生物节律蛋白、乙醇脱氢酶4、下丘脑泌素受体2基因多态性与丛集性头痛关联性的Meta分析
2022-03-04李涛卢改会赵敏
李涛,卢改会,赵敏
丛集性头痛(CH)又称“自杀性头痛”,是目前已知疼痛程度最为剧烈的原发性头痛。其主要临床特征是周期性丛集样发作的单侧眶周疼痛,伴随典型的三叉神经自主症状[1]。CH的患病率为101~151/100 000,男女比例约为4∶3[2]。虽然CH的患病率较低,但是治疗难度大,医疗费用高昂,往往给患者自身和社会带来沉重的负担[3]。大量关于CH的基础和临床研究被开展,但是确切病因和病理生理机制并不明确。近年来,随着基因测序技术和基因-疾病关联研究的发展,研究人员越来越关注遗传因素在CH发病过程中的作用[4]。截至目前,多项关于生物节律蛋白(CLOCK)、乙醇脱氢酶4(ADH4)、下丘脑泌素受体2(HCRTR2)基因多态性和CH易感性的研究在不同种族人群中被开展,但是结果并不一致。考虑到单一研究的样本量有限,把握度较低,且人群代表性较差,往往不能反映候选基因和疾病之间的真实关联。因此,本研究采用Meta分析的方法,对现有研究结果进行合并分析,以期得到更加真实可信的结果。
1 资料与方法
1.1 检索策略 系统检索PubMed、Cochrane Library、Web of Science、EMBASE、中国知网和万方数据库,末次检索时间为2020年6月20日。PubMed数据库的检索式:“Hypocretin receptor 2 OR HCRTR2 OR CLOCK OR Circadian Locomotor Output Cycles Kaput OR ADH4 OR Alcohol dehydrogenase Ⅳ OR Alcohol dehydrogenase 4” and “Polymorphism OR SNP OR Mutation OR mutant OR variant” and “Cluster headache”。中文数据库的检索式:“丛集性头痛”和“多态性”。此过程中如有分歧,向第三位作者咨询并通过共同商讨来解决分歧。
1.2 纳入和排除标准 纳入标准:(1)病例-对照研究或队列研究;(2)病例组符合CH的诊断标准;(3)评价ADH4、CLOCK、HCRTR2基因多态性和CH易感性的关联;(4)能获得全文,有病例组和对照组的基因型频率,能够计算OR和95%CI;(5)经过同行评议;(6)数据有重叠的文献,纳入最近一次发表的研究。排除标准:会议摘要、综述、读者来信、述评、基础研究、病例报道和学位论文。此过程中如有分歧,向第三位作者咨询并通过共同商讨来解决分歧。
