鸡TRAF6基因外显子区功能性SNP预测与分析
2022-02-16赵采芹,王燕碧,唐宏,邢静如,石海英,段志强
赵采芹,王燕碧,唐宏,邢静如,石海英,段志强

摘要:采用生物信息學方法筛选鸡TRAF6基因中的功能性非同义单核苷酸多态性(nsSNPs)位点。从dbSNP数据库中检索出TRAF6基因的8个nsSNPs,利用生物信息学软件PROVEN、SIFT、PhDSNP和SNAP2分别对nsSNPs进行功能性预测;使用IMutant2.0和Mupro对突变位点氨基酸的稳定性进行预测;此外,利用Clustal Omega和Jalview对鸡TRAF6基因编码的氨基酸序列进行多序列比对和进化位点保守性预测。使用MutPred2预测突变可能造成的影响;最后使用Sopma和SWISSMODEL软件分别预测TRAF6野生型和突变型的蛋白质二级结构和构建它们的三级结构。结果表明:从TRAF6基因的nsSNPs位点筛选出来3个有害突变位点rs736890315(S17R)、rs734934585(C20G)和rs734774178(V472E),其中V472E是四种软件共同筛选出的突变位点。V472E突变位点可能影响鸡TRAF6蛋白的功能,且所有位点突变都会使TRAF6蛋白稳定性降低。保守性分析显示,D16G、S17R和A307T为不保守位点,C20G、S40G、S40R、T42A和V472E为保守的功能性残基;MutPred2预测结果发现,V472E位点的突变导致蛋白质的稳定性改变,其他位点的突变均无显著影响;二级结构分析结果表明,鸡TRAF6蛋白主要以无规则卷曲和α螺旋为主,V472E、S40G和C20G这三个位点上的突变都导致了无规则卷曲百分比的提高和α螺旋百分比下降;三级结构分析结果发现,TRAF6蛋白的野生型和突变体的三级结构与二级结构预测结果一致。本研究预测发现V472E位点突变可能严重影响鸡TRAF6蛋白质的结构,V472E可能是鸡TRAF6基因的潜在功能性位点。
关键词:鸡;TRAF6基因;非同义SNP;SNP功能预测;生物信息学
中图分类号:S831.2文献标识码:A
文章编号:1008-0457(2022)01-0001-010国际DOI编码:10.15958/j.cnki.sdnyswxb.2022.01.001
Prediction and Analysis of Functional SNP on Exon Region of Chicken TRAF6 Gene
Zhao Caiqin1,2,Wang Yanbi1,2,Tang Hong1,2,Xing Jingru1,2,Shi Haiying1,2,Duan Zhiqiang1,2*
(1.Key Laboratory of Plateau Mountain Animal Genetics,Breeding and Reproduction,Ministry of Education,Guizhou University,Guiyang,Guizhou 550025,China; 2.Guizhou Province Key Laboratory of Animal Genetics,Breeding and Reproduction,Guizhou University,Guiyang,Guizhou 550025,China)
Abstract:The purpose of this study was to screen the gene of chicken TRAF6 functional nonsynonymous single nucleotide polymorphisms (nsSNPs) by the bioinformatics method.In this study,the 8 nsSNPs of TRAF6 gene were retrieved from dbSNP database,and the functional prediction of nsSNPs was performed by using bioinformatics software PROVEN,SIFT,PhDSNP,and SNAP2,respectively; The amino acid stability