油松人工林新生枝叶碳氮磷含量及化学计量比对氮添加的响应
2022-03-03杨屹宇张芸香马晓霞郭晋平
杨屹宇,崔 爽,张芸香,马晓霞,郭晋平
(1.山西农业大学 林学院,山西太谷 030800;2.北方功能油料树种培育与研发山西省重点实验室,山西太谷 030800;3.山西省桑干河杨树丰产林实验局科技服务中心,山西怀仁 038300)
碳(C)、氮(N)和磷(P)是植物细胞结构组成和功能代谢最重要的元素,是植物生长过程中必不可少的基本营养元素,在植物生长和各种生理调节过程中发挥重要作用[1]。作为重要的化学计量特征指标,C、N 和P 含量及其化学计量比可揭示植物不同器官养分含量和养分限制关系;其中,C/N 和C/P 反映植物的生长速率及植物自身对N 和P 的利用效率[2],N/P 广泛用于植物个体、群落及生态系统中N和P 限制格局的判断[3-4]。N 是植物生长必需的大量元素,在森林生态系统中最易耗竭,是限制植物生长的主要元素[5]。施N对植物生长、代谢及植物体内C、N 和P 养分调节机制有重要影响。适量施肥能在一定程度上缓解土壤N 含量不足的状况,促进植物生长[6];过量施肥则会导致土壤酸化、抑制凋落物分解[7]等,进而影响生态系统的稳定性。施肥量如何影响树体各器官C、N 和P 含量及其化学计量比,林龄对树体各器官C、N和P含量的影响以及施肥对各器官生长限制的影响等问题,还有待深入研究。
油松(Pinus tabuliformis)是我国特有树种,广泛分布于北方山地,常用于暖温带地区人工造林[8],也是水源涵养林主要造林树种。李素新等[9]开展黄土高原地区外源N 添加改变土壤性状,进而影响植物生长发育等问题的研究,结果显示由于外源N 的持续增加,植物内稳态发生显著变化,C/N 显著减小,N/P、C/P 显著增加。李瑞瑞等[10]研究N 添加对墨西哥柏(Cupressus lusitanica)人工林土壤化学计量的影响,结果显示随N添加量的增加,土壤有机碳含量显著增加,总氮(TN)含量表现为先升高后降低,总磷(TP)含量无显著变化,C/N、C/P 显著高于对照样地。Sistla 等[11]通过Meta 分析发现,N 添加显著降低植物C/P,显著增加植物N/P。长期N 添加对油松人工林针叶性状影响显著[12];子午岭林区不同林龄(9、23、33 和47年)油松人工林土壤有机C 和TN 含量均随林龄增加呈先增加后减小的趋势,P 含量随林龄增加呈先减少后增加的趋势[13]。以上研究主要集中于土壤养分对N的响应。