1.3 文献筛选和资料提取 两名研究人员分别按照检索策略进行文献检索,然后通过阅读题目、摘要和全文,并按照纳入和排除标准进行文献的筛选。确定文献符合要求后,提取以下资料:第一作者姓名、国家、人种、研究对象的来源、诊断标准和基因型频率分布。此过程中如有分歧,向第三位作者咨询并通过共同商讨来解决分歧。
1.4 方法学质量评价 两位研究人员分别根据纽卡斯尔-渥太华量表(NOS)评估纳入研究的方法学质量。NOS从人群选择、组间可比性及暴露因素3个方面对纳入研究进行评价,共包括8个条目,具体见http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp。当纳入研究得分不少于6分时,即为高质量研究。此过程中如有分歧,向第三位作者咨询并通过共同商讨来解决分歧。
1.5 统计学分析和发表偏倚评估 计算纳入文献中的对照组人群基因频率是否符合哈代-温伯格遗传平衡定律(HWE)(http://ihg.gsf.de/cgi-bin/hw/hwa1.pl)[5]。利用RevMan 5.3软件进行统计学分析。分别用1和2代表等位基因1和等位基因2,计算出各多态性位点在等位基因模型(1vs.2)、纯合子模型(11vs.22)、杂合子模型(12vs.22)、显性模型(11+12vs.22)及隐性模型(11vs.12+22)的OR及95%CI。用I2检验来评估纳入研究之间的异质性:当P≥0.1,I2<50%时,表明异质性较小,采用固定效应模型(FEM);反之,则采用随机效应模型(REM)[6]。
2 结 果
2.1 文献筛选结果 通过对电子数据库的系统检索,共得到96篇相关文献。去除重复和不符合要求的文献后,最终纳入了13篇[7-19],文献筛选过程见流程图(图1)。
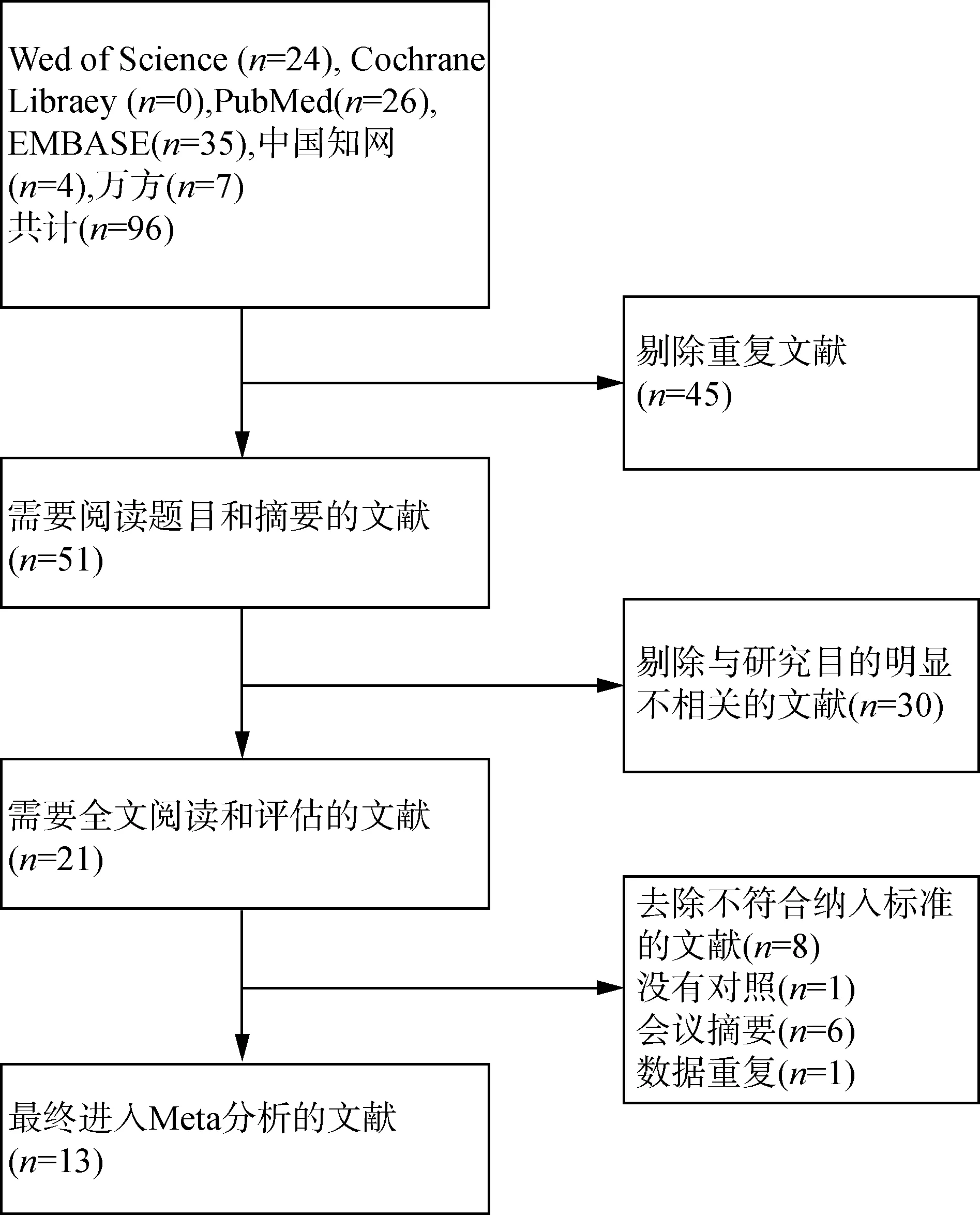
图1 文献筛选的流程图
2.2 纳入文献的基本特征 本研究共纳入13篇文献,包括15项独立研究,基本特征见表1。其中Baumber等[18]的研究包括三项独立研究,分别在丹麦、瑞典和英国三个不同人群中开展。被2篇以上文献报道的位点共有6个,分别是CLOCK基因的rs1801260位点、ADH4基因的rs1800759和rs1126671位点,以及HCRTR2基因的rs2653349、rs3122156和rs2653342位点。除了Fan等[10]的研究是在亚洲人群中开展外,其余各项研究均在高加索人中实施。3项研究[13,17-18]为队列研究,其余10项研究[7-12,14-16,19]为病例-对照研究。分别有5项[8-12]、1项[7]、7项[13-19]研究使用ICHD-Ⅲ beta[20]、ICHD-Ⅲ[21]和ICHD-Ⅱ[22]作为CH的诊断标准。