大麦品种(系) HvPDIL5-1和 HvEIF4E基因的单倍型及其黄花叶病抗性研究
2022-03-01潘雨涵栾海业郭宝健沈会权许如根
潘雨涵,洪 益,徐 肖,栾海业,朱 娟,吕 超,郭宝健,沈会权,许如根
(1.植物功能基因组学教育部重点实验室/江苏省作物基因组学与分子育种重点实验室/江苏粮食作物现代产业技术协同创新中心/扬州大学农业科技发展研究院,江苏扬州 225009; 2.江苏沿海地区农业科学研究所,江苏盐城 224001)
大麦黄花叶病是由大麦黄花叶病毒(barley yellow mosaic virus, BaYMV)、大麦温和花叶病毒(barley mild mosaic virus, BaMMV)或两种病毒复合体引起的病毒病。两种病毒均属正义单链RNA病毒。病毒以土壤中的禾谷多黏菌()为传播介体,冬前禾谷多黏菌侵入大麦幼苗的根部并释放出病毒,病毒随水分由根部运输至叶片;待翌年春季的气温回升到10 ℃以上时,病毒在叶片内快速增殖,破坏叶片的叶绿体造成褪绿斑点。随病毒数量的增加,褪绿斑点连成线,继而连成面,造成叶片黄化、死亡,继而光合效率下降或光合功能丧失,最终导致大麦减产,严重时可减产70%~80%。大麦黄花叶病于20世纪70年代由日本传到中国,首先在沿海和长江中下游大麦区流行,对大麦生产造成严重影响,目前该病害已蔓延到全国大部分麦区。
抗病品种的选育与推广是防治大麦黄花叶病最经济有效的途径。鉴定筛选抗病种质,发掘定位抗病基因,是培育抗黄花叶病品种的关键。大麦黄花叶病是由主基因控制的性状,随着病毒株系的进化与变异,原有抗性品种的抗性逐渐丧失,因此需要不断发掘新的大麦黄花叶病抗性基因并在育种中加以利用。目前已定位的大麦黄花叶病抗性基因共计22个,其中仅有和基因被克隆,和基因来自木石港3号,基因来自Russia 57,基因来自Ragusa。位于大麦4H染色体长臂上,为广谱抗性,抗欧洲所有大麦黄花叶病毒株系,其抗性是由于感病基因外显子区段1 375 bp的序列缺失而无法产生转录产物导致的。Yang等研究表明,基因的30种单倍型中,具有黄花叶病抗性的单倍型仅有7种。位于3H染色体末端,在大麦黄花叶病抗性育种中应用广泛,其抗性是由真核翻译起始因子基因编码的氨基酸突变导致,对BaMMV和BaYMV不同毒株的抗性存在差异,目前报道的基因的单倍型共有67种。本研究以29个大麦黄花叶病抗性存在差异的大麦品种(系)为材料,对供试品种的和基因单倍型及黄花叶病抗性进行分析,并探究供试品种黄花叶病抗性基因单倍型的亲缘关系,以期为大麦品种黄花叶病抗性改良提供依据。
1 材料与方法
1.1 试验材料
本研究以扬州大学大麦研究所育成的14个大麦品种(系)和15个黄花叶病抗性存在差异的大麦品种(系)为材料,供试材料的名称和来源见表1。
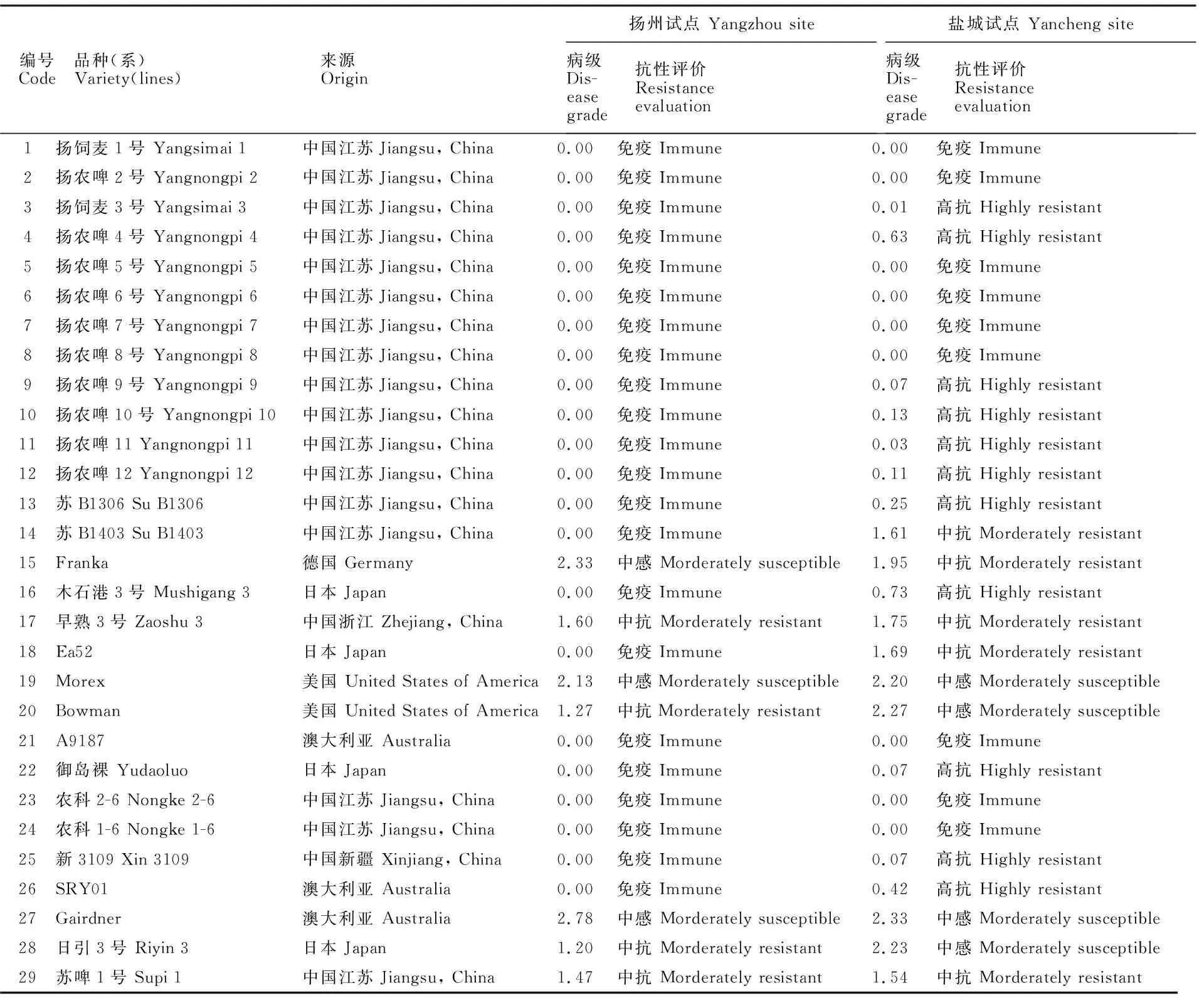
表1 供试材料的来源及其黄花叶病抗性评价Table 1 Origin and resistance evaluation of yellow mosaic disease in the test materials
