钱塘江干流夏季浮游植物群落结构特征及其对水文气象的响应*
2022-03-05国超旋权秋梅姚建良王吉毅叶小锐朱梦圆孙起亮朱广伟
张 萍,国超旋,俞 洁,权秋梅,姚建良,王吉毅,叶小锐,朱梦圆,孙起亮,朱广伟
(1:西华师范大学环境科学与工程学院,南充 637002) (2:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京 210008) (3:浙江省生态环境监测中心,杭州 310012) (4:杭州市桐庐县环境保护监测站, 桐庐 311500) (5:哈尔滨师范大学生命科学与技术学院,哈尔滨 150025)
浮游植物是河流生态系统中的初级生产者,在信息传递、物质循环和能量流动过程中发挥着不可替代的作用. 浮游植物群落结构与河流水环境因子之间会相互影响[1],其可以快速地响应环境变化,能够较好地指示河流水质状态[2-4]. 浮游植物的异常增殖会引发藻类水华,对河流生态系统的结构和功能造成严重危害,威胁着沿岸渔业、旅游业的发展[5],因此,阐明浮游植物群落结构动态特征及影响因素在河流生态系统管理中至关重要[6].
浮游植物在河流中的生长既受到营养盐限制,也受水文气象条件的显著影响. 营养盐是浮游植物生长的基础[7],气象与水文动力学过程组合对浮游植物群落结构的变化和水华的快速形成具有重要影响[8]. 例如,降雨能将营养盐带入水体,改变藻类的生长环境,造成浮游植物群落结构组成的变化[9-11];水体流量影响着浮游植物功能群和生物量的变化[12];水库入库流量、水位[13]和水体稳定度[14]等是水华形成的关键因素;全球变暖有利于蓝藻在水生生态系统中占优势[15],极度干旱影响水动力条件,进而对水华的形成构成显著影响[16].
钱塘江干流不仅是重要的水源地,还是开发可持续发展渔业资源的重要水域,水域环境背景条件和水质状况复杂多变[17]. 2004、2016和2017年钱塘江干流的富春江段均发生过蓝藻水华[18]. 水华的暴发不仅仅对旅游业造成一定冲击,也对饮用水安全构成严重威胁. 鉴于此,阐明钱塘江干流浮游植物群落结构的时空特征和影响因素,对流域水质安全保障尤为重要.
盛海燕等[19]利用2006-2007年20次富春江水库的调查数据,发现富春江水库浮游植物群落结构、组成及密度受水文、水温、营养盐等环境因子的显著影响;谷雨等[20]在钱塘江(杭州段)4个断面用野外藻类测定仪(BBE)测定,分析不同门藻类的季节变化特征及其与水温和总磷的关系;赫雅宾等[21]对富春江下游夏季浮游植物群落结构进行调查,认为氮是影响浮游植物的主要因子;国超旋等[22]分析极端天气对富春江水库蓝藻水华暴发的促发作用;吴挺峰等[23]通过数值模拟推算,认为当水库日均下泄流量超过600 m3/s以上,能够有效预防水库水华暴发;王婕等[24]初步探究了富春江水库变动回水区蓝藻水华的成因. 此外,还有少量关于钱塘江河口浮游植物分布和蓝藻水华的研究[25-26]. 然而,上述研究虽然表明水文气象因子是富春江水库浮游植物群落结构的重要影响因子,但是缺乏在较宏观尺度上对不同江段浮游植物群落结构、组成和细胞密度的对比分析与探讨,也未见有关浮游植物对水文气象过程动态响应的详细观测研究. 因此,本研究选取了浮游植物群落结构演替剧烈的梅雨季及之后的高温晴热时段,在钱塘江干流的11个断面采样进行浮游植物和水质物理化学指标监测,并结合同期水文气象数据,探究水文气象过程及河流筑坝影响下钱塘江干流浮游植物群落结构时空变化特征及关键驱动因子,以期为钱塘江流域的水环境管理提供科学依据.
1 材料与方法
1.1 研究区域概况
钱塘江流域(28°0′~30°30′N, 117°30′~120°30′E)位于浙江省西部,发源于安徽省休宁县,主要支流有衢江、金华江、兰江、新安江、富春江和钱塘江,属于亚热带季风气候,年均降雨量980~2000 mm,年均气温15~18℃[27]. 受梅雨控制,6月中下旬到7月上旬常有连绵阴雨,并伴有暴雨[28].
富春江水库地处钱塘江干流兰江与新安江交汇后的江段,是一个过流型水库. 出库流量主要受控于上游兰江自然河道的径流量,以及新安江水库的下泄流量,库区段多年平均水域面积为56 km2[19],平均库容为4.4×108m3[29]. 富春江水库是桐庐县的主要供水水源,其富营养化程度与水质安全备受关注.
1.2 样品采集与数据来源
于2020年夏季6-8月在钱塘江干流设置11个监测点位(图1)进行样品采集,其中S1位于衢江,S2位于金华江,S3~S4位于兰江,S5位于千岛湖下游新安江,S6~S9位于富春江水库库区,S10~S11位于富春江水库下游. 其中,S5(新安江)6月数据缺失.
