群不断变少,自然竞争力较弱。
3.2 环境筛分析
物种在自然进化过程中各自形成一种适宜生存的竞争机制,由无性生殖向有性生殖进化有利于物种产生多样性个体以适宜不同环境,使物种在变化环境中保存下来。物种生殖机制与物种个体生长机制共同作用形成的一种适宜生存优势特性,所形成的生殖机制与生存机制成功与适宜生存的进化优势的量度称作适合度[1-4]。某一物种在生活史的某一阶段中,存在影响或限制物种生存或生殖的环境因子,限制种群增长的因子称为环境筛,环境筛是环境因子强烈筛选物种生活史阶段起决定作用的影响因子,用以确定某些因子是某一物种致濒阶段和产生致濒因子,直接影响物种生存。
钟萼木为多年生木本植物,采用“空间代替时间”方法编制静态生命表,静态生命表的编制内容分X表示年龄级;lx表示X年龄开始时标准化存活数;dx表示从X到X+1年龄期的标准化死亡数;qx为X年龄级的个体死亡率,qx=dx/lx×1 000;Lx表示从X到X+1年龄期时的平均存活的个体数,Lx=(lx+lx+1)/2;Tx表示X年龄级及以上各年龄个体存活总数,Tx=lx+lx+1+…;ex表示进入X年龄级个体的平均生命期望,ex=Tx/lx;ax表示X年龄级开始时的实际存活数;Kx表示种群消失率,Kx=lnlx-lnlx+1,根据调查结果编制的静态生命表,见表2[1-4]。

表2 钟萼木静态生态表Table 2 The static life table of Bretschneidera sinensis
由表2可知,静态种群生命表可以分析物种在生命史中出现的个体死亡率或消失率较高阶段,在高死亡率或消失率阶段就是受环境影响较大的时期,该时期就是受环境筛强烈作用阶段,物种自我更新的缺失或受环境作用的结果。钟萼木的死亡率或消失率高的阶段,该物种种群消失率阶段在第1、2年。主要是种子不能处于安全状态下进行自我更新、种子深度休眠与幼苗期存活率低的影响,种子由母树脱落后,大量种子在休眠状态下被某些动物作为食物而消失,使该树种缺少足够多种子数量来完成天然更新。根据人工培育结果,该树种种子属于深度休眠类型,幼苗期生长易受病虫害危害,在自然条件下,幼苗保存率低于人工培育,天然更新幼树保存率较低[12-13]。该树种在第1、2年生幼树生长阶段,受环境因子如病虫害、水分、光照及动物的干扰而消失,该阶段是该树种种群消失的重要阶段,改善该树种在第1、2年生长阶段的环境因子,可有效提高幼树保存率,促进种群的增长。
3.3 钟萼木种群生长过程中关键因子分析
种群增长过程受种群结构、种群发育和环境因子等综合影响,根据野外调查与人工培育的观察结果,筛选了影响钟萼木生物量增长(Y)的几个主要因子,郁闭度(X1)、植被盖度(X2)、土壤湿度(X3)、坡度(X4)、种群密度(X5)、种群年龄(X6),前4个因子为定性因子,定性因子根据不同强度分成3个水平进行量化处理,种群密度与种群年龄为定量因子[1-4]。根据影响种群生物量增长因子的相关数据,定性因子见表3。