需要注意的是Zarrilli等[12]关于rs1800759、rs1126671、rs2653349位点的研究,Fourier等[8]关于rs3122156位点的研究及Rainero等[15]关于rs2653342位点的研究中对照组不符合HWE。根据NOS评价标准,纳入的每项研究至少得到了6分,均为高质量研究(表2)。
2.3 Meta分析
2.3.1CLOCK基因多态性和CH的关系 见表3。5项研究[9-10,12,16,19]报道了rs1801260多态性和CH风险的关系,共纳入了823例CH患者和1 367名对照者。五种遗传模型下的异质性均较小,采用FEM进行统计分析。Meta分析结果显示,在等位基因模型下,rs1801260多态性和CH易感性(Cvs.T)无相关性(OR=1.10,95%CI:0.95~1.28,P=0.19)(图2)。在其余遗传模型下,rs1801260多态性和CH风险也无相关性(表3)。
2.3.2ADH4基因多态性和CH的关系 见表3。5项研究[7,10-12,14]报道了rs1800759多态性和CH风险的关系,共纳入了780例CH患者和1 548名对照者。
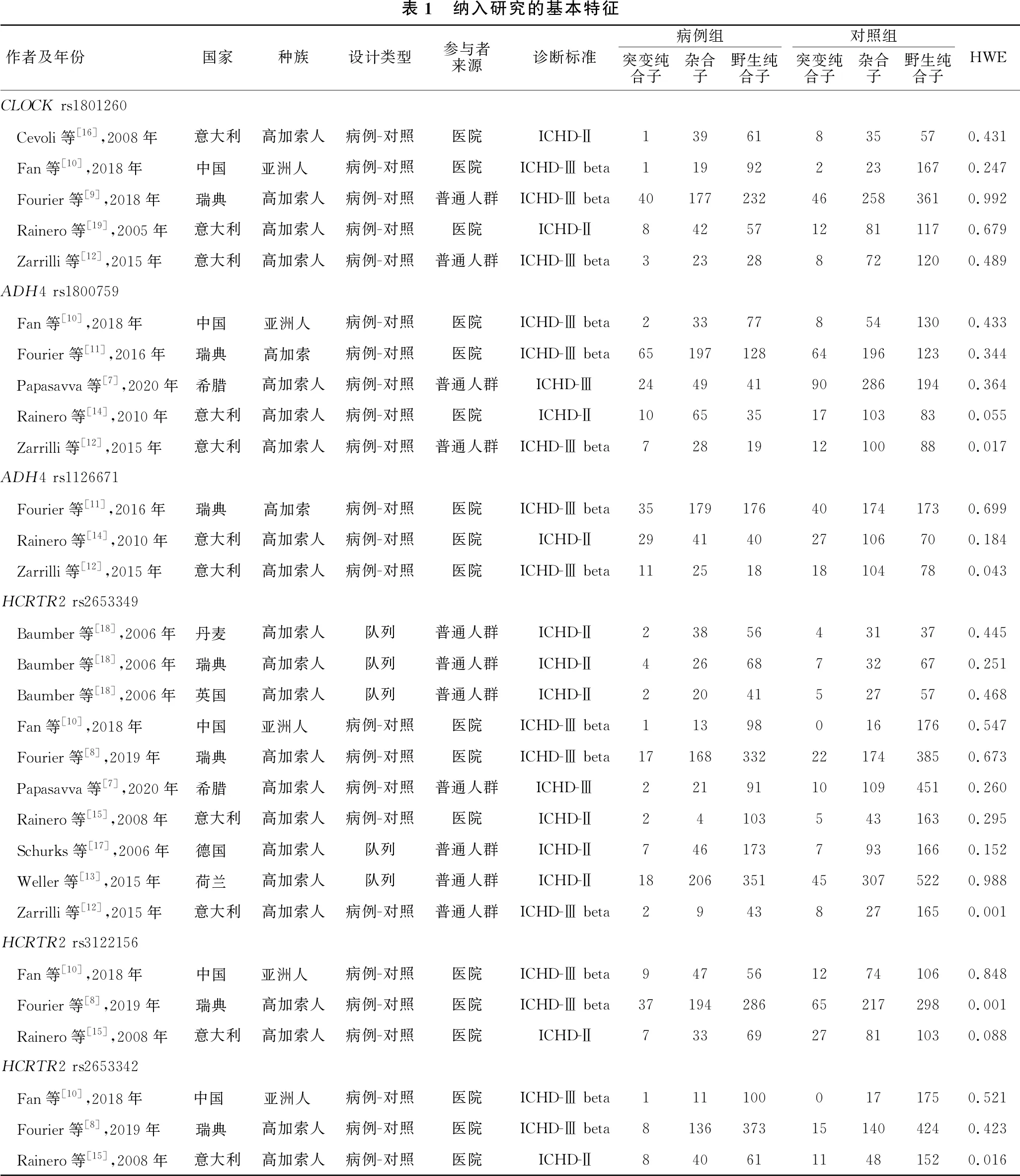
表1 纳入研究的基本特征作者及年份国家种族设计类型参与者来源诊断标准病例组突变纯合子杂合子野生纯合子对照组突变纯合子杂合子野生纯合子HWECLOCK rs1801260 Cevoli等[16],2008年意大利高加索人病例-对照医院ICHD-Ⅱ13961835570.431 Fan等[10],2018年中国 亚洲人 病例-对照医院ICHD-Ⅲ beta119922231670.247 Fourier等[9],2018年瑞典 高加索人病例-对照普通人群ICHD-Ⅲ beta40177232462583610.992 Rainero等[19],2005年意大利高加索人病例-对照医院ICHD-Ⅱ8425712811170.679 Zarrilli等[12],2015年意大利高加索人病例-对照普通人群ICHD-Ⅲ beta323288721200.489ADH4 rs1800759 Fan等[10],2018年中国 亚洲人 病例-对照医院ICHD-Ⅲ beta233778541300.433 Fourier等[11],2016年瑞典 高加索 病例-对照医院ICHD-Ⅲ beta65197128641961230.344 Papasavva等[7],2020年希腊 高加索人病例-对照普通人群ICHD-Ⅲ244941902861940.364 