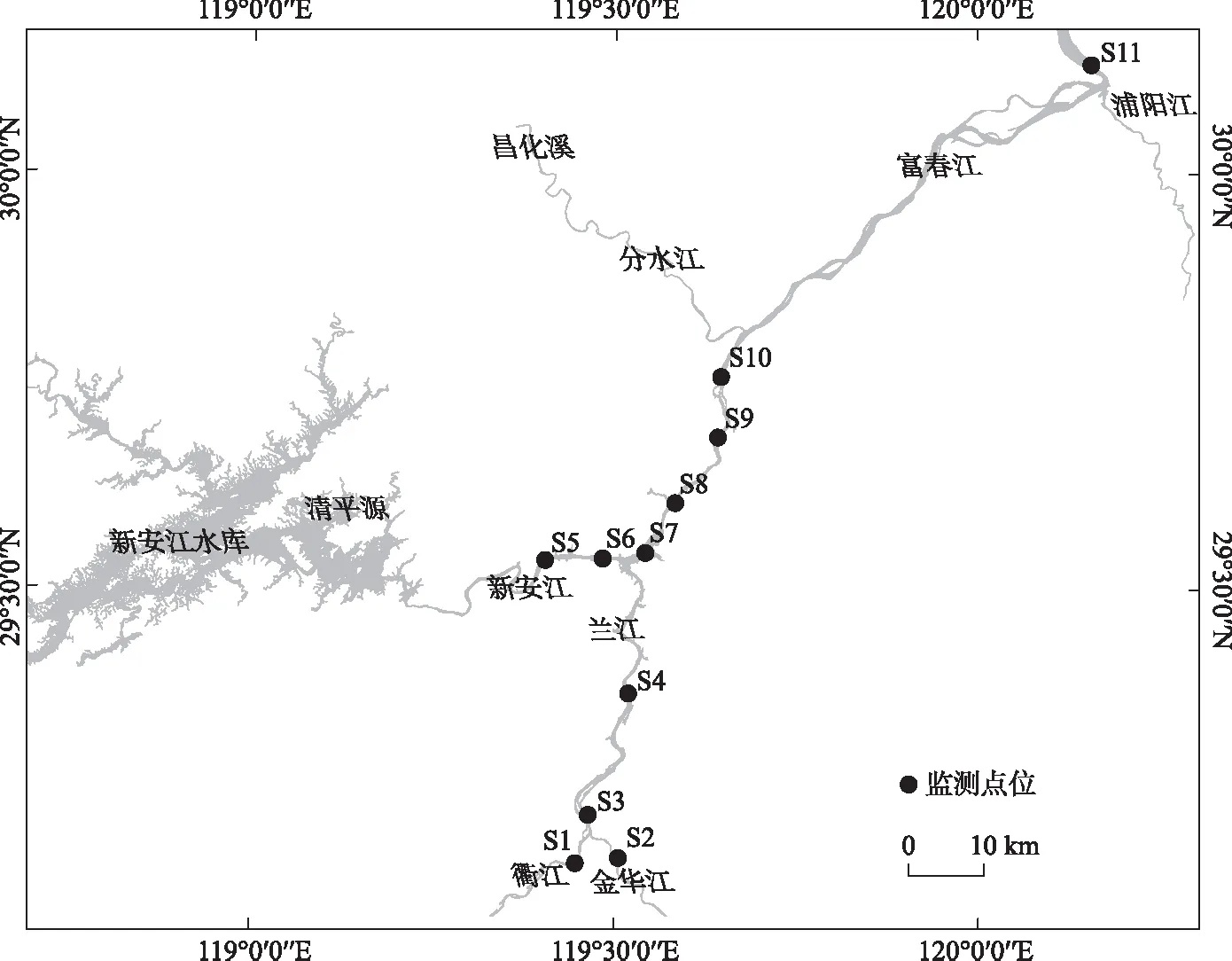
图1 钱塘江干流采样点位(S1: 衢江;S2: 金华江;S3: 兰江;S4: 将军岩;S5: 新安江;S6: 梅城大桥; S7: 三都大桥;S8: 子胥渡;S9: 严陵坞;S10: 桐庐水厂;S11: 闻家堰)Fig.1 Sketches of sampling sites in Qiangtang River
使用YSI公司的EXO2多参数水质分析仪原位测定水温(WT)、叶绿素a(Chl.a)、浊度等水质指标,并现场查看EXO数据,选取叶绿素a最大层,采集该层的浮游植物样品及水样. 取1.5 L水样立即冷藏,带回实验室测定总氮(TN)、总磷(TP)、溶解性总氮(DTN)、溶解性总磷(DTP)、溶解性有机碳(DOC)、高锰酸盐指数(CODMn)、有机悬浮颗粒物(OSS)等水质指标. DOC指标采用总有机碳测定仪(multi N/C 2100)测定,其余参数测定方法参照文献[30].
浮游植物定量样品的采集,使用有机玻璃采水器取水样1 L,置于样品瓶中,现场用1%(体积比)鲁哥试剂固定,带回实验室静置、沉淀48 h后,利用虹吸法将上清液吸除,定容至30 mL,存放在样品瓶中. 浮游植物计数时,吸取0.1 mL水样于浮游植物计数框中,在40倍物镜下,观察100个视野. 浮游植物鉴定和计数使用光学显微镜OLYMPUS-BX53. 浮游植物种类鉴定参照《中国淡水藻类——系统、分类及生态》[31].
新安江出库和富春江出库流量分别来源于新安江电站和富春江电站,兰江径流量根据区间占比计算而出. 同期气温、日照时数、降雨数据来源于国家气象科学数据共享服务平台杭州站监测数据(http://data.cma.cn).
1.3 数据处理与分析
累积降雨以及流量、气温和日照时数平均值使用2次采样间隔的数据作为当月值,其中6月使用采样前31 d的数据进行分析. 降雨量等级的划分依据国家气象局的降雨量等级划分标准,24 h内25~49.9 mm为大雨;50~99.9 mm为暴雨.
根据浮游植物在不同采样点的优势度(Y)来判定优势属[32]:
Y=(ni/N)×fi
(1)
式中,ni为i种的浮游植物细胞个数,N为所有种类浮游植物总细胞个数,fi为第i种浮游植物出现的频率,当Y>0.02时,该种类即为优势属.
换水周期计算公式为:
T=W/Qu
(2)
式中,T为换水周期(d),W为多年平均蓄水量(m3),Qu为多年平均出库流量(m3/s).
首先进行Kruskal-Wallis检验以探究6-8月营养盐和浮游植物细胞密度的差异性. 其次,进行Wilcoxon秩和检验以探究2020年与历史同期水文气象因子间的差异,以上方法在SPSS 22.0进行. 最后,运用CANOCO 5.0软件分析浮游植物群落和环境因子的关系,将浮游植物细胞密度进行Hellinger转换,将环境因子(除pH外)进行lg(x+1)转换,在排序前对浮游植物细胞密度进行去趋势对应分析(DCA),检验结果显示排序轴最大为2.91<3,故选用冗余分析(RDA). 通过共线性检验排除了水温、浊度、新安江出库流量、富春江出库流量和兰江出库流量,采用蒙特卡洛置换检验选出显著影响因子(P< 0.05)进行RDA分析,本文进行排序的物种数据满足至少在一个点出现密度大于2%.
数据处理及图形绘制使用 Excel 2010、Origin 2018及ArcGIS 10.2软件完成.
2 结果与分析
2.1 水文气象因子变化特征
调查期间,钱塘江干流的水文气象因子变化特征如图2所示. 从图可以看出,降雨量的月间差异显著,6月和7月总降雨量显著高于8月. 其中,7月共发生4场暴雨,总降雨量达到476.7 mm. 然而,8月连续19 d无降雨,该月总降雨量也仅为32.5 mm. 2020年夏季与2016-2019年同期相比,6、7月月降雨量分别增加了68.7和182.4 mm,其中7月累积降雨量为前4年均值的1.6倍;而2020年8月累积降雨比历史同期减少了96.6 mm,仅为前4年同期的17.38%(表1).