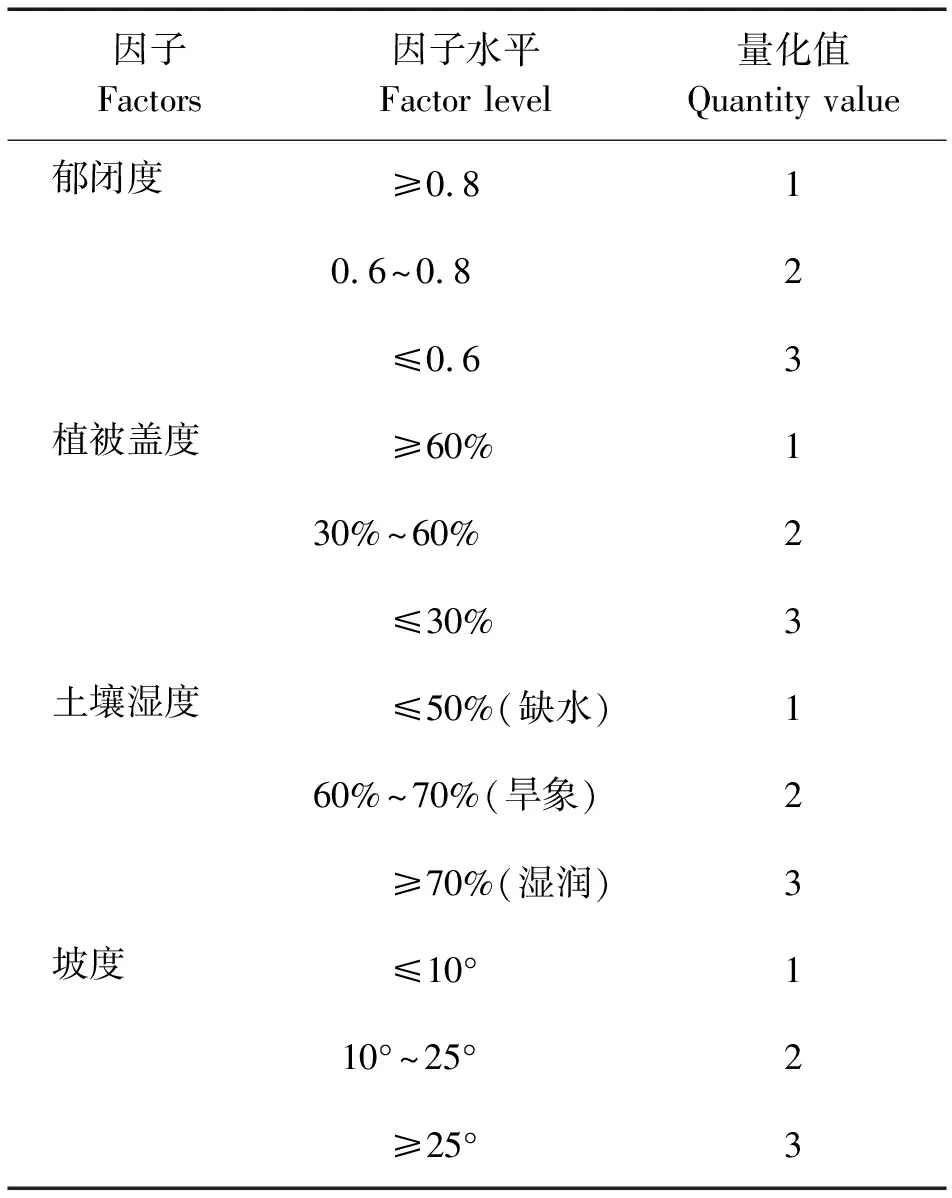
表3 不同定性因子量化值Table 3 The quantity value of the different qualitative factor
由表3可知,经DPSS10版软件处理,种群生物量增长与各因子变量的线性回归模型如下:
Y=-2.872 4+0.265 7X1+0.587 9X2-0.405 7X3-0.478 2X4+0.287 9X5+0.157 8X6
R2=0.987 8(n=8,P<0.01)
回归模型的相关系数R2为0.987 8,达显著水平,表明方程模拟适用[12]。种群生物量增长与各因子变量的偏相关系数都达到极显著水平,根据各偏相关系数值大小排序为种群年龄(r=0.992 1)>郁闭度(r=0.987 6)>土壤湿度(r=0.964 5)>植被盖度(r=0.954 2)>种群密度(r=0.922 8)>坡度(r=0.915 8)。根据调查与前期研究发现,该物种生长过程,早期在林下自然更新的小树不易生长侧枝,许多小树生长至10多米高才有少数侧枝,人工培育小树要生长2.0 m以上才开始出现侧枝生长[11]。调查结果发现,生长30多年个体,树高仅10多米、胸径小于10 cm,侧枝小而少或未有侧枝生长,也未见开花结果。能进行结果繁殖的个体树龄都较大,当树冠处于林分上层或亚林层状态下才能有效开花结果,该树种的开花结果年龄明显影响种群增长。该树种所处林分类型直接影响结果母树的初始结果年龄与结果数量,群落中优势种群郁闭度大,抑制该树种个体生长,个体间空间竞争加剧,使结果母树分枝少,冠幅小,受光面积小,导致母树结果量少,影响种群增长;土壤湿度影响种子在冬季的催芽与休眠处理,影响次年种子萌发,冬季降雨量影响土壤湿度,直接影响种子的催芽处理。植被盖度影响更新小树的生长,延迟种群初始结果年龄。种群密度与坡度影响种子的分布密度,从而影响种群增长。在自然条件下,该物种的生长特性与结果初始年龄极大的限制了种群的增长,通过主要影响因子分析研究,可以通过人工促进改变该树种种群增长方式。认为天然保护,较难使该树种种群得到较快增长,人工培育能有效促进该树种的种群增长,目前受种质资源影响与人工繁殖的技术限制,大量人工培育还受限制。
4 小结
一个树种处于濒危状态是由内在的生物学特性、生态习性以及外在的人为干扰等因子综合作用的结果,通常一个树种易导致濒危状态与他的天然更新能力有极大关系,特别的更新模式是一个物种易产生濒危的重要原因[1,5,6]。钟萼木是一种单科单属的孑遗树种,生长特性、天然更新能力都存在独特性,它的生长特性与人为的干扰使该树种处于濒危状态[5,6,9]。