1.2 黄花叶病抗性鉴定
于2019年秋季,将供试材料同时播种于扬州大学大麦黄花叶病病圃(江苏扬州,32° N,119° E)及江苏沿海地区农业科学研究所大麦黄花叶病病圃(江苏盐城,33° N,120° E)。每个材料种植1行,行长1.5 m,行距0.2 m,每行人工均匀点播10粒,3次重复。
于2020年春季,参照大麦黄花叶病鉴定标准,对供试材料进行大田黄花叶病自然抗性鉴定。每行连续调查5株,取平均值。扬州试点调查6期(2020年2月2日、2月10日、2月18日、2月24日、3月4日和3月15日),取平均值;盐城试点调查5期(2020年2月9日、2月18日、2月24日、3月4日和3月15日),取平均值。根据发病的病级判断黄花叶病抗性,具体标准:平均病级=0.00,免疫;平均病级=0.00~1.00,高抗;平均病级=1.00~2.00,中抗;平均病级=2.00~ 3.00;中感;平均病级=3.00~4.00,高感。
1.3 HvPDIL5-1和 HvEIF4E基因的扩增与纯化
供试材料均采用植物基因组DNA小量提取法提取DNA。采用20 μL PCR扩增体系,包括2 μL (40 ng)基因组DNA、2 μL上、下游引物(表2)、10 μL 2×Green Taq Mix (南京诺唯赞生物科技有限公司)和6 μL ddHO。参考Don等开发的Touch-down PCR程序,稍加改变进行扩增,即94 ℃预变性5 min;94 ℃变性30 s, 66 ℃退火30 s,72 ℃延伸1~2 min,退火温度每循环1次降低1 ℃,直到60 ℃;保持94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸1~2 min,循环35次;72 ℃延伸10 min,4 ℃冷却30 s。

表2 本研究所用的引物信息Table 2 Primer information in this study
将目的片段在紫外灯下快速切割,用北京全式金生物公司提供的Easy Pure Quick Gel Extraction试剂盒对供试材料的目的片段进行回收纯化,纯化片段溶于20 μL EB缓冲液中,用NanoDrop进行QC检测和浓度测定,供试材料1~25(名称见表1)送北京博迈德基因技术有限公司进行Sanger测序。供试材料SRY01、Gairdner、日引3号和苏啤1号的目的片段纯化后送至华大基因公司进行Sanger测序。
1.4 数据分析
在NCBI网站下载已公开的大麦基因和的CDS序列,利用Sequencher 5.4.6软件对测序结果进行整理分析,依据Yang等的研究结果,对和基因单倍型的抗性进行判断。依据的单倍型,利用MEGA X软件的Neighbor-joining算法对该基因在供试品种间的亲缘关系进行分析。
2 结果与分析
2.1 供试大麦品种(系)的黄花叶病抗性表现