受降雨的影响,月间流量相差很大. 7月兰江径流、新安江出库流量和富春江出库流量显著高于8月,7月平均流量分别为2138、4285、1842 m3/s,8月平均流量分别为354、655、1046 m3/s. 受流量影响,富春江水库8月换水周期为5.77 d,而6月和7月分别为1.76和1.32 d. 与2016-2019年同期相比,7月新安江出库和富春江水库平均流量分别为历史同期的4.14和1.94倍.
气温在月间差异明显. 8月日照充足,连续12 d日照时数大于10.7 h;8月气温明显高于6、7月,连续15天平均温度高于30.9℃,平均温度为(30.6±2.6)℃. 与前4年的同期均值相比,2020年7月平均气温显著低于历史同期(P<0.01),为(25.9±2.5)℃,而6月和8月温度相较历年无显著差异(P>0.05).
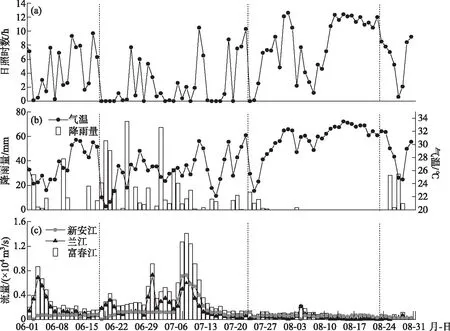
图2 2020年夏季钱塘江水文气象因子逐日变化图(图中虚线代表采样时间)Fig.2 Diurnal variation of hydro-meteorological factors of Qiantang River in summer of 2020 (The dotted line represents the sampling time)
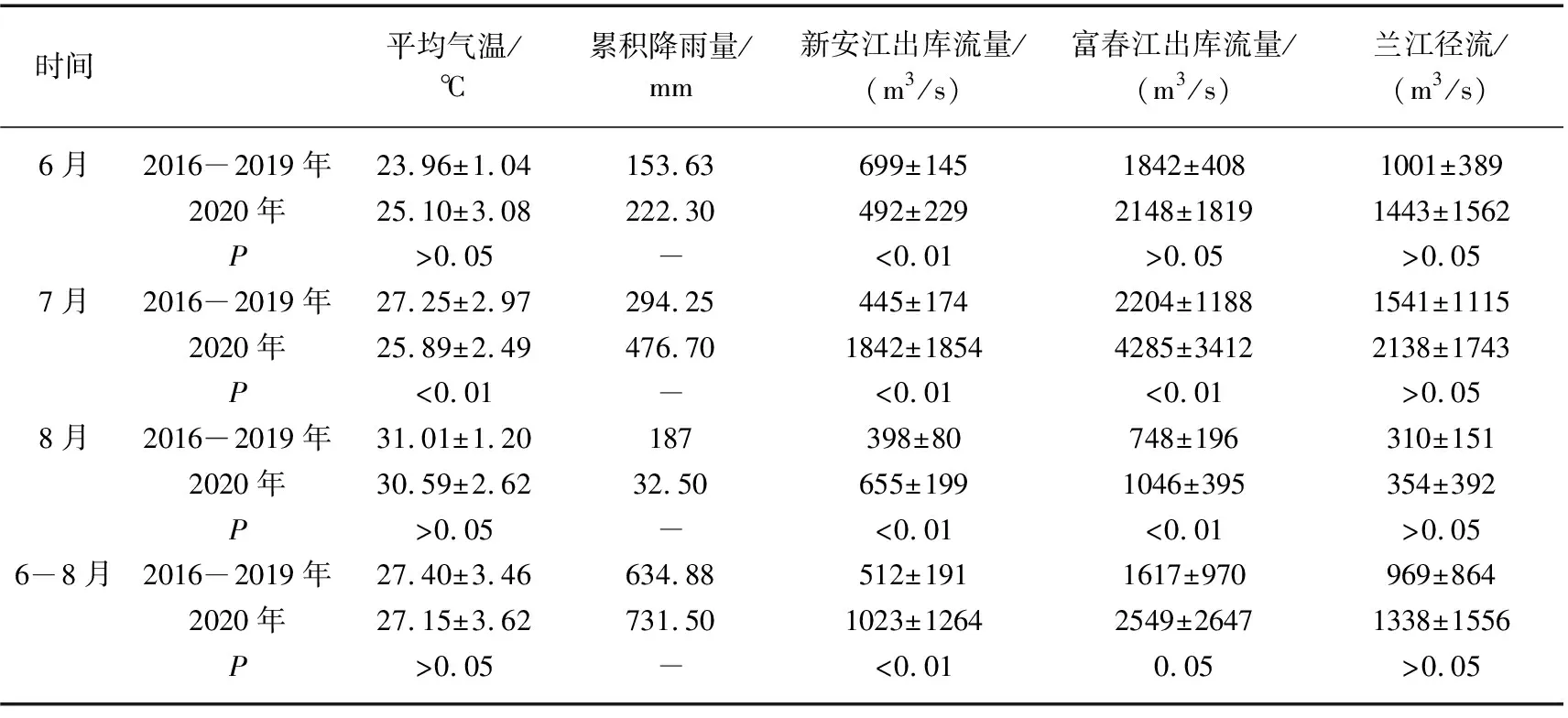
表1 2016-2019年与2020年钱塘江夏季主要水文气象因子比较Tab.1 Differences in hydro-meteorological factors of Qiantang River in summer between 2016-2019 and 2020
2.2 水体理化指标变化特征
钱塘江干流的水体理化特征的时间变化如图3所示. 研究期间,除了CODMn外,碳氮磷浓度在月间无显著性差异(P>0.05),DOC、TN、TP的平均浓度分别为2.28、1.64和0.08 mg/L. CODMn在月间差异显著(P<0.05),其中8月为2.37 mg/L,显著高于6月(1.47 mg/L)和7月(1.67 mg/L). 空间变化如图4所示,富春江库区的各水体理化指标均与上下游江段具有明显差异:库区上游的新安江段(S5)水质较好,而兰江及以上江段(S1~S4)水质较差,尤其在金华江段(S2),营养盐浓度明显高于其他断面. DOC浓度变化范围为1.10~4.34 mg/L,最大值在金华江段(S2),最小值在新安江段(S5). TN浓度变化范围为1.10~4.34 mg/L,最大值在金华江段(S2),最小值在富春江库区(S6);TP浓度在0.022~0.208 mg/L之间变化,最大值在下游钱塘江河口区(S11),最小值在富春江库区(S6). CODMn在0.85~4.43 mg/L之间变化,最大值和最小值分别出现在兰江的将军岩(S4)和新安江段(S5).