本研究以不同林龄油松人工林为研究对象,进行全生长季野外施N 对比试验,对不同林龄油松人工林新生枝叶总碳(TC)、TN 和TP 含量及其化学计量比的变化规律进行研究,以期揭示油松人工林N利用策略及其对N 添加的响应,为立地生产力维护和林地管理提供依据。
1 材料与方法
1.1 试验地概况
试验地位于山西省杨树丰产林实验局金沙滩林场(112°58′ ~113°10′E,39°37′ ~39°40′N),属温带大陆性季风气候,海拔970 ~1 076 m,年均降水量380 mm,年均气温6.8 ℃,年均蒸发量1 840 mm,年均日照时长1 900 h,无霜期120 ~126 天;土壤为淡栗钙土,成土母质为风积沙土。乔木主要有油松、刺槐(Robinia pseudoacacia)、樟子松(Pinus sylvestrisvar.mongolica)和榆树(Ulmus pumila)等;灌木主要有柠条锦鸡儿(Caragana korshinskii)、百里香(Thy⁃mus mongolicus)和锦鸡儿(Caragana sinica)等;草本植物主要有艾(Artemisia argyi)、狗尾草(Setaria viri⁃dis)、羊胡子草(Eriophorum scheuchzeri)和披碱草(Elymus dahuricus)等。
1.2 研究方法
1.2.1 样地设置
2020年10月,经实地调研和踏查,选择20(幼龄林)、29(中龄林)、37(近熟林)和48(成熟林)年生油松人工林纯林,油松占比不低于90%,立地条件相近。在各林龄林分中,布设面积为20 m×20 m 样地5块。测定样地海拔、坡度和土壤质地,测定并记录树种、胸径、树高、郁闭度和冠幅等林分因子,调查林下主要灌木和草本植物种类,取样并测定土壤pH值(表1)。
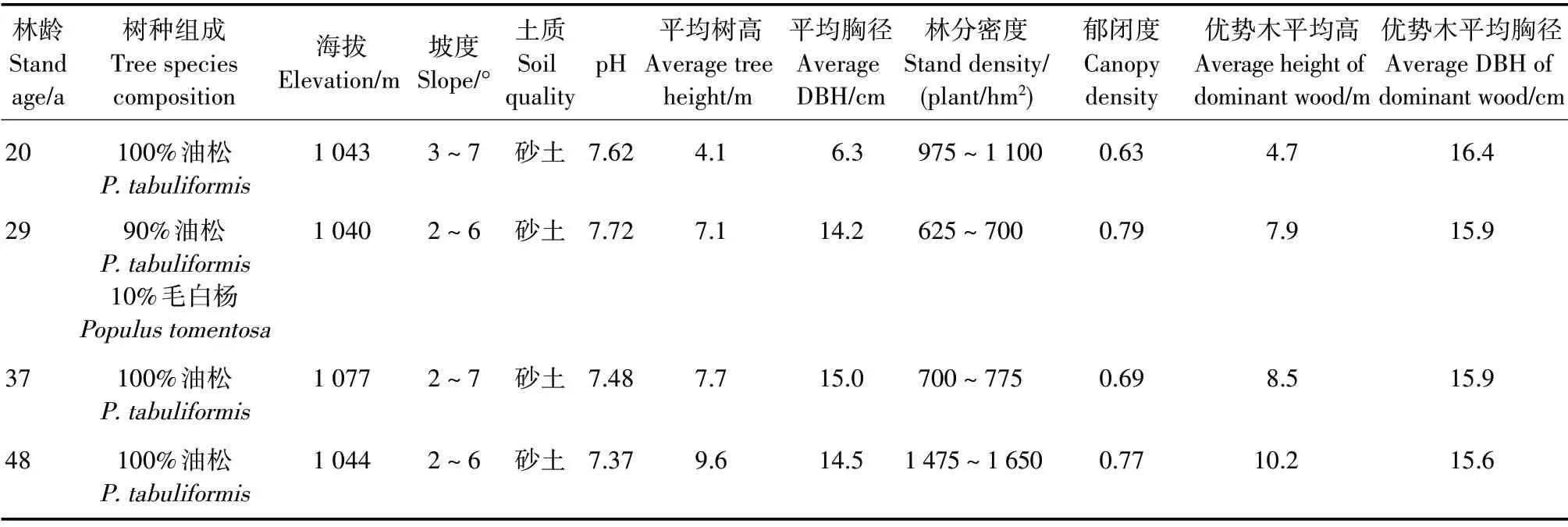
表1 样地初始林分特征和土壤理化性质Tab.1 Initial stand characteristics and soil physical and chemical properties of sample plots
1.2.2 试验设计