根据不同分布点调查结果,在人为活动较频繁的分布点,小树保存数比较少,人为干扰多的地方种群数量随着时间延长林分小树保存数量明显少于未受干扰的分布点,人为因子的作用极大的影响种群增长。
根据作者等对该树种人工培育研究结果,种子采用赤霉素处理再进行层积催芽可较好的解除休眠,种子有较好的发芽效果,研究见参考文献9、12、13。人工培育1年生苗木的生长表现显著的高生长性状遗传多样性与早期的生长分化,具体见文献18、19。生长类型权重表现为平均值及相邻2个生长类型权重比例占全部的85%以上,较好生长类型约占5%~6%,优良生长类型少于1%[16-17]。多年生人工培育的个体表现为2.0 m以下主干基本不分枝,平均年高生长量约在40~50 cm,优良个体高年生长量可达1.0 m[11]。人工造林3年后生长分化与优良个体生长表现为好的遗传品质个体具有速生效果,有潜在的栽植价值,具体见文献16、17、20。
钟萼木致濒成因是内在与外在因素作用的结果,内在因子主要包含生殖能力、生殖模式、抗逆性、竞争能力等;外在因素主要包含环境条件、人为干扰等。种子的深休眠与幼树的抗逆性差是该树种的内因产生结果,种子安全岛缺失是外在因子影响的结果,内外因子综合结果降低了该树种天然更新能力。通过编制种子更新生殖力表,应用环境筛分析种群在群落竞争中的生存竞争力,分析种子生殖与生殖生态对种群增长力与影响种群增长的主要因素,认为该树种幼树第1、2年生长环境条件对幼树生长产生较大影响,种子休眠、安全岛缺失、种子深休眠抑制种子萌发等导致该物种种群增长力差,使种群在自然竞争中处于弱势状态,是濒危的重要原因。
由生境因子与种群动态增长模式研究表明,对种群增长产生影响的因子为种群年龄>郁闭度>土壤湿度>植被盖度>种群密度>坡度;该树种种群结实年龄对种群增长影响最大,该树种在天然更新过程,幼树生长期长,许多30多年生个体胸径生长还不到10.0 cm,没有分生侧枝或少分生侧枝,树冠很小,处于林分下层,不能开花结果进入生殖阶段,生活史较其他乡土树种要长许多,许多结果母树年龄都是百年以上,且该树种结果大小年明显,间隔期通常都达2~4年,显著影响自然繁殖更新。生活史长、结实年龄大及结实大小年显著是树种的内因,该树种所处林分郁闭度、土壤湿度、植被盖度、种群密度、坡度是外在环境因子,二者共同作用影响种群增长。
经内外因子研究结果表明,该树种内在影响因子作用是主要的,内在因子还造成现有种质资源的缺乏,使人工培育的快速发展受到限制。该树种无性繁殖属于难生根、萌芽少的树种,给无性繁殖带来困难。