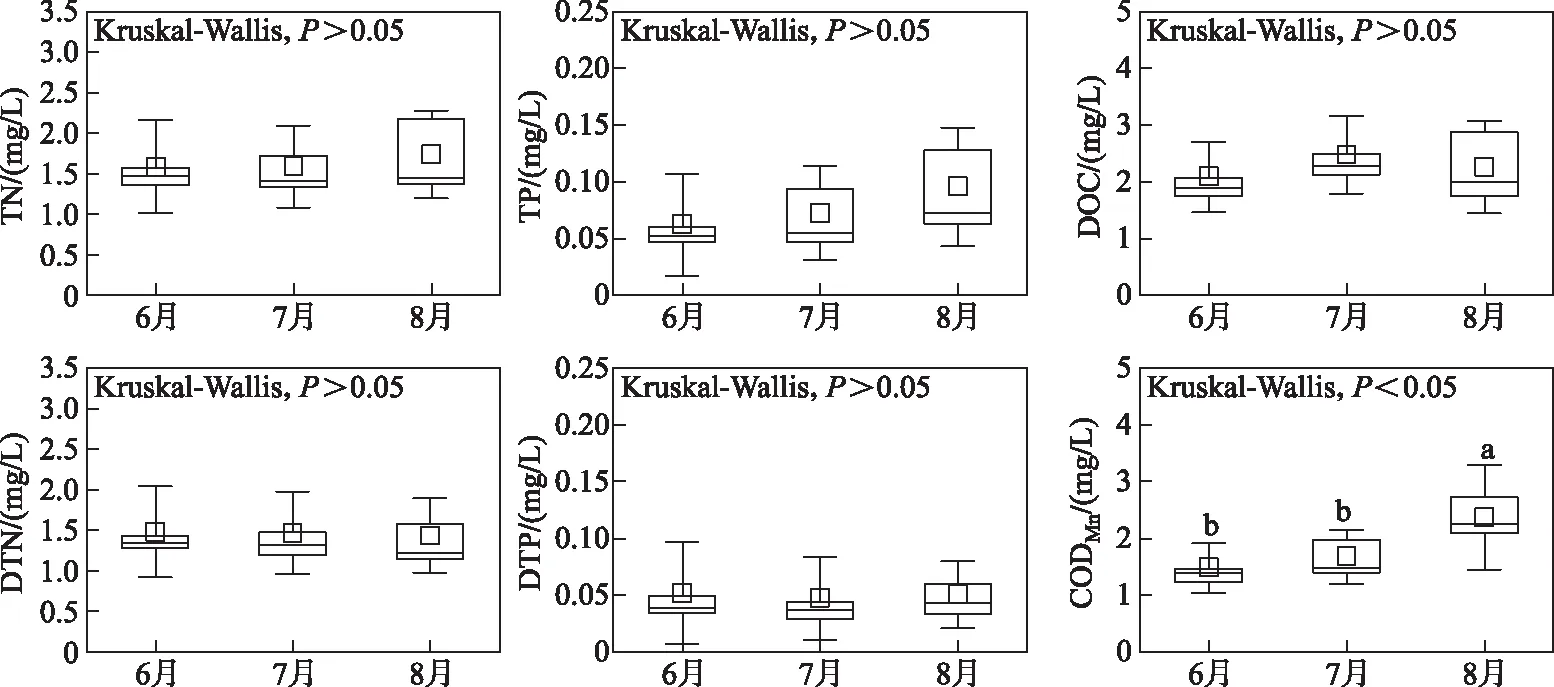
图3 2020年夏季钱塘江水体理化指标浓度时间变化Fig.3 Temporal variation of water quality index concentration at the Qiantang River in summer of 2020
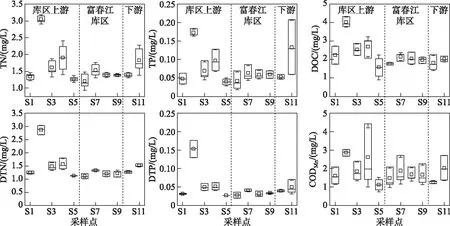
图4 2020年夏季钱塘江水体理化指标浓度空间变化Fig.4 Spatial variation of water quality index concentration at the Qiantang River in summer of 2020
2.3 浮游植物群落结构组成和优势属变化
2.3.1 浮游植物群落组成及细胞密度 2020年夏季钱塘江干流共鉴定出浮游植物95种,隶属于6门59属,包括蓝藻门8属9种、硅藻门21属38种、绿藻门24属41种、隐藻门1属2种、甲藻门2属2种和裸藻门3属3种. 钱塘江干流浮游植物细胞密度时空变化如图5所示,浮游植物细胞密度月间差异显著(P<0.01),8月细胞密度显著高于6、7月(图5a). 浮游植物夏季平均细胞密度为(1.64±3.07)×106cells/L,6-8月平均细胞密度分别为(0.19±0.14)×106、(0.54±0.42)×106和(4.06±4.38)×106cells/L. 空间上浮游植物细胞密度在富春江库区高于库区上下游,最高值在富春库区的三都大桥(S7),为1.31×107cells/L,最低值在新安江段(S5),为3.6×104cells/L(图5b).