以尿素为N 源。在每个林分的5 块样地中设置不同的N 添加量,分别为0、5、10、15和20 g/m2,其中0 g/m2为对照(CK)。根据调查结果,在样地内选定3株优势木(树干通直、顶梢完整、无分叉、无受伤且无明显被压现象)为重复样株,共测定60 株样株。施肥处理在样地设置后立即进行(2020年10月),将设计施用量溶于20 L 水中,均匀喷洒至样地内,CK样地喷洒等量清水。所有处理范围扩大至样地外10 m。
1.2.3 样品采集
2021年5月10日取样。在选定样株的树冠上层东、西、南和北4 个方向,用高枝剪各剪取当年生枝1条,摘取全部完全伸展、无枯黄且无病虫害的完整针叶,充分混合均匀,采用四分法取约500 g样品;取摘除针叶后的当年生枝,剪短后充分混合,采用四分法取约500 g样品。将样品带回实验室。
1.2.4 样品处理与测定
将枝和针叶样品置于烘箱65 ℃烘干至恒重,粉碎过100目筛,封存备用。
TC 含量采用TOC 分析仪(multi N/C 2100 分析仪,德国耶拿)测定;TN 含量采用凯氏定氮法[14]测定,TP含量采用钼锑抗比色法[15]测定。
1.3 数据处理
采用Excel 2010 和SPSS 26.0 软件进行数据统计与分析,采用单因素方差分析法(One-way ANO⁃VA)分析林龄和N 添加量及其交互作用对油松新生枝叶C、N和P含量及其化学计量比的影响。
2 结果与分析
2.1 新生枝C、N和P含量N添加效应和林龄差异
林龄、N 添加量及其交互作用对新生枝TC 含量均影响不显著;林龄、N添加量及其交互作用对新生枝TN含量均影响极显著(P<0.001);林龄对新生枝TP含量影响显著(P<0.05),N 添加量及林龄和N 添加量交互作用对新生枝TP 含量均影响极显著(P<0.001)(表2)。各林龄新生枝TC、TN和TP含量均随N 添加量增加呈先增加后减少的趋势(图1)。20 和29年生新生枝TC 含量均在N 添加量为15 g/m2时最高,分别为610.93 和632.47 mg/g;37 和48年生新生枝TC 含量均在N 添加量为10 g/m2时最高,分别为623.42 和618.70 mg/g。20、29、37 和48年生新生枝TN 含量均在N 添加量为15 g/m2时最高,分别为7.02、7.52、6.94 和7.54 mg/g。20、29、37 和48年生新生枝TP 含量均在N 添加量为10 g/m2时最高,分别为1.91、1.87、1.61 和1.65 mg/g。新生枝TC 和TN 含量均随林龄增加呈先增加后减少的趋势,29年生时较高;新生枝TP含量随林龄增加呈先减少后增加的趋势,37年生时较低。