Rainero等[14],2010年意大利高加索人病例-对照医院ICHD-Ⅱ10653517103830.055 Zarrilli等[12],2015年意大利高加索人病例-对照普通人群ICHD-Ⅲ beta7281912100880.017ADH4 rs1126671 Fourier等[11],2016年瑞典 高加索 病例-对照医院ICHD-Ⅲ beta35179176401741730.699 Rainero等[14],2010年意大利高加索人病例-对照医院ICHD-Ⅱ29414027106700.184 Zarrilli等[12],2015年意大利高加索人病例-对照医院ICHD-Ⅲ beta11251818104780.043HCRTR2 rs2653349 Baumber等[18],2006年丹麦 高加索人队列普通人群ICHD-Ⅱ23856431370.445 Baumber等[18],2006年瑞典 高加索人队列普通人群ICHD-Ⅱ42668732670.251 Baumber等[18],2006年英国 高加索人队列普通人群ICHD-Ⅱ22041527570.468 Fan等[10],2018年中国 亚洲人 病例-对照医院ICHD-Ⅲ beta113980161760.547 Fourier等[8],2019年瑞典 高加索人病例-对照医院ICHD-Ⅲ beta17168332221743850.673 Papasavva等[7],2020年希腊 高加索人病例-对照普通人群ICHD-Ⅲ22191101094510.260 Rainero等[15],2008年意大利高加索人病例-对照医院ICHD-Ⅱ241035431630.295 Schurks等[17],2006年德国 高加索人队列普通人群ICHD-Ⅱ7461737931660.152 Weller等[13],2015年荷兰 高加索人队列普通人群ICHD-Ⅱ18206351453075220.988 Zarrilli等[12],2015年意大利高加索人病例-对照普通人群ICHD-Ⅲ beta29438271650.001HCRTR2 rs3122156 Fan等[10],2018年中国 亚洲人 病例-对照医院ICHD-Ⅲ beta9475612741060.848 Fourier等[8],2019年瑞典 高加索人病例-对照医院ICHD-Ⅲ beta37194286652172980.001 Rainero等[15],2008年意大利高加索人病例-对照医院ICHD-Ⅱ7336927811030.088HCRTR2 rs2653342 Fan等[10],2018年中国 亚洲人 病例-对照医院ICHD-Ⅲ beta1111000171750.521 Fourier等[8],2019年瑞典 高加索人病例-对照医院ICHD-Ⅲ beta8136373151404240.423 Rainero等[15],2008年意大利高加索人病例-对照医院ICHD-Ⅱ8406111481520.016