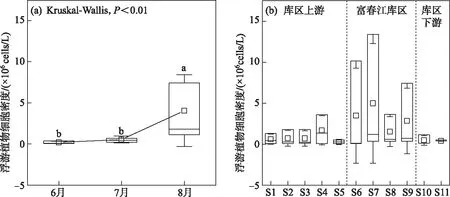
图5 2020年夏季钱塘江浮游植物细胞密度时空变化Fig.5 Spatiotemporal variation of phytoplankton cell densities at Qiantang River in summer of 2020
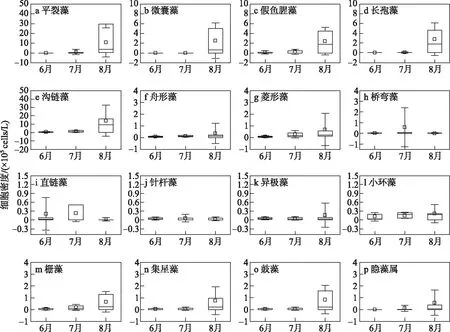
图6 2020年夏季钱塘江浮游植物优势属细胞密度时间变化趋势(因矛丝藻仅在S8出现, 辐环藻只在S11出现,故在图中未展现. a~d为蓝藻门;e~l为硅藻门;m~o为绿藻门;p为隐藻门)Fig.6 Temporal change of cell densities of dominant genus of phytoplankton in Qiantang River in summer of 2020
2.3.2 浮游植物优势属组成及丰度的变化特征 夏季各月份之间优势属组成和丰度差异较大,如图6所示. 整个夏季沟链藻(Aulacoseira)占据绝对优势,梅雨期6和7月主要为硅藻门的沟链藻、直链藻(Melosira)、小环藻(Cyclotella)、菱形藻(Nitzschia)、舟形藻(Navicula)、桥弯藻(Cymbella)、异极藻(Gomphonema)和针杆藻(Synedra)占优,其次为绿藻门的栅藻(Scenedesmus),蓝藻门的平裂藻(Merismopedia)和假鱼腥藻(Pseudoanabaena). 梅雨后主要优势属为蓝藻门的平裂藻、微囊藻(Microcystis)、长孢藻(Dolichospermum)和假鱼腥藻,以及硅藻门的沟链藻.
各点位之间优势属组成和丰度差异较大,如表2所示. 富春江库区(S6~S8)出现的优势属种类最多,细胞密度最高,其次为钱塘江河口区闻家堰(S11)和兰江及以上(S1~S4)的各监测断面,种类最少、细胞密度最低的为新安江(S5). 从兰江到富春江库区,浮游植物优势属从硅藻占优逐渐演替为蓝藻占优;富春江库区下游则又变为硅藻占优. 其中衢江(S1)主要优势属为沟链藻、平裂藻和菱形藻,平均细胞密度分别为3.66×105、1.12×105、0.09×105cells/L;金华江(S2)主要优势属为菱形藻、舟形藻和假鱼腥藻,平均细胞密度分别为1.68×105、1.12×105、1.08×105cells/L;在金华江和衢江的汇合断面兰江(S3)主要优势属为沟链藻、假鱼腥藻和栅藻,平均细胞密度分别为3.39×105、1.50×105、0.42×105cells/L. 新安江(S5)主要为硅藻门的沟链藻、直链藻和针杆藻,平均细胞密度分别为1.08×105、0.23×105、0.24×105cells/L. 在富春江库区(S6~S8),优势属为平裂藻、长孢藻、微囊藻和沟链藻,细胞密度最高值分别为3.94×106、1.02×106、1.03×106、5.38×106cells/L;下游坝下(S10)主要为沟链藻和平裂藻,平均细胞密度分别为1.75×105和1.28 ×105cells/L;闻家堰(S11)主要优势属为辐环藻(Actinocyclus)和沟链藻,平均细胞密度分别为1.37×105和0.84×105cells/L.
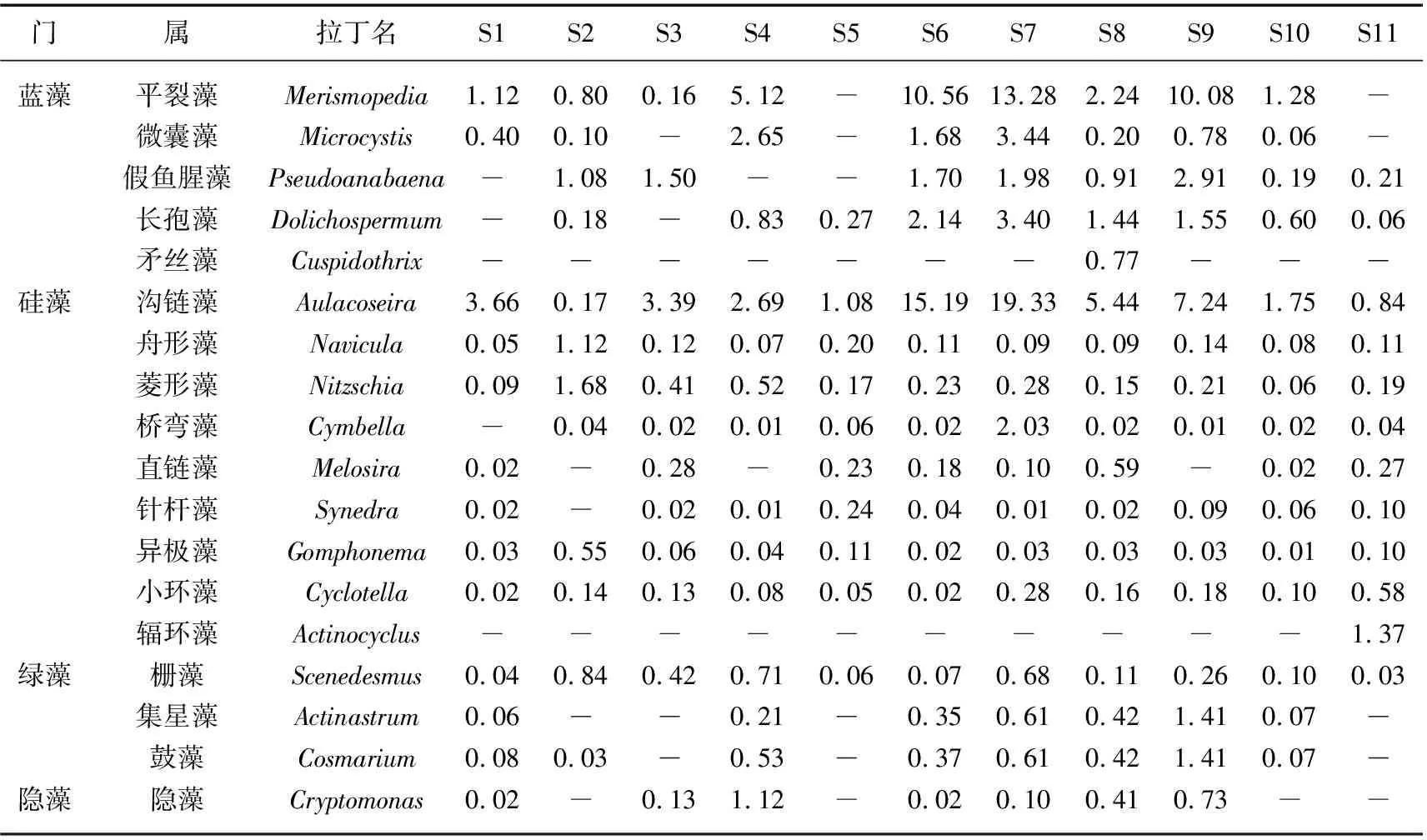
表2 2020年夏季钱塘江各监测河段浮游植物优势属平均细胞密度(×105 cells/L)Tab.2 Spatial change of cell densities of dominant genus of phytoplankton in Qiantang River in summer of 2020(×105 cells/L)
2.4 浮游植物与环境因子之间的关系
浮游植物与环境因子的关系如图7所示,平均气温(AT)、DTP、TP、DOC、CODMn、OSS和累积降雨量(Rainfall)与钱塘江干流浮游植物群落结构呈相关(P< 0.05),7个环境因子共解释了72.73%的浮游植物群落结构变化. 其中平均气温解释率为30.5%(P=0.002),累积降雨量解释率为25.9%(P=0.002). 因此推断,气温与降雨量是驱动钱塘江干流河道浮游植物群落结构变化的最主要因子. 蓝藻门主要与气温呈正相关,与降雨量呈负相关,主要包括微囊藻、长孢藻、平裂藻和假鱼腥藻. 但浮丝藻与之相反,与气温呈负相关,与降雨呈正相关. 硅藻门与气温和降雨量的关系与蓝藻门相反,其中沟链藻受到气温和降雨量的影响较小. 绿藻门种类丰富,生态位较宽,与气温呈正相关,与降雨量呈负相关.