由表1可知,2019-2020年度在扬州和盐城试点,大麦黄花叶病发病普遍较轻,其中高感大麦黄花叶病品种早熟3号和苏啤1号在两试点均表现为中抗,比早熟3号和苏啤1号更感病的品种(系)有Franka、Morex、Bowman、Gairdner、日引3号,其中Gairdner病级最重,在扬州和盐城试点分别为2.78和2.33。其他22个品种(系)在扬州试点均表现为免疫。其中,有9个品种(扬饲麦1号、扬饲麦2号、扬农啤5号、扬农啤6号、扬农啤7号、扬农啤8号、A9187、农科2-6和农科1-6)在盐城试点也表现为免疫,其他13个品种(系)在盐城试点均有不同程度地感病。
2.2 HvPDIL5-1基因的扩增、回收及测序结果
用引物PDI_45_2138对供试材料进行扩增,结果如图1A和图1B所示,可以看出,29个材料均能扩增出全CDS区片段,全长2 093 bp。其中22个材料可扩增出明亮的目的条带,符合回收纯化测序的要求,对目的片段进行回收、纯化及测序,均获得测序结果。但有7份材料(扬农啤8号、早熟3号、新3109、SRY01、Gairdner、日引3号和苏啤1号)的条带均较弱,达不到纯化、回收及测序的要求。因此,进一步用引物Till_M56_957和Till_1749_2200对达不到纯化回收测序要求的7个材料的全CDS区片段进行扩增,均能扩增出1 013 bp和451 bp的目的条带,扩增结果如图1C和图1D所示。2对引物均能扩增出明亮的目的条带,符合回收、纯化及测序的要求,对目的片段进行回收、纯化及测序,均获得测序结果。

M1:5 000 bp DNA Marker;M2:2 000 bp DNA Marker。1~29所对应的品种(系)名称见表1。图3同。M1:5 000 bp DNA Marker; M2:2 000 bp DNA Marker. The names of varieties(lines) corresponding to 1-29 are shown in table 1. The same in figure 3.图1 29份材料 HvPDIL5-1基因的扩增结果Fig.1 Amplification results of HvPDIL5-1 gene in the 29 varieties(lines)
2.3 供试材料 HvPDIL5-1基因的单倍型与抗性
由图2可知,仅木石港3号、A9187和SRY01中基因的外显子区存在SNP突变,其中,木石港3号在182 bp处发生了G→A的非同义突变,为单倍型hap-VII(),该突变使编码色氨酸的密码子TGG突变为TAG(终止密码子),导致感病基因提前终止编码,感病功能丢失,获隐性黄花叶病抗病性;A9187中基因的外显子区存在2个SNP突变位点,分别在138 bp和374 bp处发生了A→T的同义突变和C→G的非同义突变,其编码氨基酸由脯氨酸突变为精氨酸,但不影响基因编码蛋白质的功能,为感病单倍型hap-XIII;SRY01在26 bp处发生了C→A的非同义突变,其编码氨基酸由丝氨酸突变为色氨酸,但不影响基因编码蛋白质的功能,为感病单倍型hap-XVII。