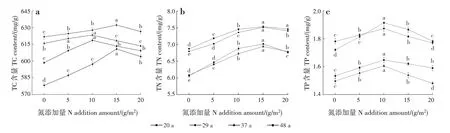
图1 各林龄新生枝C、N和P含量随N添加量的变化Fig.1 Changes of C,N and P contents in new branches at different stand ages with different N addition amounts
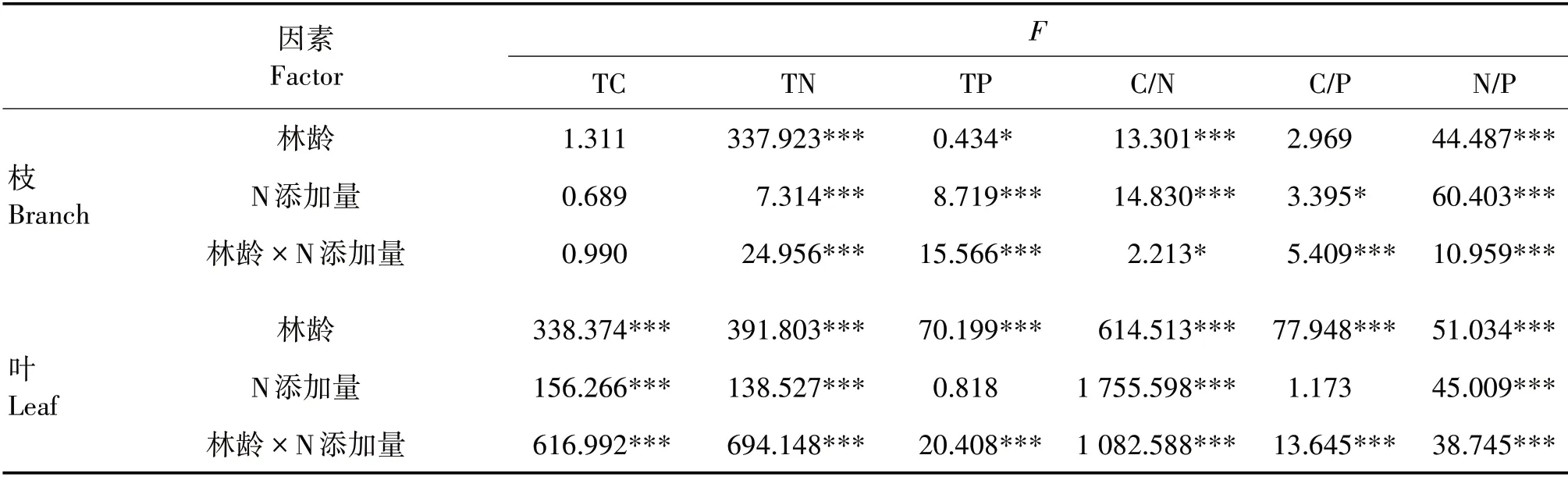
表2 林龄与N添加量对新生枝叶C、N和P含量及其化学计量比的影响Tab.2 Effects of stand ages and N addition amounts on contents of C,N and P and their stoichiometric ratios in new branches and leaves
2.2 新生枝C、N 和P 化学计量比N 添加效应和林龄差异
林龄和N添加量对新生枝C/N 均影响极显著(P<0.001),其交互作用影响显著(P<0.05);林龄对新生枝C/P 影响不显著,N 添加量影响显著(P<0.05),其交互作用影响极显著(P<0.001);林龄、N添加量及其交互作用对新生枝N/P均影响极显著(P<0.001)(表2)。新生枝C/N 和C/P 均随N 添加量增加呈先下降后升高的趋势(图2)。29、37 和48年生新生枝C/N 均在N 添加量为15 g/m2时最低,分别为84.12、89.07和81.40,20年生新生枝C/N 在N添加量为10 g/m2时最低(86.77)。20、29、37和48年生新生枝C/P 均在N 添加量为10 g/m2时最低,分别为312.00、335.19、388.33和375.46。各林龄新生枝N/P随N 添加量增加而升高。新生枝C/N 和C/P 均随林龄增加呈先升高后下降的趋势,37年生时较高;新生枝N/P随林龄增加而升高。

图2 各林龄新生枝C、N和P化学计量比随N添加量的变化Fig.2 Changes of stoichiometric ratios of C,N and P in new branches at different stand ages with different N addition amounts