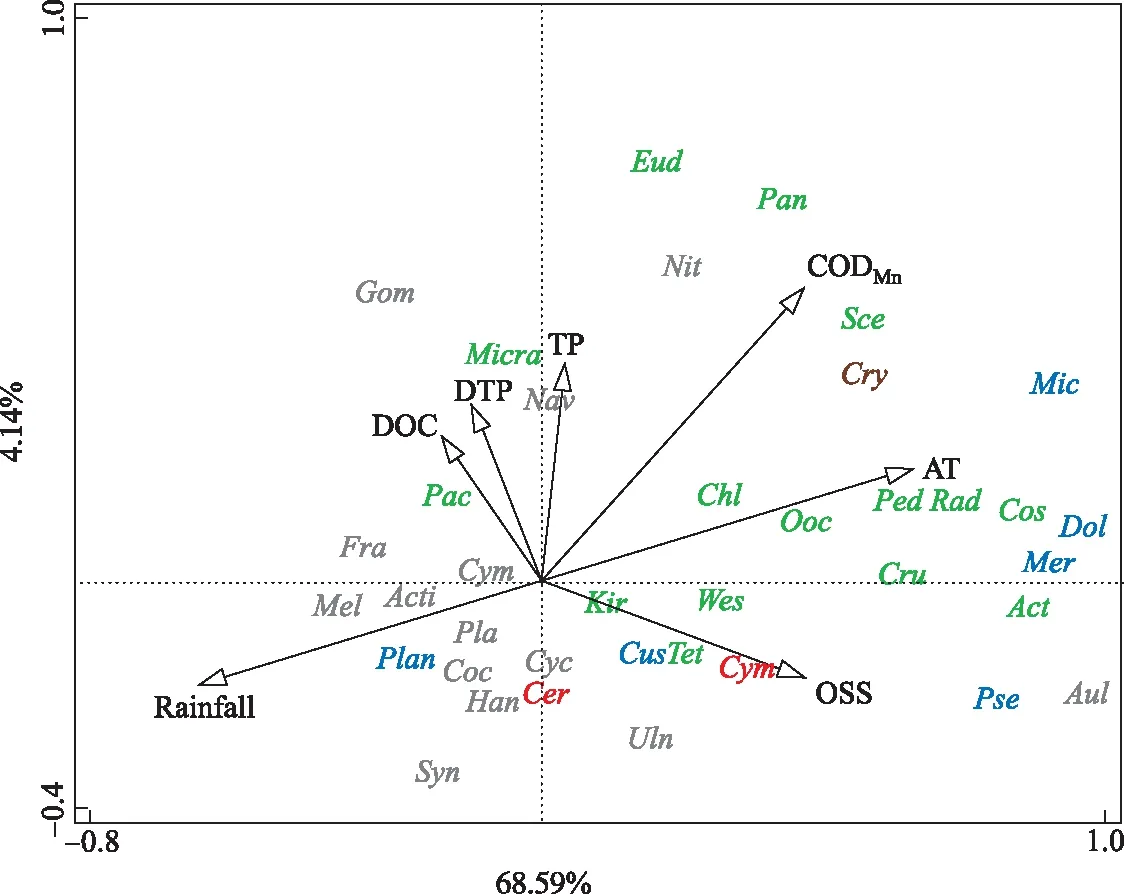
图7 钱塘江浮游植物与环境因子的RDA排序图(蓝藻门:微囊藻(Mic);平裂藻(Mer);矛丝藻(Cus); 假鱼腥藻(Pse);浮丝藻(Plan);长孢藻(Dol);绿藻门:栅藻(Sce);衣藻(Chl);韦斯藻(Wes); 微芒藻(Micra);蹄形藻(Kir);四角藻(Tet);实球藻(Pan);十字藻(Cru);盘星藻(Ped); 卵囊藻(Ooc);空球藻(Eud);集星藻(Act);厚枝藻(Pac);鼓藻(Cos);辐球藻(Rad); 硅藻门:肘形藻(Uln);舟形藻(Nav);直链藻(Mel);针杆藻(Syn);异极藻(Gom);小环藻(Cyc); 桥弯藻(Cym);平面藻(Pla);卵形藻(Coc);菱形藻(Nit);菱板藻(Han);沟链藻(Aul); 脆杆藻(Fra);辐环藻(Acti);隐藻门:隐藻(Cry);甲藻门:裸甲藻(Gym);角甲藻(Cer). 蓝色代表蓝藻门,绿色代表绿藻门,灰色代表硅藻门,褐色代表隐藻门,橙色代表甲藻门)Fig.7 RDA analysis of phytoplankton and environmental factors in Qiantang River
3 讨论
在调查期间,钱塘江的浮游植物群落结构由梅雨期的硅藻占优,演替为梅雨后的蓝藻占优,包括微囊藻、长孢藻等水华蓝藻常见属. 2020年梅雨期的累积降雨量比历史同期高,其中7月为历史同期的1.6倍(表1),并且在7月发生了4场暴雨. 暴雨不仅会对温度、光照、营养盐和流量等水体环境造成改变,对浮游植物的生长带来影响;而且会使水中的悬浮颗粒物增多,影响水体的浊度,抑制浮游植物的光合作用;还会增加水体掺杂作用,稀释浮游植物的细胞密度[33-34]. 因此降雨量的增加使得研究区域内水体流量变大,换水周期变短,水体冲刷速率变快[35]. 虽然高换水率有利于硅藻的生长和占优[36],但是大流量伴随的强扰动对光照条件产生影响,影响藻类在水中的生存状态,不利于藻类对营养盐的吸收. 暴雨影响了钱塘江干流浮游植物群落结构的演替. 所以在整个梅雨期硅藻占优的情况之下,浮游植物的细胞密度仍然较低[37]. 而2020年,梅雨后降雨骤减,累积降雨量仅为同期17.38%,使得光照条件显著改善,对藻类生长具有明显的促进效应[38]. 此外,梅雨后富春江库区换水周期明显变长,由梅雨期的1.32 d变为5.77 d,在这一条件下,水体扰动变小,使得微囊藻、长孢藻等水华蓝藻能够快速的生长繁殖,进而占据优势,具备了形成蓝藻水华的条件.