黄色高亮标记表示SNP突变位点。图4同。右侧红色和绿色罗马数字分别表示感病单倍型和抗病单倍型。SNPs are highlighted in yellow. The same in fugure 4. Red and green Roman numerals on the right indicate the haplotypes of susceptible and resistant, respectively.图2 供试材料 HvPDIL5-1单倍型与黄花叶病抗性Fig.2 Haplotypes of HvPDIL5-1 and resistance to yellow mosaic disease in the test materials
2.4 HvEIF4E基因的扩增、回收与测序结果
利用3个分段引物(eIF4e_m56 s/309as、eIF4e_1659 s/2394as和eIF4e_4857 s/5396as)对29个大麦供试材料的基因进行扩增,结果如图3A和图3B所示,由图3可以看出,所有供试材料均能扩增出目的条带,引物eIF4e_m56 s/309as可扩增出365 bp的目的条带,引物eIF4e_1659 s/2394as可扩增出735 bp的目的条带,引物eIF4e_4857 s/5396as可扩增出539 bp的目的条带,所有条带均达到回收、测序要求,对目的片段进行回收、纯化及测序,均获得测序 结果。

图3 29份材料 HvEIF4E基因的扩增结果Fig.3 Amplification results of HvEIF4E gene in 29 varieties(lines)
2.5 供试材料 HvEIF4E基因的单倍型与抗性
以大麦基因的野生型hap-wt0A为参考序列进行比对,结果(图4)发现,供试材料基因外显子区的12个位置出现14种SNP突变,所有SNP均为非同义突变,产生14种单倍型。其中,感病单倍型有:扬农啤7号、Morex、Bowman和新3109的野生型hap-wt0A;早熟3号的单倍型hap-I;扬饲麦3号、木石港3号、农科2-6和农科1-6的单倍型hap-II;Franka的单倍型hap-IV;扬农啤2号和扬农啤8号的单倍型hap-XIV;扬农啤10号和扬农啤11的单倍型hap-XX;Gairdner的单倍型hap-XXXVI;A9187的单倍型hap-XLIV;SRY01的单倍型hap-L。抗病单倍型有:扬农啤9号的单倍型hap-XIII;日引3号和苏啤1号的单倍型hap-XV;扬农啤4号、扬农啤5号、扬农啤6号、扬农啤12、苏B1306、苏B1403和Ea52为抗病单倍型hap-(携带抗大麦黄花叶病基因)。抗性未知单倍型有:扬饲麦1号的单倍型hap-XII和御岛裸的单倍型hap-XVI。
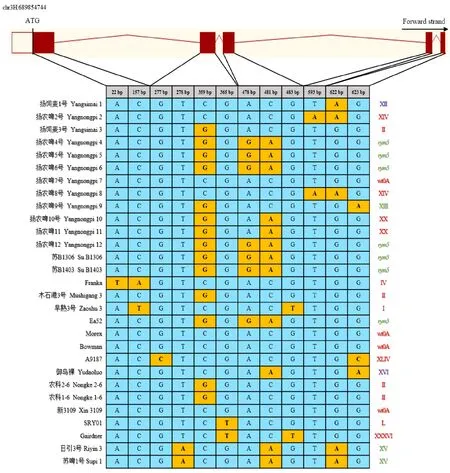