2.3 新生叶C、N和P含量N添加效应和林龄差异
林龄、N添加量及其交互作用对新生叶TC和TN含量均影响极显著(P<0.001),林龄及林龄和N添加量的交互作用对新生叶TP 含量均影响极显著(P<0.001),N 添加量对新生叶TP 含量影响不显著(表2)。各林龄新生叶TC、TN 和TP 含量均随N 添加量增加呈先增加后减少的趋势(图3)。20年生新生叶TC 含量在N 添加量为15 g/m2时最高(600.80 mg/g),29、37和48年生新生叶TC含量在N添加量为10 g/m2时最高,分别为622.94、621.90 和612.10 mg/g;20、29、37 和48年生新生叶TN 含量均在N 添加量为15 g/m2时最高,分别为9.55、9.53、8.61 和8.77 mg/g;20、29、37 和48年生新生叶TP 含量均在N 添加量为10 g/m2时最高,分别为2.06、1.96、1.60和1.98 mg/g。新生叶TC含量随林龄增加呈先增加后减少的趋势,29和37年生时较高;TN 和TP 含量均随林龄增加呈先减少后增加的趋势,均在37年生时较低。
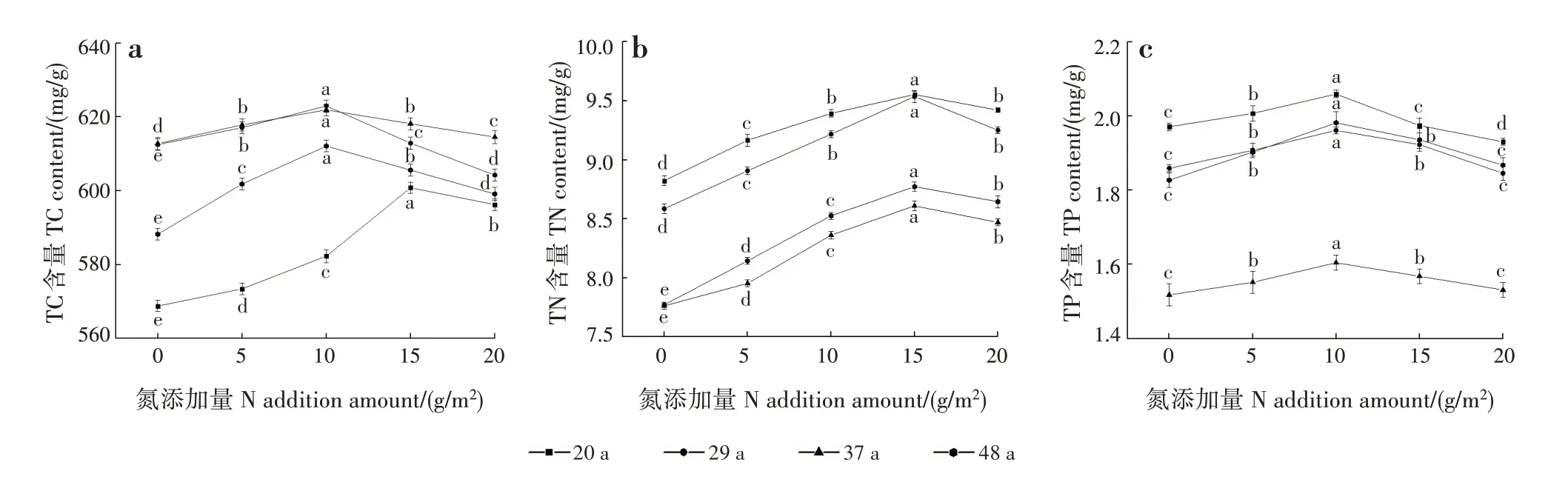
图3 各林龄新生叶C、N和P含量随N添加量的变化Fig.3 Changes of C,N and P contents in new leaves at different stand ages with different N addition amounts
2.4 新生叶C、N 和P 化学计量比N 添加效应和林龄差异
林龄、N添加量及其交互作用对新生叶C/N和N/P均影响极显著(P<0.001),林龄及林龄和N添加量的交互作用对新生叶C/P 均影响极显著(P<0.001),N 添加量对新生叶C/P 影响不显著(表2)。各林龄新生叶C/N 和C/P 均随N 添加量增加呈先下降后升高的趋势;N 添加量为10 g/m2时,20、29、37 和48年生新生叶C/P 及20年生新生叶C/N 最低,分别为282.81、317.68、387.61、308.90 和61.99;29、37 和48年生新生叶C/N 在N 添加量为15 g/m2时最低,分别为64.27、71.80 和69.03(图4)。各林龄新生叶N/P均随N 添加量增加逐渐升高。随林龄增加,各林龄新生叶C/N、C/P 和N/P 均呈先升高后下降的趋势,均在37年生时较高。