of mutation sites was analyzed by IMutant2.0 and Mupro methods; In addition,Clustal Omega and Jalview were used to perform multiple sequence alignment and conservatively predict the evolution sites of amino acid sequences encoded by chicken TRAF6 gene,and Mutpred2 was used to predict the possible effects of mutations.Finally,Sopma and SWISSMODEL software were used to predict the secondary structure and construct the tertiary structure of TRAF6 wildtype and mutant proteins,respectively.The results showed that three harmful mutation sites,rs736890315 (S17R),rs734934585 (C20G),and rs734774178 (V472E),were screened out from nsSNPs sites of TRAF6 gene,of which V472E was the sites jointly screened by the four software.The V472E mutation site might affect the function of chicken TRAF6 protein and all site mutations would reduce the stability of TRAF6 protein.Conservative analysis showed that D16G,S17R,and A307T were not conserved sites,and C20G,S40G,S40R,T42A,and V472E were conserved functional residues; MutPred2 prediction results revealed that V472E mutation site led to changes in protein stability,while mutation at all other sites had no significant effects.The results of secondary structure analysis showed that the chicken TRAF6 protein was composed of the irregular coil and alpha helix.The mutations at V472E,S40G,and C20G all led to the decrease of the irregular coil and the increase of alpha helix.The tertiary structure analysis showed that the tertiary structure of the wildtype and mutant of TRAF6 protein were consistent with the predicted secondary structure.This study predicted that V472E mutations might seriously affect the structure of chicken TRAF6 protein,and V472E may be a potential functional site of the chicken TRAF6 gene.
Keywords:chicken;TRAF6 gene;nonsynonymous SNP;SNP function prediction;bioinformatics