五种遗传模型均未检测到显著异质性,采用FEM进行统计分析。Meta分析结果显示,在等位基因模型下,rs1800759多态性(Avs.C)和CH无关(OR=1.06,95%CI:0.93~1.22,P=0.37)(图3A)。在其余遗传模型下,也未发现rs1800759多态性和CH易感性有关。
3项研究[11-12,14]报道了rs1126671多态性和CH易感性的关联,纳入了554例CH患者和790名对照者。纯合子模型和隐性模型检测到了显著的异质性,采用REM进行统计分析,其余遗传模型采用FEM分析。结果显示,在等位基因模型下,rs1126671多态性(Avs.G)和CH无关(OR=1.09,95%CI:0.92~1.28,P=0.32)(图3B)。在其余遗传模型下,未检测到rs1126671多态性和CH有关(表3)。
2.3.3HCRTR2基因多态性和CH的关系 8项研究[7-8,10,12-13,15,17-18]报道了rs2653349多态性和CH风险的关系,共纳入1 964例CH患者和3 161名对照者。在等位基因模型、杂合子模型和显性模型下检测到了显著的异质性,采用REM进行统计分析,其余遗传模型采用FEM分析。Meta分析结果显示,在等位基因模型下,rs2653349多态性(Avs.G)和CH无关(OR=0.85,95%CI0.69~1.03,P=0.10)(图4)。在其余遗传模型下,未发现rs2653349多态性和CH存在关联。
3项研究[8,10,15]报道了rs3122156多态性和CH风险的关系,纳入了738例CH患者和983名对照者。在等位基因模型、纯合子模型和显性模型下检测到了显著的异质性,采用REM进行统计分析,其余遗传模型采用FEM分析。结果显示,在等位基因模型下,rs3122156多态性(Gvs.T)和CH无关(OR=0.83,95%CI:0.60~1.16,P=0.27)。在其余遗传模型下,未发现rs3122156多态性和CH的关联。
3项研究[8,10,15]报道了rs2653342多态性和CH风险的关系,纳入了738例CH患者和982名对照者。在等位基因模型、杂合子模型和显性模型下检测到了显著的异质性,采用REM进行统计分析,其余遗传模型采用FEM分析。结果显示,在等位基因模型下,rs2653342多态性(Avs.G)和CH无关(OR=1.28,95%CI:0.86~1.92,P=0.22)。在其余遗传模型下,未发现rs2653342和CH的关联。