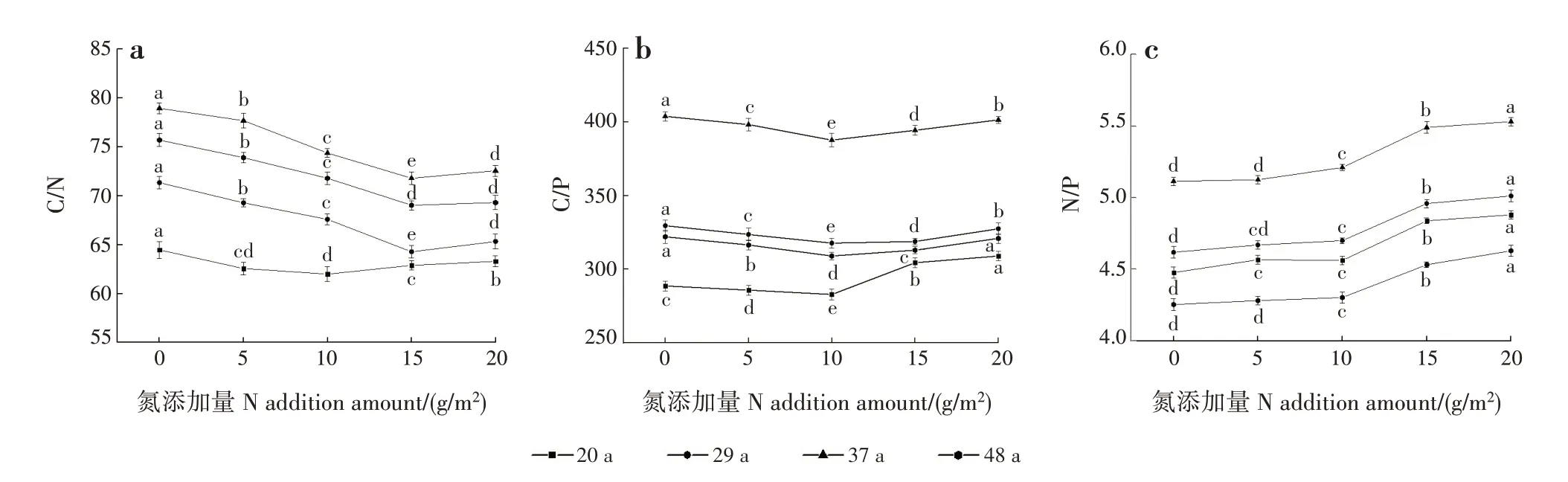
图4 各林龄新生叶C、N和P化学计量比随N添加量的变化Fig.4 Changes of stoichiometric ratios of C,N and P in new leaves at different ages with different N addition amounts
3 讨论与结论
3.1 同一林龄油松人工林新生枝叶C、N 和P 含量及其化学计量比对N添加量的响应
N 和P 在植物生长和发育的各个阶段均发挥着重要作用[16]。同一林龄下,随N 添加量的增加,油松人工林枝叶的TC、TN 和TP 含量均呈先增加后减少的趋势,这可能是由于油松N 饱和的限制性[17]。添加外源N,土壤中供植物生长所需的有效N 含量增加,枝叶中的N含量随之升高[18];植物对N 的吸收达到饱和状态时,继续添加N会对植物产生负面影响,造成N 流失。N 添加量增加可促进叶片光合作用和根系生长,根系可为叶片提供生长所需的C 和N,维持植物较高的光吸收能力,增加C 含量[19];随N 添加量持续增加,土壤中N 含量过高,抑制各器官的生长,TC 含量减少[20]。持续的N 添加使得植物通过自身调节所诱导的P 的有效性不能满足其对P 养分的需求,植物需消耗更多的P以弥补对N 元素的吸收,导致叶片P 含量减少。本试验地的土壤类型为砂土,土壤养分含量较低,添加外源速效养分后,油松吸收部分养分;但随N添加量增加,土壤中的N出现饱和,枝叶生长所需的N也开始下降,出现了高N降低枝叶TN 含量的情况,新生枝叶的TN 含量减少[21]。本次取样时间为5月,处于林木生长初期,枝叶对土壤中养分的吸收不完全,结果可能出现偏差,部分结论有待继续研究。
N/P 普遍用于判断外界环境对植物N 和P 养分的限制情况;植物叶片N/P >16 时,认为该植物受P限制;N/P 为14 ~16 时,认为该植物生长受N 和P 同时限制;N/P <14 时,认为该植物受N 限制[22]。本试验中,油松人工林新生枝叶的N/P 均随N 添加量增加持续升高,且各林龄新生枝叶的N/P 均在3 ~6 范围内,均小于14,油松人工林生长受N限制严重。随N 添加量增加,土壤中N 的有效性增强,缓解了N 限制,增加了P限制,总体上N/P呈升高趋势[23]。C/N和C/P代表植物同化C的能力。本研究中,油松人工林新生枝叶的C/N 均随N 添加量增加呈先下降后升高的趋势,可能是由于N 含量上升或养分较充足,N 利用率下降[24]。油松人工林新生枝C/N 和C/P 比叶高是因为枝内含有大量富C类的次生代谢产物[25-26]。