来水量增加会减少水力滞留时间,减缓蓝藻水华,这时可适当通过加大水体冲刷作用的方式,降低蓝藻水华暴发风险[39-40],此次调查中富春江水库的平均出库流量为2549 m3/s,调查期间也未见蓝藻水华,这一研究结果相符于此前吴挺峰等[23]在研究中表明的当水体下泄流量达到600 m3/s,就能防止富春江水库的水华暴发.
流态变化是钱塘江干流浮游植物群落结构时空差异的重要影响因子,回水区、水流滞缓区是蓝藻水华形成的易发区. 富春江水库浮游植物细胞密度最高,其次是金华江段和下游钱塘江河口. 河流筑坝建库发电会影响水动力条件,根据文献报道,兰江口水体流速能达到梅城段的5倍左右,自兰江口到三都境内的流速都低于兰江,在富春江库区河流渐宽,流速渐小[41]. 新安江来水的梅城大桥(S6)到三都大桥(S7)段和兰江来水的将军岩(S4)到三都大桥(S7)段是富春江水库的主要回水区. 新安江电站和富春江电站是日调节型电站,在夏季白天用电高峰发电,晚上停发. 这种调度模式之下,富春江水库形成了时断时流的间歇性水流特征,当停止发电时,富春江水库库区是底层低温水流向兰江上溯,富春江水库和兰江水向新安江回流,从而导致水体滞留,相关的研究结果也表明富春江存在一定时间内的往复运动和静止状体[24]. 与此同时,富春江水库水深相对较深(图8),水体开阔,增加了水流滞缓的程度,再加上水体温度分层的作用,致使浮游植物细胞密度在此江段最高. 而筑坝减缓水体的流速在文献中已有报道[42],在金华江段的采样点上是一个水坝,也导致该江段水流相对滞缓,其浮游植物细胞密度也较衢江和兰江较高;而钱塘江口则属于咸淡水交汇区,浮游植物细胞密度相对富春江库区较低. 此次调查的金华江、将军岩、钱塘江口都是处于水体汇合区域,可能存在回力顶托的作用,使得水体流速减缓,进而有助于浮游植物相对较高.
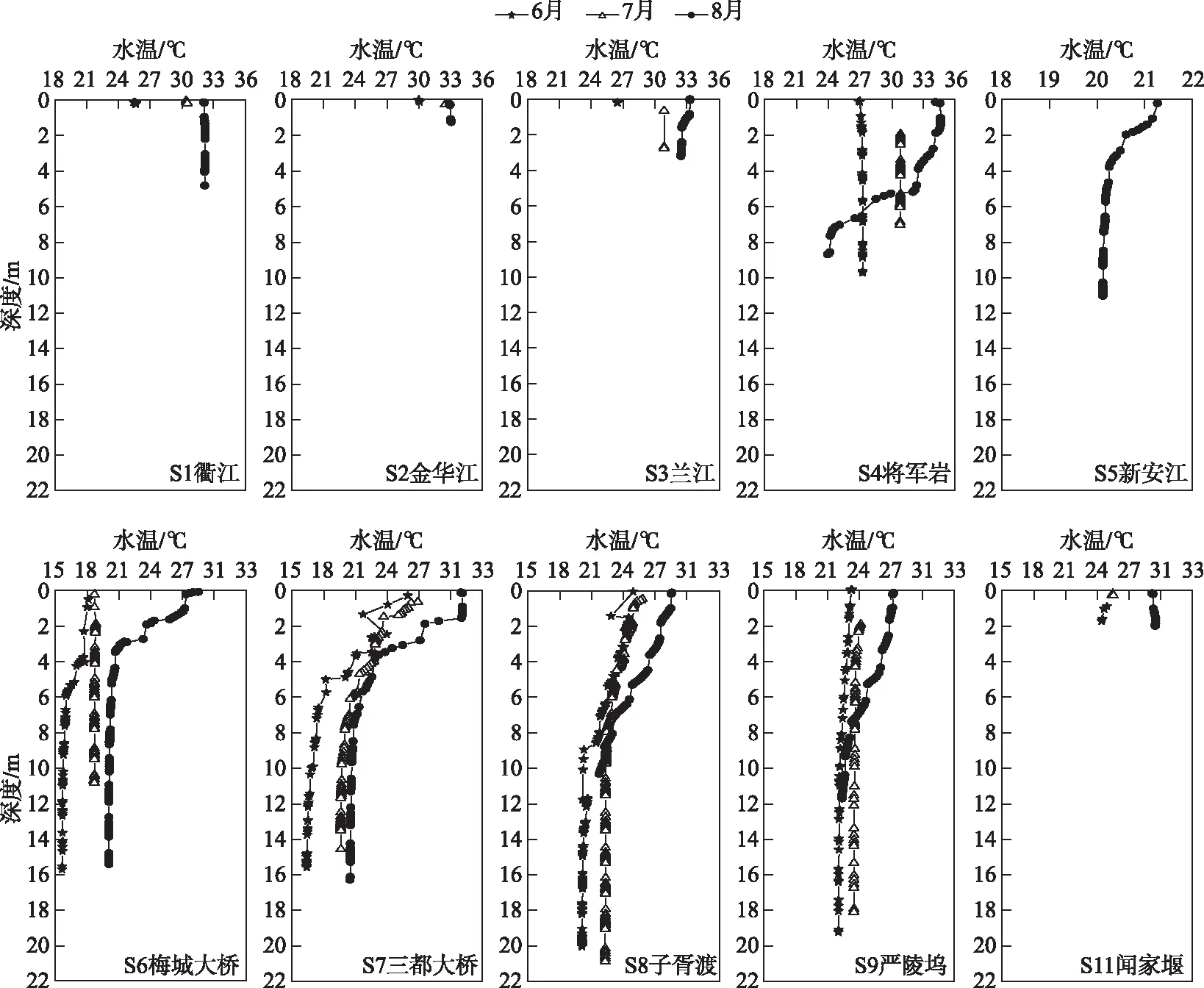
图8 钱塘江关键断面水温垂直剖面Fig.8 Stratification of environmental water temperature in summer obtained at Qiantang River
此前许多研究报道了在水库的回水区均暴发过蓝藻水华. 例如三峡水库的小江回水区、香溪河回水区、澎溪河回水区等[8,43-45]. 富春江水库作为典型的亚热带大型河流型水库,因为筑坝建库使得库区的水体流速相对滞缓,相对利于藻类的生长繁殖,多个优势属的细胞密度均在富春江库区出现峰值,如微囊藻、长孢藻、平裂藻、沟链藻等. 相比于库区而言,衢江段、金华江段和兰江段(S3)水较浅(图8),水团混合较为充分,换水速率较高,所以夏季浮游植物细胞密度低于库区. 而新安江段(S5)则主要是因为水温低,水质良好,营养盐浓度较低,其浮游植物生长受到限制. 因此,应当重点关注富春江水库回水区和水流滞缓区的浮游植物动态.