右侧红色、绿色和紫色罗马数字分别表示感病单倍型、抗病单倍型和未知抗性单倍型。Red, green and purple Roman numerals on the right represent haplotypes of susceptible, resistant and unknown resistance, respectively.图4 供试材料 HvEIF4E基因的单倍型与黄花叶病抗性Fig.4 Haplotypes of HvEIF4E gene and resistance to yellow mosaic disease in the test materials
2.6 供试材料黄花叶病抗性基因 HvEIF4E单倍型的亲缘关系
根据黄花叶病抗性基因的单倍型,利用MEGA X软件构建供试材料的进化树(图5)。其中,单倍型为hap-XIII的扬农啤9号单独聚在一枝,与单倍型为hap-XV的日引3号和苏啤1号聚在一个大分枝上,且这两类单倍型均为抗病型,说明这两类抗病单倍型亲缘关系较近;单倍型为hap-的扬农啤4号、扬农啤5号、扬农啤6号、扬农啤12、苏B1306、苏B1403和Ea52聚在一个分枝上,与单倍型为hap-XX的扬农啤10号和扬农啤11聚在一个大分枝上,说明单倍型hap-和hap-XX亲缘关系较近,而hap-为抗病单倍型,hap-XX为感病单倍型,具体原因尚待分析;扬饲麦1号为抗性未知单倍型hap-XII,单独聚在一枝,与单倍型为hap-XIV的扬农啤2号与扬农啤8号聚在一个大分枝上,而hap-XIV为感病单倍型,推测hap-XII也为感病单倍型;而单倍型为hap-XVI的御岛裸单独聚为一枝,其亲缘关系有待进一步研究。

图5 基于供试材料 HvEIF4E基因的单倍型构建的的进化树Fig.5 Phylogenetic tree of the tested materials based on HvEIF4E haplotype
3 讨 论
3.1 野生大麦可能是黄花叶病抗性基因的来源之一
基因的所有单倍型中,有7种单倍型抗大麦黄花叶病毒株系BaMMV-ASL,包含一个来自野生大麦()的单倍型hap-XVIII,表明野生大麦是大麦黄花叶病抗性基因的来源之一。Yang等对1 816份大麦种质基因的单倍型进行分析,发现9份来自中东地区的二棱野生大麦种质属于单倍型hap-XIII,单倍型hap-XIII种质在人工接种BaMMV-ASL后表现为感病;同时,在半野生大麦A9187基因外显子区段检测到对BaMMV-ASL表现为感病的单倍型hap-XLIV。本研究发现,A9187的基因在外显子区存在同义突变和非同义突变,为单倍型hap-XIII,且在扬州和盐城试点对大麦黄花叶病均表现为免疫(平均病级=0.00),说明A9187虽然不具有BaMMV-ASL病毒抗性,但存在抗江苏大麦黄花叶病病毒的抗性基因,可进一步发掘该抗病基因,为江苏大麦黄花叶病抗性品种的培育提供帮助。