3.2 不同林龄油松人工林新生枝叶C、N 和P 含量及其化学计量比对N添加量的响应
除新生枝TC 含量和C/P 外,油松人工林新生枝叶的TC、TN 和TP 含量及其化学计量比均随林龄变化差异显著,可能是因为C 不是植物生长的限制元素,且在植物体内的存在方式多为纤维素等多糖类物质,受环境影响远小于N 和P 等功能性物质[27]。幼龄林生长速度较快,需要大量核糖体合成蛋白质,核糖体是植物体中重要的P 库,本研究中20年生油松幼龄林新生枝叶的TP含量较高。本研究中,油松人工林新生枝叶的C/N 和C/P 均随林龄增加呈先升高后下降的趋势,这可能是由于树木在幼龄林时期生长迅速,代谢活动旺盛[28]。生长速率假说认为,生物体在生长过程中,会分配大量P到rRNA中,使核糖体能快速合成蛋白质;近熟林和成熟林时,富集C 的物质积累,C/N 和C/P 升高[29]。油松人工林新生枝叶的N/P 随林龄增加持续升高(除48年生叶),表明随林龄增加油松人工林生长受N限制的情况有所缓解,这与崔宁洁等[30]对马尾松(Pinus masso⁃niana)人工林的研究结果一致。在油松人工林,特别是幼龄林的管理中,可适当增施N 肥,促进幼树生长。
3.3 结论
油松人工林新生枝叶的TC、TN 和TP 含量均随N 添加量的增加呈先增加后减少的趋势,均在10 或15 g/m2添加量下最高;枝叶的TC、TN 和TP 含量均存在林龄间差异,枝的变幅分别为7.53%、13.37%和18.67%,叶的变幅分别为7.74%、13.67%和29.61%。
油松人工林新生枝叶的C/N、C/P 和N/P 均随N添加量的变化小幅变化,平均变化幅度为3.99% ~9.91%;C/N 和C/P 均随N 添加量增加呈先下降后升高的趋势,N/P持续升高、C/P和N/P均存在林龄间差异,均随林龄增加呈先升高后下降的趋势,37年生时较高;新生枝的N/P 随林龄增加持续升高,叶的N/P则先升高后降低。
各林龄油松人工林新生枝叶的N/P 在3 ~6 之间,均低于14,土壤N 限制显著,N 添加显著缓解土壤N 限制;N 添加量增加,N/P 持续升高,在所设N 添加量范围内未达到峰值。