肿瘤坏死因子受体相关因子6(TRAF6)是肿瘤坏死因子受体相关因子家族(TRAFs)中研究最为广泛的成员之一[1]。与TRAFs家族其他成员不同的是,TRAF6作为胞内重要的多功能衔接分子和E3泛素连接酶,能特异性识别氨基酸序列,从而参与白介素1受体/Toll样受体(IL1/TLR)和肿瘤坏死因子受体家族等炎症信号通路,通过激活信号通路,对免疫细胞的存活和激活及其重要[2]。此外,TRAF6蛋白在人和不同动物的组织中广泛表达,其中在免疫器官脾臟、胸腺和法氏囊中分布较多,在脑和腿肌中也有表达[35]。研究证实TRAF6蛋白不仅介导信号的传导,还在免疫应答,以及淋巴结、细胞分化与凋亡、多种组织生长发育等过程中也起着关键作用[4,6]。
大量研究表明,基因多态性与许多疾病的发生相关。Strickson等[7]将小鼠野生型TRAF6的第74位氨基酸L突变为H,小鼠可以存活5周,之后小鼠因为皮肤和器官出现炎症而死亡。与野生型小鼠相比,突变后的L74H没有出现骨质疏松的现象且与野生型的巨噬细胞相比,信号传导和抗炎因子的分泌都减弱了,在TRAF6敲除的小鼠胚胎来源的巨噬细胞中,无法诱导信号传导,也没有细胞因子的产生。陈子文[8]发现,TRAF6基因多态性位点rs5030411、rs5030416可能参与调节缺血性中风风痰癖阻证的炎性反应过程,从而影响缺血性中风的发病机制。有研究者发现,位于染色体11p12处的TRAF6基因与类风湿关节炎(RA)和系统性红斑狼疮(SLE)相关[910],TRAF6基因内含子处rs540386多态性与自身免疫疾病表型存在关联,TRAF6基因rs331457多态性可降低皮肤恶性黑色素瘤出现的风险[11]。此外,有研究报道,TRAF6基因缺乏会导致胸腺基质异常发育,从而改变免疫耐受性[12]。总之,TRAF6作为一种多功能细胞因子,以上研究表明TRAF6在多种疾病的免疫应答中起着调控作用。
遗传变异可能会改变蛋白质结构,从而导致各种疾病的发生,由此可见,遗传变异与畜禽的抗病育种存在密切关系[13]。而单核苷酸多态性(SNP)是遗传变异的主要形式,预测非同义单核苷酸多态性(nsSNPs)与疾病相关关系的研究在近些年也越来越多。nsSNPs通常被认为是导致畜禽表型和基因型变异的重要因素[1415]。nsSNPs又可分两类:若使提前终止蛋白质翻译,而不改变所编码的氨基酸序,则称为无义突变;若导致氨基酸序列发生改变,则称为错义突变,突变位点与疾病的发生相关[16]。
近年来,对TRAF6的研究多见于人、小鼠和畜禽[1719],此外,也存在于许多非脊椎动物如青蛤[20]以及斑马鱼等鱼类[21]。而目前对于TRAF6基因的多态性研究大多是其表达对于人类疾病的影响[22],而对禽类TRAF6基因nsSNPs的功能研究报道较少。本研究利用生物信息软件对鸡TRAF6基因编码区nsSNPs进行分析,筛选其中可能具有潜在生物学功能的影响鸡抗病育种的候选功能性错义突变位点,并预测其可能的作用机理。筛选和分析与鸡免疫相关的TRAF6基因nsSNPs位点,为后续开展鸡抗病育种工作提供理论参考。
1材料与方法
1.1鸡TRAF6基因nsSNPs数据的收集
根据TRAF6基因的序列号(ENSGALT0000010 5433)从Ensembl genome browser(http://asia.ensembl.org)数据库中进行检索。由dbSNP(https://ncbiinsights.ncbi.nlm.nih.gov/tag/dbsnp/)数据库[23]检索的信息收集各SNPs在基因中位置,筛选出位于编码区的nsSNPs;从dbSNP数据库中检索收集nsSNPs(rs IDs);从UniProtKB数据库(http://www.uniprot.org/)检索氨基酸序列;并使用Origin软件绘制TRAF6基因的SNPs分布图。
1.2鸡TARF6基因功能性nsSNPs预测
利用SIFT(http://sift.jcvi.org/)[24]、PROV EAN(http://provean.jcvi.org/seq_submit.php)[25]、