表2 纳入研究根据NOS的方法学质量评分结果作者及年份病例的确定是否恰当病例的代表性对照的选择对照的确定组间可比性暴露因素的确定采用相同的方法确定病例和对照组的暴露因素无应答率合计Papasavva等[7],2020年111111118Fourier等[8],2019年101111117Fourier等[9],2018年111111118Fan等[10],2018年100121117Fourier等[11],2016年101111117Zarrilli等[12],2015年111111118Weller等[13],2015年111101117Rainero等[14],2010年111121119Rainero等[15],2008年111121119Cevoli等[16],2008年100111116Schürks等[17],2006年111111118Baumber等[18],2006年111111118Rainero等[19],2005年111121119

表3 CLOCK、ADH4、HCRTR2基因多态性和丛集性头痛易感性的Meta分析结果遗传模型统计学关联OR值95%CIP值异质性检验I2(%)P值统计模型CLOCK rs1801260 C vs. T1.10 0.95~1.280.1900.45FEM CC vs. TT1.17 0.80~1.700.41250.26FEM CT vs. TT1.120.93~1.350.2400.85FEM CC+CT vs. TT1.13 0.94~1.350.2000.75FEM CC vs. CT+TT1.13 0.79~1.630.50240.26FEMADH4 rs1800759 A vs. C1.06 0.93~1.220.3700.46FEM AA vs. CC1.13 0.84~1.520.42190.29FEM AC vs. CC1.040.85~1.270.6900.43FEM AA+AC vs. CC1.06 0.88~1.280.5400.44FEM AA vs. AC+CC1.14 0.87~1.490.35170.30FEM

续表
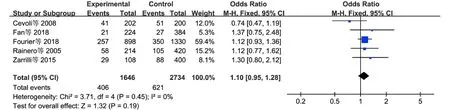
图2 等位基因模型下rs1801260多态性和CH风险的关系
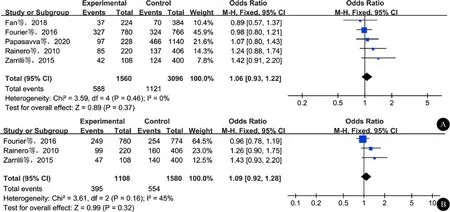
图3 等位基因模型下ADH4基因多态性和CH的关系 A:rs1800759多态性和CH风险的关联性;B:rs1126671多态性和CH风险的关联性