Yang等用BaMMV-ASL接种不同来源的大麦种质,检测到基因的感病单倍型hap-XVII和基因的感病单倍型hap-L均来自大麦野生种。本研究在半野生大麦SPY01中也检测到这两种感病单倍型。但该品种在扬州病圃对大麦黄花叶病表现为免疫(平均病级= 0.00),在盐城病圃表现为高抗(平均病级= 0.47),说明半野生大麦SRY01对BaMMV-ASL病毒不具有抗性,但存在抗江苏大麦黄花叶病病毒的抗性基因,且其抗性基因不同于和抗性单倍型。
3.2 HvPDIL5-1和 HvEIF4E基因单倍型与供试品种黄花叶病抗性的关系
本研究发现,29个供试大麦品种(系)中,只有木石港3号基因的单倍型hap-XIII对大麦黄花叶病表现为抗病,其余材料的单倍型均表现为感病;29个供试大麦品种(系)的14种单倍型中,17个品种(系)有hap-wt0A、hap-I、hap-II、hap-IV、hap-XIV、hap-XX、hap-XXVI、hap-XLIV和hap-L共9种感病单倍型,10个品种(系)有hap-XIII、hap-XV和hap-共3种抗病单倍型,分别有1个品种(系)为hap-XII和hap-XV 抗性未知单倍型。从大麦黄花叶病病圃抗性鉴定结果来看,29个供试大麦品种(系)中,有20个品种(系)在扬州和盐城均表现为免疫或高抗,2个品种(系)在扬州表现为免疫、在盐城表现为中抗。从基因单倍型和供试品种(系)的抗性来看,具有抗性单倍型的品种在江苏不一定抗大麦黄花叶病,甚至表现为中感,如日引3号基因检测到抗病单倍型hap-XV,但在扬州和盐城分别表现为中抗和中感,说明江苏扬州和盐城病圃存在不同的致病 株系。
来自中国木石港3号的是欧洲黄花叶病抗性育种中的主要抗源,高抗大麦黄花叶病毒主要的株系BaMMV-ASL、BaYMV-1和BaYMV-2。啤酒大麦品种(系)扬农啤4号、扬农啤5号、扬农啤6号、扬农啤12、苏B1306和苏B1403均携带抗性基因,在扬州病圃对大麦黄花叶病均表现为免疫,而除扬农啤5号和扬农啤6号外的4个品种(系)在盐城病圃对大麦黄花叶病表现为中抗或高抗。大麦黄花叶病抗性基因较多,每一个抗性基因可能只抗一种或几种病毒株系,且大麦黄花叶病的病毒株系在不断变异,不断产生新的病毒株系,基因的抗病性被克服也被报道过。2017年之前扬州试点鉴定的抗性品种在盐城试点对大麦黄花叶病也表现为抗病,但本研究发现,近几年出现了在扬州试点对大麦黄花叶病表现为免疫,而在盐城试点表现为高抗或中抗的品种,说明盐城试点可能传入或产生新的病毒株系,需不断发掘黄花叶病抗性基因,以保证大麦黄花叶病抗性育种的可持续性。
本研究29个供试大麦品种(系)中,扬农啤2号、扬饲麦3号、扬农啤7号、扬农啤8号、扬农啤10号、扬农啤11、A9187、农科2-6、农科1-6和SRY01在扬州和盐城试点对大麦黄花叶病均表现为免疫或高抗,但其和基因均未检测到抗黄花叶病的单倍型,说明这些品种(系)中含有除和以外的黄花叶病抗性基因,有待进一步研究与发掘;木石港3号基因检测出抗性单倍型hap-VII(-d),基因检测出感病单倍型hap-II,且单倍型hap-II中只有1个与一致的SNP位点,而Yang等研究发现,共有3个SNP位点,因此还需进一步试验研究。