PhDSNP(https://snps.biofold.org/phdsnp/phdsnp.html)[26]、SNAP2(https://www.rostlab.org/services/SNAP/)[27]四种软件预测分析各nsSNPs对鸡TRAF6蛋白功能的影响,运用R语言绘制四种软件预测结果的韦恩图。
SIFT是一种主要利用SIFT算法,基于氨基酸序列同源性预测氨基酸替代是否影响蛋白功能的软件[24]。PROVEAN是一款对单个或多个氨基酸替换、插入和缺失进行预测的开放性工具[25]。PhDSNP是一种支持向量机的分类器,基于SVM算法,预测分析目标nsSNPs与疾病是否相关[26]。SNAP2是一种识别有影响和中性变异的工具[27]。
1.3鸡TRAF6蛋白质稳定性分析
利用IMutant2.0(https://folding.biofold.org/imutant//pages/IMutant2.0_TutSeqVal.html)[28]和Mupro(http://mupro.proteomics.ics.uci.edu/)[29]在线软件预测nsSNP引起蛋白质稳定性的改编。IMutant2.0是一个基于支持SVM的工具,可以自动预测单点突变时蛋白质稳定性的变化[28]。
1.4鸡TRAF6氨基酸多序列比对及进化保守位点分析利用Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)对TRAF6的氨基酸序列进行多序列比对,并使用Jalview软件分析TRAF6氨基酸位点的进化保守性。越是保守的位点发生变异,越可能影响蛋白质的功能[30]。
1.5鸡TRAF6基因与疾病表型相关nsSNPs预测
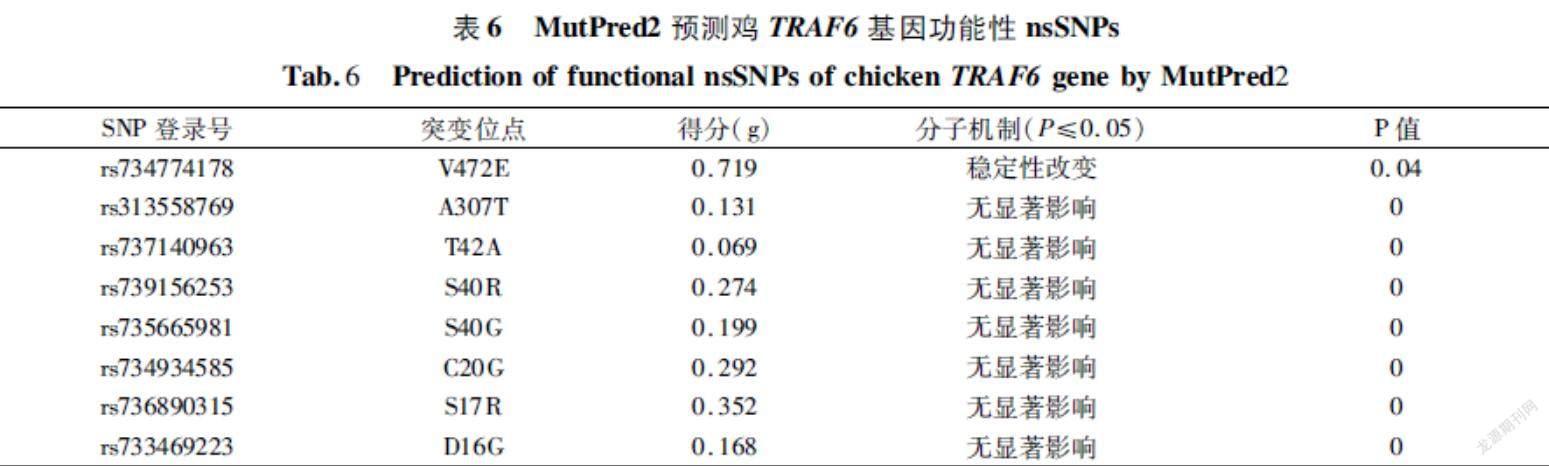
MutPred2(http://mutpred.mutdb.org/)[30]是一款可以用于预测突变与疾病相关性的在线工具。预测结果有两个重要分值总分数(g)和属性分数(P),其中g表示氨基酸替换是有害或疾病相关的概率,P则表示蛋白结构或功能是否有影响。结合g和P值,预测结果g>0.5且P<0.05的被称为中假设可信,g>0.75且P<0.05的被称为假设非常可信[30]。
1.6鸡TRAF6蛋白质二级和三级结构预测
利用Sopma(https://npsaprabi.ibcp.fr/cgibin/npsa_automat.pl?page=npsa_sopma.html)和SWISSMODEL(https://swissmodel.expasy.org/interactive)[30,31]在线软件对鸡TRAF6蛋白及其突变体的二级结构和三级结构分别进行预测,进而评估nsSNPs可能造成的影响。
2结果与分析
2.1鸡TRAF6基因nsSNPs的筛选
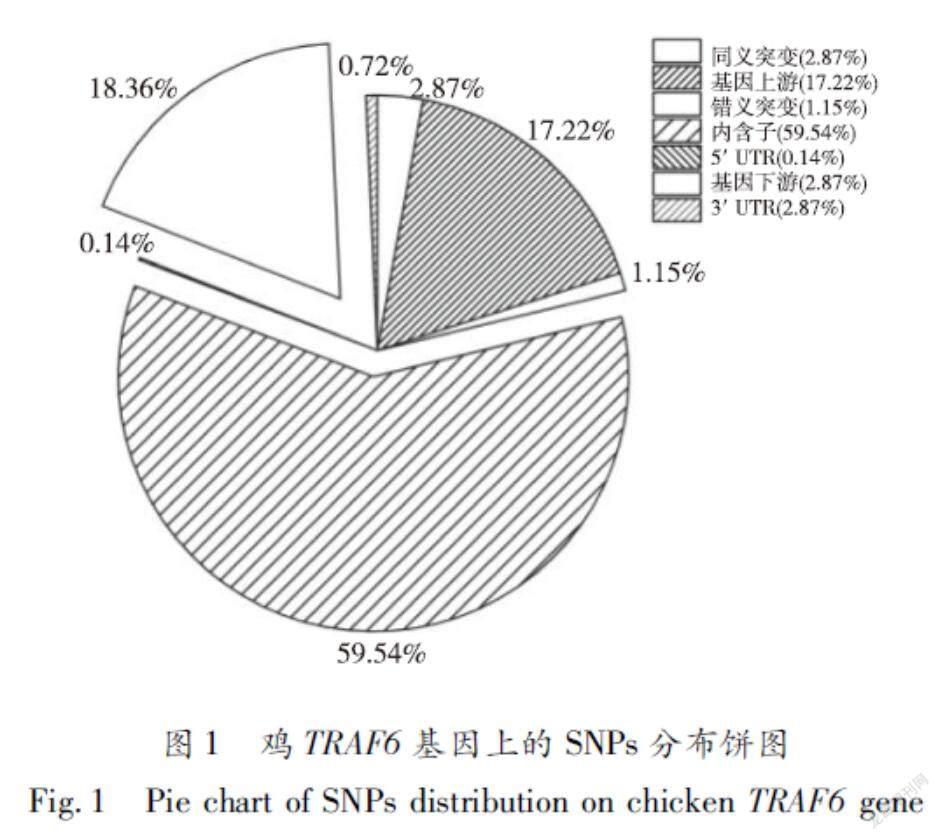
2.1.1鸡TRAF6基因上的SNPs分布
从Ensembl数据库中检索到TRAF6基因的转录本TRAF6203(ENSGALT00000105433),检索发现697个SNPs被标记在鸡TRAF6基因序列中,其中错义突变、同义突变、3′非翻译区(3′UTR)变异和5′非翻译区(5′UTR)变异分别有8个、20个、5个和1个;415(含2个剪接区变异)个变异位于内含子区,位于基因上游和下游的变异分别有120个和128个。SNPs数量百分比如图1所示。
2.1.2鸡TRAF6基因编码区nsSNPs的分布
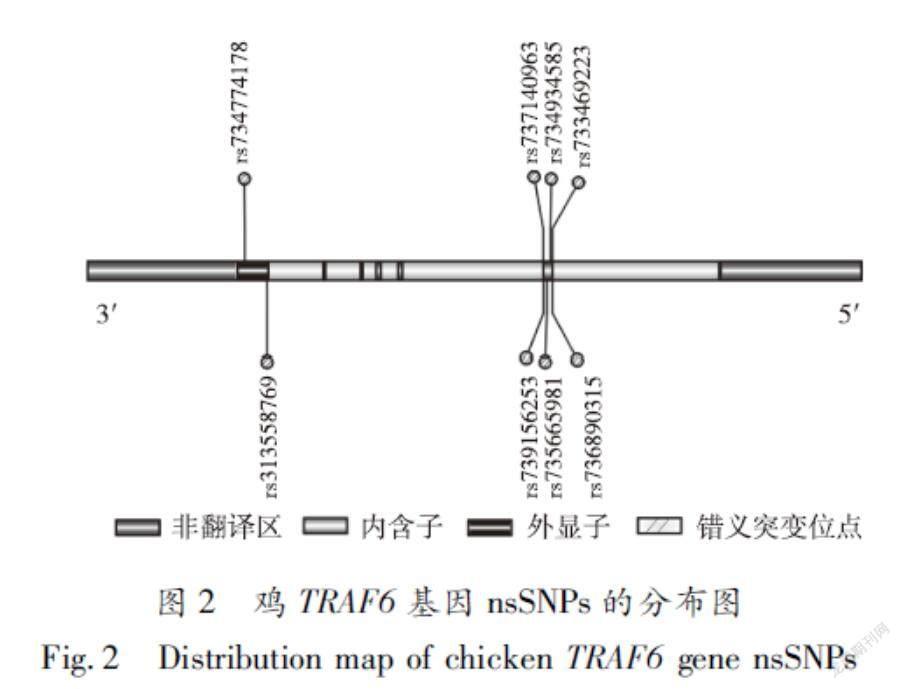
从dbSNP和Ensembl数据库中共筛选出8个(rs733469223、rs736890315、rs734934585、rs735665 981、rs739156253、rs737140963、rs313558769和rs734774178)nsSNPs位于鸡TRAF6基因的编码区。通过IBS软件绘制该8个nsSNPs的分布情况如图2所示。
2.2.1SIFT预测功能性nsSNPs
预测的得分表示该位点突变对蛋白质序列的影响。得分>0.05,可以耐受;得分≤0.05,有害。结果显示“有害”的nsSNPs有2个,其ID分别为rs734774178、rs736890315;可耐受的nsSNPs有6个,分别为rs313558769、rs737140963、rs7391562 53、rs735665981、rs734934585、rs733469223(表1)。得分越低,说明该nsSNP对蛋白质的功能影响越大。