2.4 敏感性分析及发表偏倚评估 剔除不符合HWE的研究后,剩余研究总体效应值未发生显著改变。为了提高把握度,在最后的数据合并中没有将其排除。逐个剔除其余各项研究后,总体的效应值方向和大小均未发生明显改变,表明稳定性良好。漏斗图未见明显不对称,未检测到显著的发表偏倚(图5)。
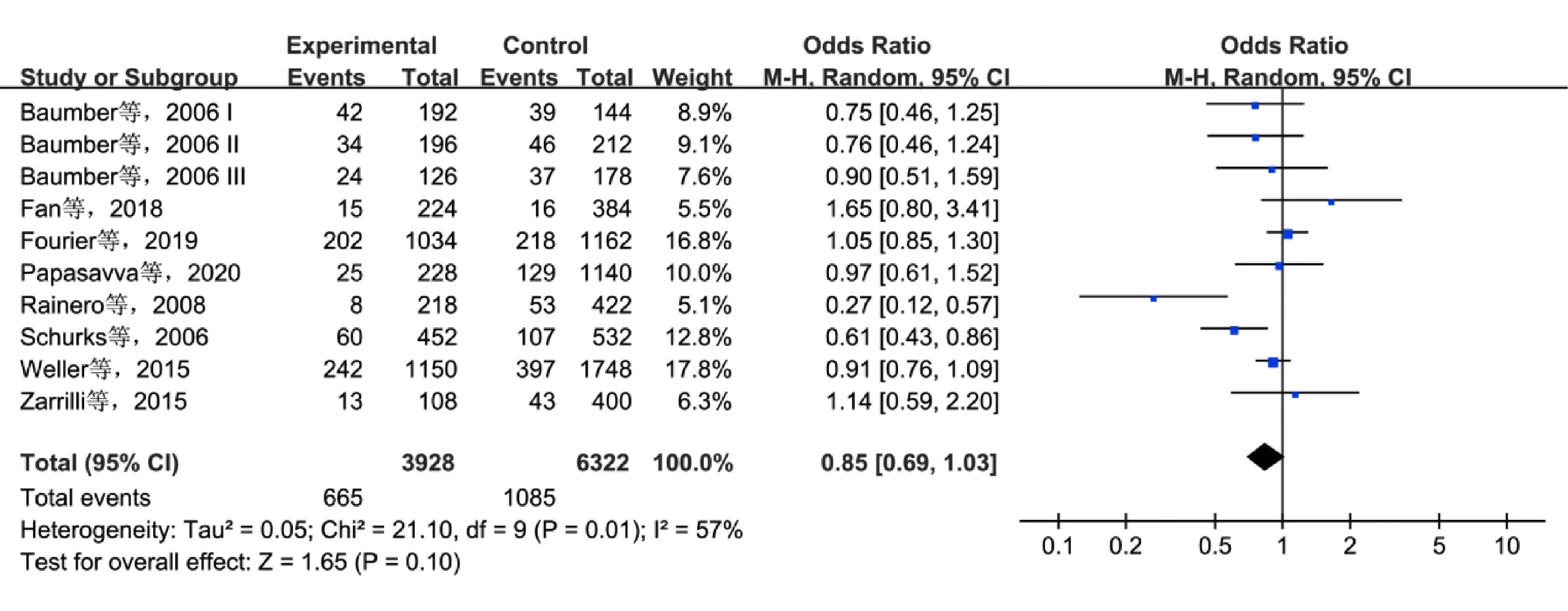
图4 在等位基因模型下rs2653349多态性和CH风险的关联性
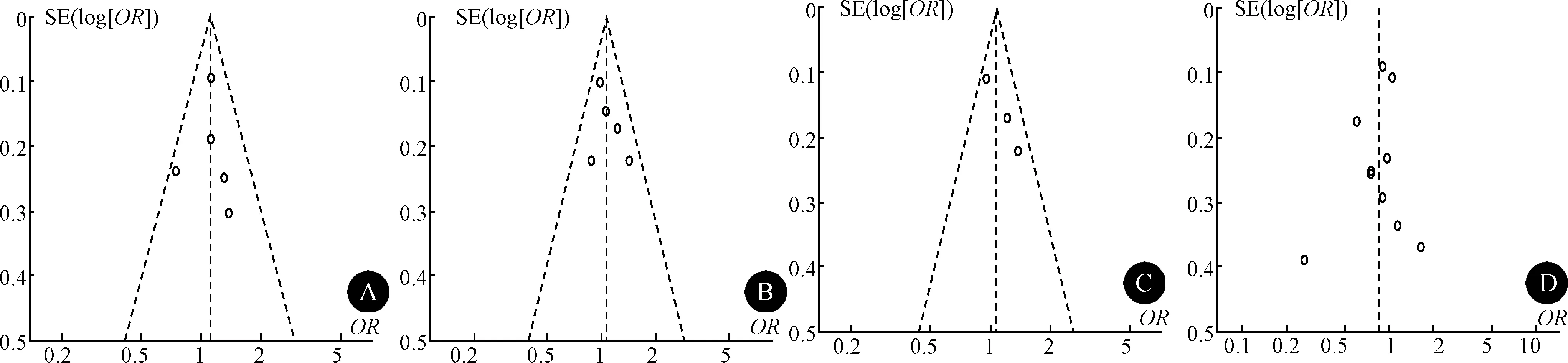
图5 发表偏倚评估结果 A:rs1801260多态性和CH关联性的漏斗图(C vs. T);B:rs1800759多态性和CH关联性的漏斗图(A vs. C);C:rs1126671多态性和CH关联性的漏斗图(A vs. G);D:rs2653349多态性和CH关联性的漏斗图(A vs. G)
3 讨 论
明确CH的病因和发病机制能够更好地指导其预防和临床治疗。然而,CH的确切病因仍不清楚。包括吸烟和饮酒在内的环境因素能够诱发CH的发作,但是环境因素只能部分解释CH的发生[23]。过去的十几年里,遗传因素在CH发生过程中的作用受到了越来越多的关注,且发现CH有明显的家族聚集现象。Leone等[24]实施的大样本临床研究显示,CH患者的一级亲属患CH风险是普通人群的39倍。随着基因关联研究的不断开展,CLOCK、HCRTR2、ADH4和CACNA1A等多个基因都被报道和CH有关[4]。关于CLOCK[9-10,12,16,19]、HCRTR2[7-8,10,12-13,15,17-18]、ADH4[7,10-12,14]基因多态性研究最多,但结果并不一致。这些差异可能是不同研究之间的临床异质性、遗传背景差异、诊断标准不一致、检测手段不同等造成的。Meta分析可以评估研究之间的异质性,并且能够通过合并数据,增加把握度和人群多样性,从而使研究结果更加真实可靠。本研究通过对rs1801260、rs1800759、rs1126671、rs2653349、rs3122156和rs2653342这6个最为广泛报道的多态性位点进行整合分析,以期通过扩大样本量来寻找其与CH风险的潜在遗传相关性。