持续的高温晴热天气为蓝藻异常增殖提供了契机. 梅雨后的高温晴热,导致富春江库区出现明显的温度分层(图8). 同时库区蓝藻细胞密度快速增高,微囊藻的最高密度达到了1.03×106cells/L. 梅雨后,雨量骤降,高温晴热天气的接踵而至,加之新安江电站下泄水温较低,兰江来水温度较高,更加利于将军岩和富春江库区出现明显的温度分层[24]. 温度分层会阻碍水体垂向的水动力交换,使得水体中溶解氧、营养盐等指标的分层[35]. 当水体表层温度较高,并且混合减弱时,不利于个体小,沉降速率高的藻类细胞密度增加,而利于蓝藻尤其是具有气囊的微囊藻、长孢藻占优[15,46]. 微囊藻和长孢藻能够利用气囊上浮,在稳定的分层条件下,进入表层水体从而获得最优的光热条件[47],甚至在水体表面形成致密的水华,遮盖其他更深的非浮性真核浮游植物,从而增加其竞争力[48].
在分层条件下还可能会加剧底泥表面缺氧而使氧化还原点位发生改变,促进磷的内源释放,从而进一步提高水华蓝藻的细胞密度[49]. 以往的研究表明,在升温的条件下,2016年因为持续高温晴热天气在钱塘江干流发生了蓝藻水华[22],在白洋淀也因为温度的升高而导致浮游植物向蓝藻为主转变[50]. 温度上升能够促进藻类的生长,但是不同藻种具有不同的温度适应性,硅藻在16~26℃时生长较快,而蓝藻喜高温,在25~30℃具有最佳生长速度[51-52]. 总之,晴热天气的持续使得水体温度分层更加稳定,分层增强则更加有利于蓝藻占优势地位.
营养盐是浮游植物生长的重要因子,但其并不是钱塘江干流夏季浮游植物群落结构变化的主要控制因子. 氮浓度与浮游植物的群落结构并不显著相关(图7),6-8月TP平均浓度分别为0.06、0.07和0.1 mg/L,仅6月梅城大桥(S6)TP浓度为0.022 mg/L, 其余监测断面的TP浓度在0.032~0.208 mg/L之间波动,TP达到0.03 mg/L时,满足藻类的生长[53],并且已经具有较大水华蓝藻占优的可能[54]. 根据文献[55-56]计算出钱塘江干流的TSI(Chl.a)、TSI(TP)和TSI(TN)分别为46.24、64.76和160.67. 根据营养状态指数来看,当TSI(TN)或TSI(TP)大于TSI(Chl.a)时可认为氮或磷不是浮游植物生长最主要的限制因子[57-58]. 由此来看,在钱塘江干流营养盐所能支持的藻类最大生长潜力并未达到.
综合而言,2020年夏季钱塘江干流的浮游植物群落结构主要受到水文气象过程的驱动,同时,流态变化是重要的影响因子,而受营养盐影响较小. 梅雨期,雨量增多,水体流量较大,水体扰动强而不利于浮游植物的生长和增殖;梅雨后,雨量变少,水体流量骤减,换水周期变长,为浮游植物的生长提供了极其有利的环境条件. 此外,持续高温晴热天气导致的温度分层,为蓝藻的异常增殖提供了条件. 钱塘江干流已经发生过大规模的蓝藻水华,并且仍然具有较高的蓝藻水华暴发风险. 夏季梅雨期后的持续高温晴热天气是暴发蓝藻水华的高危期,应当着重关注,加以防范,并且应当重点关注其回水区和水流滞缓区的浮游植物动态变化.
4 结论
1) 2020年夏季钱塘江干流共鉴定出浮游植物95属(种),主要由硅藻门、蓝藻门和绿藻门组成,主要优势属为沟链藻、菱形藻、平裂藻、长孢藻、假鱼腥藻和微囊藻属. 夏季平均藻细胞密度为(1.64±3.07)×106cells/L,其中梅雨期藻细胞密度为(0.37±0.36)×106cells/L,显著低于梅雨后(4.06±4.38)×106cells/L.
2) 梅雨影响了钱塘江干流浮游植物群落的演替,浮游植物群落组成从梅雨期的硅藻占优,演替为梅雨后的蓝藻占优,且梅雨后期持续的高温天气进一步为微囊藻、长孢藻等水华蓝藻的异常增殖提供契机.
3) 流态的变化影响了钱塘江干流浮游植物群落结构的空间差异,富春江库区藻密度最高,其次为兰江及以上江段,新安江段藻密度最低,因此,梅雨后期应当重点关注库区和水流滞缓区的藻类动态,防控蓝藻水华暴发.
致谢:感谢杭州市生态环境科学研究院在数据收集中提供的帮助,感谢杭州市桐庐县环境保护监测站在采样工作的大力协助,感谢江南大学王子聪和西华师范大学郑清在采样过程中的帮助,感谢胡春华副研究员和刘莉实验员在样品测定中的帮助,感谢中国科学院南京地理与湖泊研究所的邹伟、史鹏程等在数据分析、绘图及论文修改中提供的帮助.