2.2.2PROVEAN预测功能性nsSNPs
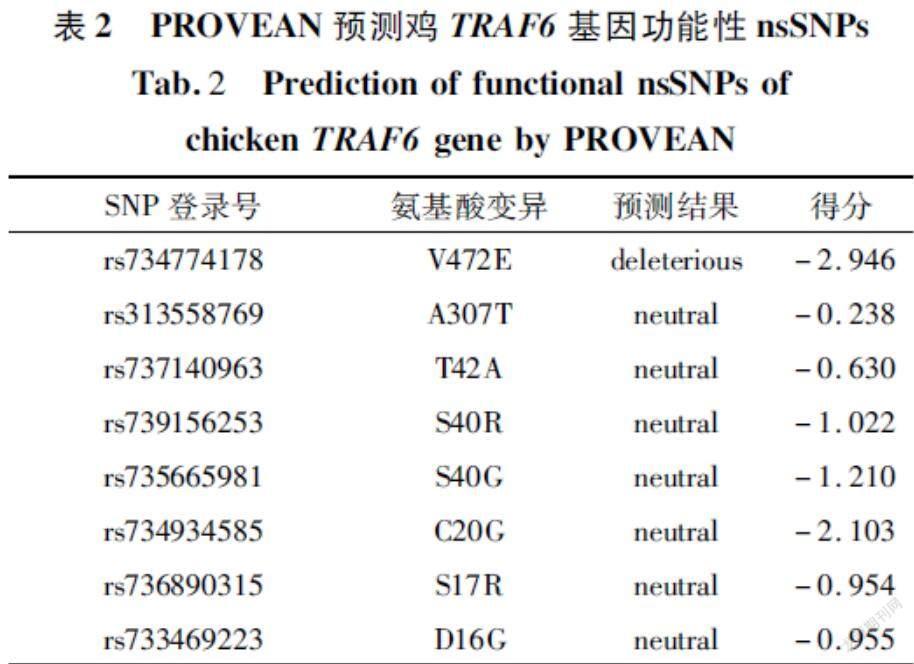
PROVEAN预测结果分为“有害”和“中性”两类。分数值≤-2.5预测为有害,分数值>-2.5预测为中性。在所分析的8个nsSNPs中,“有害”的nsSNPs有1个,其ID是rs734774178,其余的nsSNPs均为中性(表2)。
2.2.3PhDSNP预测功能性nsSNPs
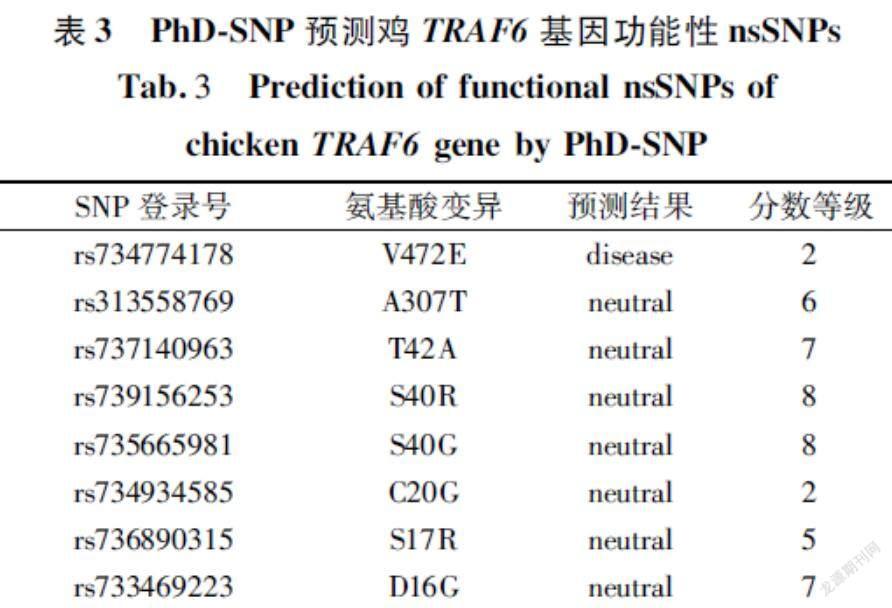
PhDSNP预测结果分为“疾病”和“中性”两类,可靠性指数(RI)在0~9之间。预测的结果中为疾病的nsSNPs有1个,其ID是rs734774178,表明该位点突变可能致病;其余的7个nsSNPs均为中性(表3)。
2.2.4SNAP2预测功能性nsSNPs
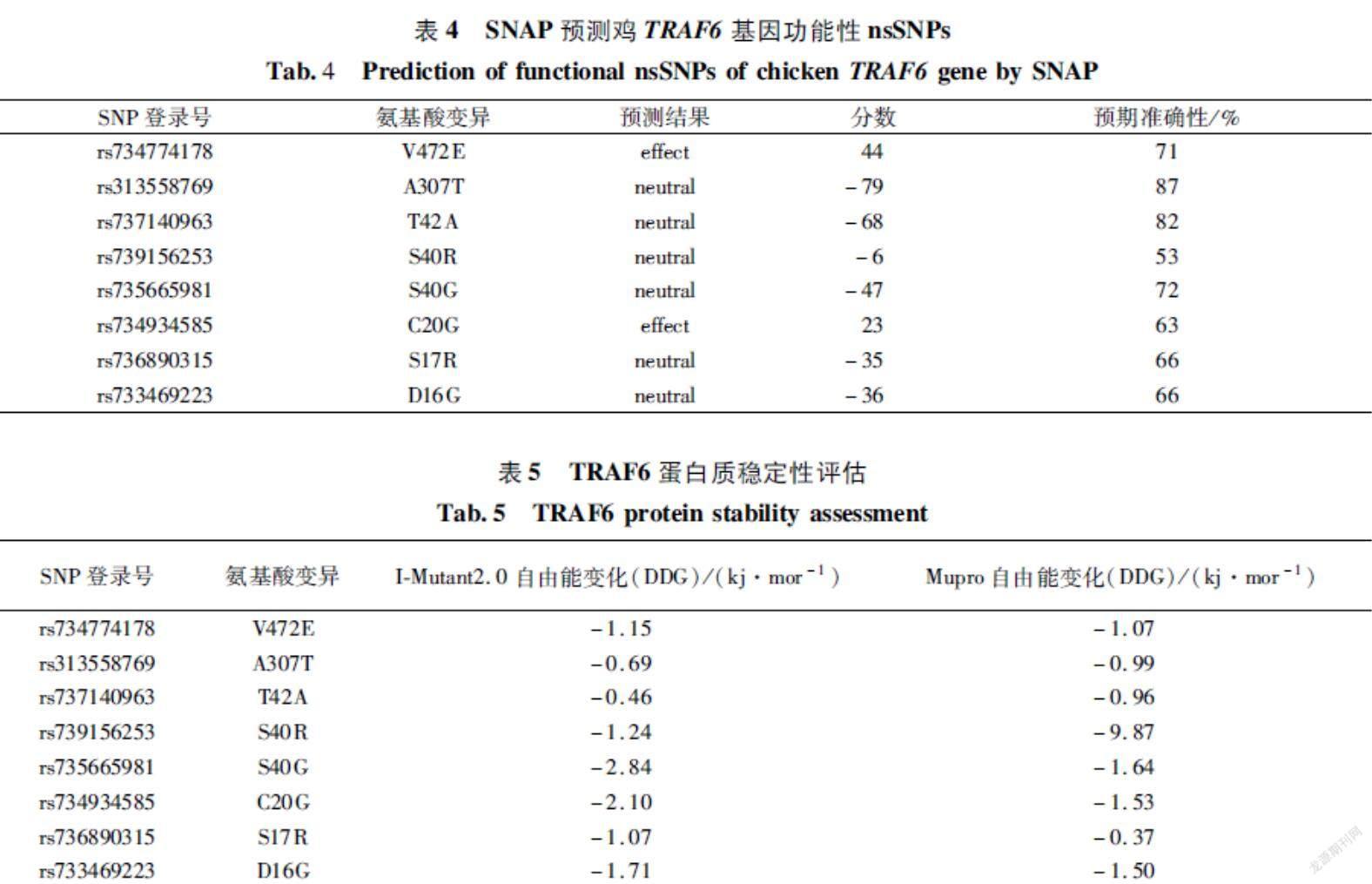
SNAP2的预测结果分为“有影响”和“中性”两类,输出分数值0~100预测为有影响,输出分数值-100~0预测为中性。预测的结果如表4所示,其中预测为有影响的有2个,其ID分别是rs734774178和rs734934585,预测为中性的有6个,分别是rs313558769、rs737140963、rs739156253、rs735665981、rs736890315、rs733469223。
2.2.5鸡TRAF6基因功能性nsSNPs预测结果
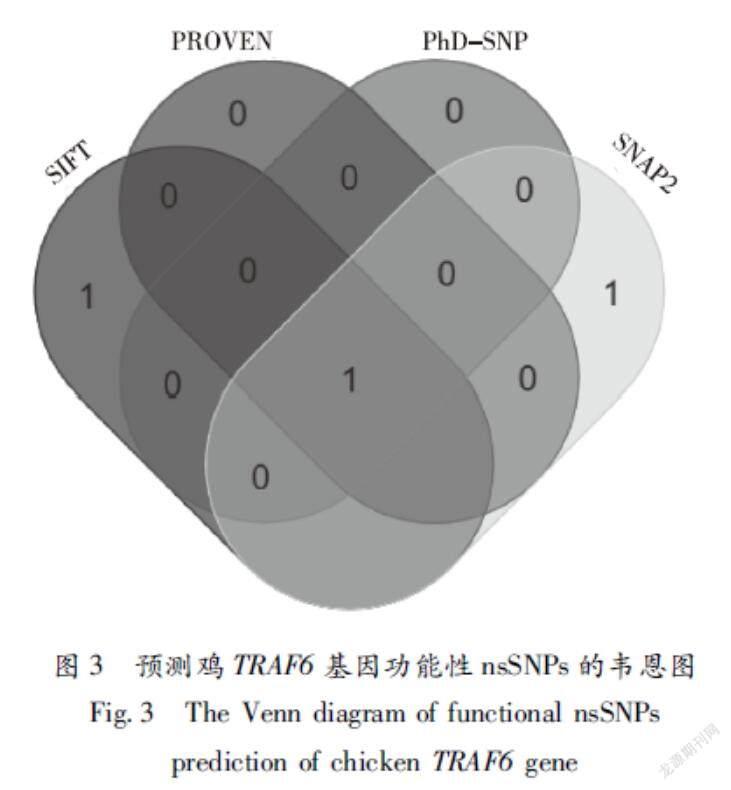
通过R语言的“Venn”package绘制SIFT、PROVEAN、PhDSNP和SNAP2四种预测方法预测的韦恩图(图3),四种软件功能性nsSNPs预测结果发现,分别预测到2个V472E、S17R、1个V472E、1个V472E和2个V472E、C20G nsSNPs位点,其中V472E有4种方法预测为有害位点。由此推测,V472E位点可能为鸡TRAF6基因功能性nsSNPs位点。
2.3鸡TRAF6蛋白质稳定性分析
大多数疾病相关的nsSNPs会影响蛋白质的稳定性。利用IMutant2.0和Mupro评估氨基酸替代对突变蛋白质稳定性的影响。自由能变化<0,表示此nsSNPs降低了蛋白质的稳定性,自由能变化>0,表示此nsSNPs增加了蛋白质的稳定性。结果如表5所示,所有突变位点均使TRAF6蛋白质稳定性下降。其中V472E为之前预测的功能性突变位点。
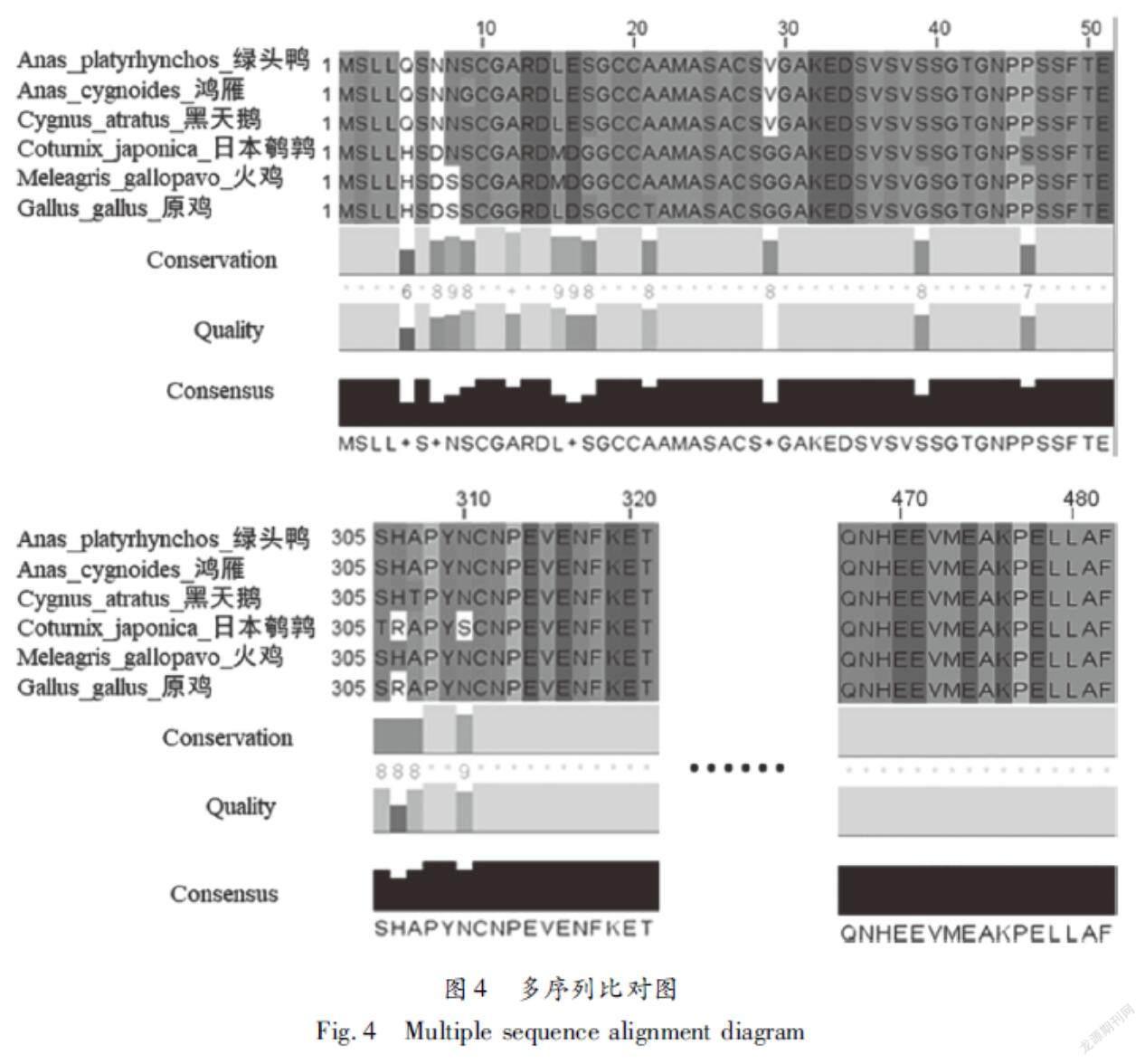
2.4鸡TRAF6蛋白的多序列比对和进化保守性位点预测在UniProt数据库中检索TRAF6蛋白在原鸡、火鸡、绿头鸭、鸿雁、黑天鹅和日本鹌鹑中的高度同源序列,通过Clustal Omega软件对TRAF6氨基酸序列进行多序列比对,再使用Jalview软件进行作图分析(如图4所示),在不同物种中不保守的nsSNPs有3个,分别是D16G、S17R和A307T;在不同物种中保守的nsSNPs有5个,分别是C20G、S40G、S40R、T42A和V472E。
2.5TRAF6突变位点可能的功能后果预测
MutPred2在线工具预测与疾病相关的表型。得分(g)>0.5且P<0.05,為中度假设可信;g>0.75且P<0.05,为假设非常可信。从表6可以看出,V472E位点的突变会导致蛋白质的稳定性改变,且V472E的功能后果预测为中度置信假设,其他位点的突变均无显著影响。
2.6鸡TRAF6突变体蛋白质二级结构预测
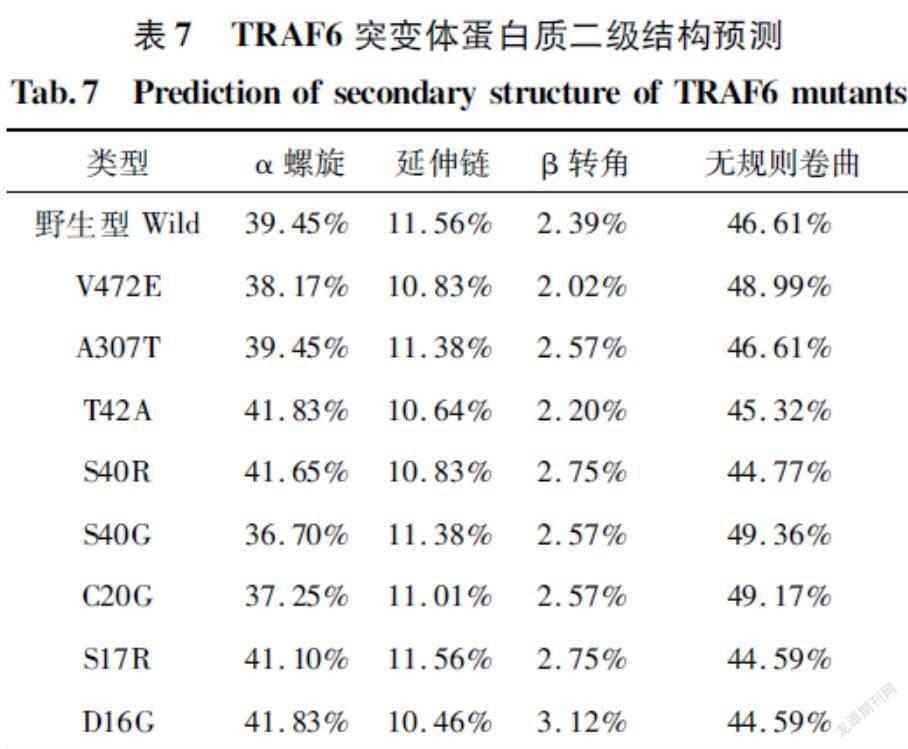