CH的显著特征是发作过程具有明显的周期性和节律性[25]。CH常在夜间的某个固定时间点发作,由下丘脑调节的生物钟功能异常可能与CH病理生理过程有关。May等[26]发现,CH发作期间存在下丘脑的异常活动。CLOCK基因位于染色体4q12,编码调控生物钟的转录因子CLOCK。CLOCK能够和其他生物钟基因共同构成正、负反馈循环回路,使内源性的生物节律表现为接近24 h的循环周期。此外,CLOCK还能够通过神经-体液调节的方式调控哺乳动物的行为活动,从而使之更好地适应外部环境[27-28]。基础研究[29]表明,CLOCK还能够调控正常的细胞周期和细胞增殖。Fourier等[9]发现,CLOCK基因突变会导致其蛋白表达水平的显著增高,提示CLOCK基因可能参与CH的发生。Ofte等[27]认为,CLOCK参与CH发生的机制是CLOCK基因的突变导致其编码蛋白的活性改变,从而引起机体内部的生物钟紊乱,导致CH发生。
饮酒是CH的危险因素,酒精能够通过和多种神经递质的相互作用,打乱神经递质激活和抑制状态之间的平衡,干扰大脑的正常功能。文献[23]报道,53.5%~79.0%的CH患者会因饮酒诱发CH。Lund等[30]的研究结果表明,CH患者的饮酒量显著高于正常对照人群。ADH4不仅在酒精代谢过程中发挥关键作用,而且参与CH发作过程中有重要作用的下视丘素等多种单胺类神经递质的代谢[31]。
HCRTR2位于人类染色体6p12.1,包含7个外显子,能够编码主要表达在大脑组织的G-蛋白偶联受体。HCRTR2能够和下丘脑分泌素结合,而后者广泛参与中枢神经性疼痛和神经内分泌活动[32]。生物信息学分析[8]显示,HCRTR2多态性能够影响其mRNA的分子结构和生物学稳定性。
CLOCK、ADH4和HCRTR2的基础研究均表明其可能参与CH发生。虽然本研究并未发现这些基因的6个多态性位点与CH相关,但是鉴于目前研究数量较少,而且纳入的研究对象数量有限,统计学整体把握度较低,所以不能排除其和CH的临床关联性。
尽管本研究对现有的研究成果进行了合并分析,但是仍存在一些局限性:(1)由于CH发病率较低,纳入的原始研究的样本量有限,这种情况下并不能排除研究结果假阴性的可能性。Button等[33]曾报道,尤其在神经科学领域,较小的样本量很难检测到阳性结果。(2)原始研究主要在高加索人群中完成,研究结果在其他人群中的适用性仍有待检验。(3)原始研究并没有严格按照吸烟及饮酒等混杂因素对研究对象进行分层,因此不能消除这些因素对研究结果的影响。(4)本研究只对6个基因的独立位点进行了分析,而基因与基因、位点与位点之间的交互作用不能被分析,这也可能导致研究结果偏离真实情况。
综上,本研究发现CLOCK(rs1801260)、ADH4(rs1800759和rs1126671)、HCRTR2(rs2653349、rs3122156和rs2653342)多态性和CH易感性无相关性。考虑到本研究存在的缺陷,并不能排除这6个多态性位点和CH的临床关联性,仍需要在不同人群中实施大样本研究来进一步验证本研究的结果。