利用在线软件Sopma对鸡TRAF6蛋白的野生型和8种突变体进行二级结构预测,结果见表7。野生型TRAF6的蛋白质二级结构有,α螺旋(3945%)、延伸链(11.56%)、β转角(2.39%)、无规则卷曲(46.61%)4种结构。其中,C20G位点的突变均造成了TRAF6蛋白质二级结构的改变,导致了α螺旋百分比下降和无规则卷曲百分比的提高,而A307T位点的突变未能影响α螺旋和无规则卷曲的比例;T42A、S40R、S17R、和D16G位点的突变均导致了α螺旋百分比上升和无规则卷曲百分比的下降;除S17R突变未能影响延伸链的比例外,其他7处突变都导致延伸链百分比下降;V472E和T42A突变都导致β转角比例下降,其余6处突变均导致β转角比例上升。二级结构的变化通过影响蛋白的稳定性从而影响蛋白翻译调控过程,其稳定性通过最小自由能表示,最小自由能越低,结构稳定性越高。有研究表明,蛋白疏水性会随着α螺旋比例的降低而随着增大,随β转角和无规卷曲比例的升高而增大[32]。
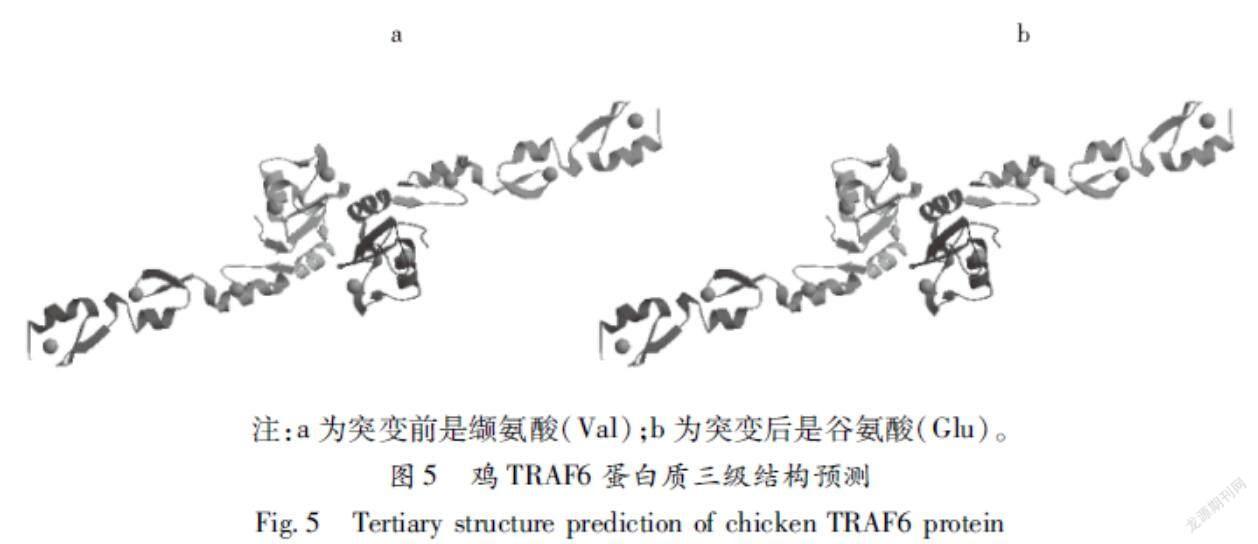
2.7鸡TRAF6蛋白质三级结构预测
使用SWISSMODEL在线软件对鸡TRAF6蛋白的野生型和突变体(V472E)的3D模型进行了预测分析。结果如图5所示,图中不同的颜色代表不同的二级结构,绿色、蓝色、红色和浅褐色分别代表α螺旋、无规卷曲、延伸链和β转角;由图5看出,TRAF6蛋白的野生型和突变体(V472E)的三级结构与二级结构预测结果一致,主要结构是α螺旋和无规卷曲。
3结论与讨论
nsSNPs是基因编码区可能对蛋白的结构和功能特性产生重大影响的单个氨基酸替换。近年来,随着生物测序技术的发展,对功能型nsSNPs预测,已越来越多地应用于人类医学和畜牧学功能型SNP的研究中。在人类和畜禽生命中,绝大部分nsSNPs会影响人和畜禽的健康,导致许多疾病发生。仅有少部分的nsSNPs不会改变蛋白质的结构和功能表达,不会影响人和畜禽的健康[33]。在家禽中,有研究者发现SNPs位点基因型对三穗鸭的蛋壳品质有影响,此外,SNP位点的突变可能影响动物的表现型[34]。研究报道[35],在人类大部分遗传病中基因变异是由SNP导致的[17]。例如,Song等[17]通过对人TRAF6 mRNA表达水平与TRAF6内含子SNP(rs4755453)基因型的关联分析发现,rs475545与脓毒症诱导人急性肺损伤的易感性显著相关,这可能是rs4755453通过影响TRAF6 mRNA的表达,来增加炎性细胞因子的产生,从而直接导致肺组织损伤。而少汗型外胚层发育不良(HED)疾病,其特征在于发育不良、汗湿不足。Fujikawa等[36]研究发现突变型TRAF6蛋白抑制了野生型TRAF6蛋白质与外胚层A相关受体死亡域蛋白(EDARADD)之间的相互作用,在外胚层衍生器官发育过程中潜在地抑制了EDARADD介导的NFκB的活性,从而导致HED表型。此外,对动物TRAF6基因的研究发现,TARF6是与生长发育、免疫调控相关的候选基因。研究报道,TRAF6自身泛素化的关键位点K132的突变抑制了NFκB的活性,随之Li等[37]研究发现,点带石斑鱼TRAF6的第132位氨基酸K突变为R,突变后的K132R通过激活NFκB下游信号通路,从而在宿主抵御寄生虫感染中起着关键作用。张芷毓等[38]在秦川牛TRAF6基因第六内含子17 628 bp处检测到的SNP位点形成两种基因型,该突变基因型对肉牛的育种具有显著影响。
本研究利用SIFT、PROVEAN、PhDSNPB和SNAP2四种生物信息学软件对鸡TRAF6基因外显子区功能性nsSNPs进行预测。这些方法既是相互独立又是相互补充的,因此,为减少预测结果差异,本研究结合四种预测软件进行预测,筛选出S17R、C20G、V472E这3个nsSNPs,其中V472E是四种软件共同筛选出的nsSNPs。研究发现,保守序列一般是生物体的生命所需,其突变常常致病。蛋白质的稳定性对生物学功能很重要,错义突变会导致蛋白质的稳定性降低,而IMutant2.0和Mupro是评估氨基酸替代对突变蛋白质稳定性的影响;MutPred2用于预测疾病表型。本研究通过对不同物种间的多序列比对和保守的多态性位点及其蛋白质的稳定性预测分析发现,V472E为保守的多态性位点且V472E位点的突变会导致蛋白质的稳定性改变,使蛋白质的稳定性下降。此外,MutPred2对氨基酸替代的功能预测与蛋白质二级结构预测结果相同,V472E位点的突变均造成了TRAF6蛋白质的稳定性改变。由此可知,V472E突变位可能严重影响鸡TRAF6蛋白质的结构,而蛋白质的结构改变会导致生物体的性状改变,故推测V472E突变位点可能是影响鸡生长发育和抗病育种的主要功能性SNPs。
本研究预测结果发现:rs734774178(V472E)nsSNPs使缬氨酸(Val)变成谷氨酸(Glu)。Val属于家禽功能性必需支链氨基酸之一,是非极性氨基酸,具有疏水性,在肌肉中氧化代谢[39]。研究发现,Val可以抑制免疫器官的发育及免疫球蛋白的合成,从而影响免疫功能,增加Val含量可以增强机体免疫力和抗氧化能力[4041]。王宇波等[42]发现,低含量Val饲粮增加了肥育猪肌内脂肪含量,从而影响肥育猪的生长性能。而Glu是一种带负电荷的极性氨基酸,具有亲水性,与多种疾病相关。一个氨基酸的疏水性和亲水性,极性与非极性都会影响氨基酸侧链基团功能的发挥及其各种酶和抗体、抗原之间的相互作用[43]。因此,本研究对V472E突变位点的二级结构预测发现与三级结構的功能预测相符。综上所述,本研究利用生物信息学软件预测筛选出致病的TRAF6基因的nsSNPs位点V472E,故推测V472突变位点可能影响鸡抗病表型的形成,但该位点的具体功能与致病表型之间的关系需要进一步研究。
鸡TRAF6基因的rs734774178(V472E)位点可能是影响鸡抗病育种的重要功能性突变位点,该位点突变可能严重影响鸡TRAF6蛋白质的结构从而发挥蛋白功能。
参考文献:
[1]Namjou B,Choi C B,Harley T W,et al.Evaluation of TRAF6 in a large multiancestral lupus cohort [J].Arthritis Rheum,2012,64(12):19601969.
[2]Stachon P,Missiou A,Walter C,et al.Tumor Necrosis Factor Receptor Associated Factor 6 Is Not Required for Atherogenesis in Mice and Does Not Associate with Atherosclerosis in Humans [J].PloS ONE,2009,5(7):e11589.
[3]Guo Y X,Xu Y,Kang X,et al.Molecular cloning and functional analysis of TRAF6 from Yangzhou great white goose Anser anser [J].Development and Comparative Immunology,2019,101(23):103435103446.
[4]王晓芬.TRAF6在家蝇免疫及生殖中的作用[D] .保定: 河北大学,2019.
[5]Jin J,Ran J S,Yang C W,et al.Molecular characterization,expression,and functional analysis of chicken TRAF6 [J].Genetics and Molecular Research:GMR,2017,16(1):28198500.
[6]Xiao L,Zhong M D,Huang Y,et al.Puerarin alleviates osteoporosis in the ovariectomyinduced mice by suppressing osteoclastogenesis via inhibition of TRAF6/ROSdependent MAPK/NFκB signaling pathways [J].Aging (Albany NY),2020,12(1):2170621729.
[7]Strickson S,Emmerich C H,Goh E T H,et al.Roles of the TRAF6 and Pellino E3 ligases in MyD88 and RANKL signaling [J].Proceedings of the National Academy of Sciences of the United States of America,2017,114:E3481E3489.
[8]陳子文.缺血性中风TRAF6基因多态性与mRNA、炎性细胞因子表达水平及MRI表现的相关性研究[D].南宁: 广西中医药大学,2017.
[9]Raychaudhuri S,Thomson B P,Remmers E F,et al.Genetic variants at CD28,PRDM1 and CD2/ CD58 are associated with rheumatoid arthritis risk [J].Nature Genetics,2009,41(1):13131318.
[10]Banram N J,Chan B C,Isaac T W,et al.Evaluation of TRAF6 in a large multiancestral lupus cohort [J].Arthritis & Rheumatism,2012,64(6):131139.
[11]Yang X R ,Pfeiffer R M,Wheeler W,et al.Identification of modifier genes for cutaneous malignant melanoma in melanomaprone families with and without CDKN2A mutations [J].International Journal of Cancer,2009,125: 29122927.
[12]Akiyama T,Maeda S,Yamane S,et al.Dependence of selftolerance on TRAF6directed development of thymic stroma [J].Science,2005,308(9):248251.
[13]Hassan M S,Shaalan A A,Dessouky M I,et al.A review study:Computational techniques for expecting the impact of nonsynonymous single nucleotide variants in human diseases [J].Gene,2019,680(8): 2033.
[14]张成岗,贺福初.生物信息学在新基因全长cDNA序列分析及功能预测中的应用 [J].生物化学与生物物理进展,2003,30 (1): 159162.
[15]Damell D K,Kaur S,Stanislaw S,et al.MicroRNA expression during chick embryo development [J].Developmental Dynamics,2007,236(1):315633165.
[16]Bromberg Y,Rost B.SNAP:predict effect of nonsynonymous polymorphisms on function [J].Nucleic Acids Research,2007,35 (11):38233835.
[17]Song Z,Yao C,Yin J,et al.Genetic variation in the TNF receptorassociated factor 6 gene is associated with susceptibility to sepsisinduced acute lung injury [J].Journal of Translational Madicine,2012,10(1):160166.
[18]Weng R X,Chen W,Tang J N,et al.Targeting spinal TRAF6 expression attenuates chronic visceral pain in adult rats with neonatal colonic inflammation [J].Molecular Pain,2020,16(1):17441754.
[19]李永濤,李红杰,王晓雪,等.猪肿瘤坏死因子受体相关因子I的克隆,表达及对伪狂犬病病毒的抑制效 [J].畜牧兽医学报,2017,48 (2): 289296.
[20]高玮玮,潘宝平,闫春财,等.青蛤 (Cyclina sinensis) TRAF6基因克隆及其在PolyI: C胁迫下的免疫应答[J].海洋与湖沼,2016,47 (5): 10401046.
[21]Phelan P E,Mellon M T,Kimim C H.Functional characterization of fulllength TLR3,IRAK4,and TRAF6 in zebrafish (Danio rerio) [J].Molecular Immunology,2005,42(2):10571071.
[22]Hu Q ,Song J,Ding B,et al.miR146a promotes cervical cancer cell viability via targeting IRAK1 and TRAF6 [J].Oncology Reports,2018,399(1):30153024.
[23]Sherry S T,Ward M H,Kholodov M,et al.The NCBI database of genetic variation [J].Nucleic Acids Research,2001,29 (1):308311.
[24]Sim N L,Kumar P,Hu J,et al.Sift web server:Predicting effects of amino acid substitutions on proteins [J].Nucleic Acids Research,2012,40 (Web Server issue):452457.
[25]Choi Y W,Chan A P.PROVEAN web server: a tool to predict the functional effect of amino acid substitutions and indels [J].Bioinformatics,2015,31(2):27452757.
[26]Capriotti E,Calabrese R.Casadio R.Predicting the insurgence of human genetic diseases associated to single point protein mutations with support vector machines and evolutionary information[J].Bioinformatics,2006,22(22):27292734.
[27]Hecht M,Bromberg Y,Rost B.Better prediction of functional effects for sequence variants [J].BMC Genomics,2015,8:S1.
[28]Capriotti E,Fariselli P,Casadio R.IMutant2.0: predicting stability changes upon mutation from the protein sequence or structure [J].Nucleic Acids Research,2005,33(21):306310.
[29]Singh S K,Reddy M S.Computational prediction of the effects of nonsynonymous single nucleotide polymorphisms on the GPIanchor transamidase subunit GPI8p of Plasmodium falciparum [J].Computational Biology and Chemistry,2021,92(12):10741080.
[30]李玉冬,王伟佳,李紫薇,等.鸡胰岛素样生长因子2基因(IGF2)外显子区功能性SNP预测与分析 [J].畜牧兽医学报,2020,51(11): 26652678.
[31]Nathan V K,Lizzy M,Edward G W.Superoxide Dismutase and Prediction of Mutational Variations through in silico Methods [J].International Journal Bioautomation,2014,18(2):2329.
[32]刘春雷,孙立斌,李相昕,等.大豆7S和11S蛋白二级结构与表面疏水性相关性的研究 [J].食品科学,2015,36(17): 2832.
[33]Hassan M S,Shaalan A A,Dessouly M I,et al.A review study:Computational techniques for expecting the impact of nonsynonymous single nucleotide variants in human diseases [J].Gene,2019,680(2):2033.
[34]李杰章,吴磊,谭光辉,等.三穗鸭蛋壳品质关联基因CALM1突变位点SNPs的鉴定与分析 [J].山地农业生物学报,2020,39(1): 6772.
[35]Ng P C,Henikff S.Predicting the effects of amino acid substitutions on protein function [J].Annual Review of Genomics and Human Genetics,2006,7(1):6180.
[36]Fujikawa H,Farooq M,Fujimoto A,et al.Functional studies for the TRAF6 mutation associated with hypohidrotic ectodermal dysplasia [J].British Journal of Dermatology,2013,168(1):62933.
[37]Li Y W,Li X,Xiao X X,et al.Molecular characterization and functional analysis of TRAF6 in orangespotted grouper (Epinephelus coioides) [J].Development and Comparative Immunology,2014,44(4):217225.
[38]张芷毓,欧德琼,邬明丽,等.秦川牛TRAF6基因多态及其与体尺性状的关联分析 [J].中国牛業科学,2018,44(4): 48.
[39]Mack S,Bercovici D,De G G,et al.Ideal amino acid profile and dietary lysine specification for broiler chickens of 20 to 40 days of age [J].British Poultry Science,1999,40(11):25765.
[40]徐琪寿.氨基酸药理学研究进展[J].氨基酸和生物资源,1996,18 (1): 3032.
[41]邢恒涛,颜桂花,刘强,等.可溶性玉米纤维(Fibersol2)对仔猪生长性能、血清生化指标和抗氧化能力的影响 [J].畜牧兽医学报,2018,49(10): 21702179.
[42]王宇波,许豆豆,何鑫,等.低蛋白饲粮缬氨酸水平对肥育猪生长性能、胴体性状和肉品质的影响 [J].畜牧兽医学报,2019,50(9): 18321840.
[43]Islam M M,Kobayashi K,Kidokoro S I,et al.Hydrophobic surface residues can stabilize a protein through improved waterprotein interactions [J].FEBS Journal,2019,2869(21):41224134.
