陆地棉种质资源株型相关性状鉴定与分析评价
2022-02-15马麒徐守振宁新柱陈红谢斌李吉莲林海
马麒,徐守振,宁新柱,陈红,谢斌,李吉莲,林海
(新疆农垦科学院棉花研究所/农业农村部西北内陆区棉花生物学与遗传育种重点实验室,新疆 石河子 832000)
陆地棉(Gossypium hirsutumL.)是世界上最具经济价值的纺织作物之一。长期以来增加纤维产量对现代棉花生产者和育种家来说一直是一个重大挑战[1],与增加单株产量潜力相比,通过增加种植密度对提高棉花单位面积产量的潜力更大,而种植密度主要由单个植株的独特株型结构决定[2]。因此,株型结构对于当前棉花生产具有重要意义。棉花株型结构是根据果枝和叶枝的分布情况以及果枝的长短而形成的综合性状,在众多的株型相关性状中,株高、果枝夹角和果枝长度是决定株型结构最关键的三个因素,决定着一个棉花品种能否适宜机采及轻简化栽培[4,5]。当前普遍认为棉花机采需要多个株型性状相互协调,一般要求始果节高度>20cm,果枝夹角相对较小,株高为75~85cm为宜,株型育种已成为棉花遗传育种的一个重要方向。因此,对株型相关性状进行精准鉴定与评价可为株型育种提供重要理论依据和参考信息。李成奇等[6]对陆地棉百棉1号的主要株型性状进行表型鉴定及遗传研究;杨万玉等[7]通过对棉花株型相关性状进行鉴定,并开展株型与产量的关系研究,发现各果节间长度与皮棉产量呈开口向下的二次抛物线关系,果枝的粗度与皮棉产量呈显著正相关,适宜的果节间长度和较粗的果枝是获得棉花高产的结构基础。Wen等[8]对121份棉花材料的株高和果枝分枝数两个性状进行三个环境下的表性鉴定与分析,发现两个性状在三个环境下存在较为广泛的变异,且两者相关系数为0.37,存在极显著(P<0.01)的相关性。迄今为止,人们对棉花株型相关性状的系统鉴定与评价的研究还相对较少,因此在进行株型结构的改良育种时,一方面存在着种质资源匮乏的问题,而另一方面,真正可供选择参考的理论依据也不多,这严重阻碍了当前棉花株型育种的进程。研究对象,对三个株型相关性状进行田间精准鉴定,开展表型变异度、相关性、方差和遗传力,以及区域差异性分析及聚类分析,最终对315份材料的株型相关性状进行综合分析与评价,以期为机采棉株型育种提供重要的理论依据和参考信息。
1 材料与方法
1.1 试验材料
本研究以315份陆地棉种质资源组成的自然群体为试验材料,这些材料主要来源于不同的国家或地区,其中包括前苏联(Former Soviet Union,FSU)材料4份,美国(United States of America,USA)材料21份,中国自育材料290份。自育材料中有125份来自于黄河流域(Yellow River region,YRR),48份来自于长江流域(Yangtze River region,YZRR),20份来自于北方特早熟区(Northern special early maturing region,NSER),97份来自西北内陆区(Inland Northwest region,NIR)。
1.2 田间试验
田间试验分别于2014年和2015年在河南省安阳(36.13'N,114.80'E)和新疆石河子(44.52'N,86.02'E)四个不同环境下开展。在每个环境中,我们将所有的材料按照三重复随机区组设计,在河南省安阳试验点,所有陆地棉材料都种植在一个0.8m×4.0m的试验小区,每个小区内种有20株;在新疆石河子试验点,小区大小为1.5m×2.0m,每个小区内种有40株。田间栽培和管理措施完全参考当地大田的耕作栽培方法。
1.3 表型性状鉴定方法
在不同的环境条件下,对自然群体的三个与株型相关性状株高(PH)、果枝夹角(FBA)、果枝长度(FBL)进行测定。株高的测定是从植株顶部到子叶节的高度,果枝长度的测定是从果枝(着生主茎部位)至果枝顶端的长度,果枝夹角的测定是植株主茎与果枝的夹角,果枝长度和果枝夹角的测量取植株中部果枝(6~8台果枝)测定。具体测定调查方法参照SU[9]等人研究中的详细描述。
1.4 数据分析
利用Excel进行表型数据的统计与分析;利用IBM SPSS 26.0软件进行基本统计分析,包括差异显著性分析、广义遗传力(Broad-sense heritability,hB2)的计算,以及表型方差显著性分析等;利用Origin9.0和R语言软件进行不同区域株型相关性状的差异图和聚类图的绘制。
2 结果与分析
2.1 陆地棉株型相关性状的表型变异分析
在4个不同的田间环境(E1~E4)下,对自然群体的3个株型相关性状,包括株高、果枝夹角和果枝长度进行鉴定和变异度评价。由表1可以看出,315份陆地棉种质的三个株型相关性状存在不同程度的变异性,其中PH的变异范围较广,在E1~E4四个环境下PH的平均值为(63.26±8.79)cm,表型变异范 围 在36.09~88.76cm之 间;FBA的 平 均 值 为(61.84±3.85)°,变异范围在48.72°~72.50°之间;FBL的平均值为(12.60±2.90)cm,变异范围为5.16~24.99cm。在四个不同的环境下,PH、FBA和FBL三个性状的变异系数分别在11.02%~15.92%、4.67%~7.51%和17.42%~28.54%之间,其中PH的平均变异系数为13.90%,在四个环境下的变化相对较为一致,FBA的平均变异系数为6.22%,FBL的平均变异系数为23.01%。上述结果说明,在陆地棉的三个株型相关性状中FBL的变异度最大,PH变异度居中,而FBA的变异度最小。

表1 陆地棉株型相关性状的表型变异分析Table 1 Phenotypic variation of plant architecture related traits
2.2 陆地棉株型性状的相关性分析
表2相关性分析表明,陆地棉三个株型相关性状之间存在着极显著(P<0.01)相关性。PH分别与FBA、FBL极显著(P<0.01)相关,相关系数分别为0.236和0.324,FBA与FBL之间也存在极显著(P<0.01)的相关性,相关系数为0.461。上述结果表明,FBL与FBA的相关性大于FBL与PH的相关性,FBA与FBL的相关性大于FBA与PH的相关性。这说明在棉花生长过程中通过调控株高同步控制果枝长度,而果枝夹角次之;但果枝长度与果枝夹角同步调控的可能性大于果枝长度与株高,而果枝夹角与果枝长度同步调控的可能性大于果枝夹角与株高。

表2 陆地棉株型相关性状的相关性分析Table 2 The correlation analysis among three plant architecture-related traits
2.3 陆地棉株型相关性状的方差及遗传力分析
表3方差分析结果表明,陆地棉三个株型相关性状在基因、环境和基因×环境互作中存在显著(P<0.01)差异性。通过计算三个性状在四个环境下的广义遗传力(hB2)可知,PH、FBA和FBL的hB2分别为72.51%、25.12%和53.40%,这一结果表明PH和FBL的广义遗传力相对较大,可以相对稳定地遗传给后代,而FBA的广义遗传力相对较小,难以稳定地遗传,且易受田间环境因素(年份和地点)的影响。
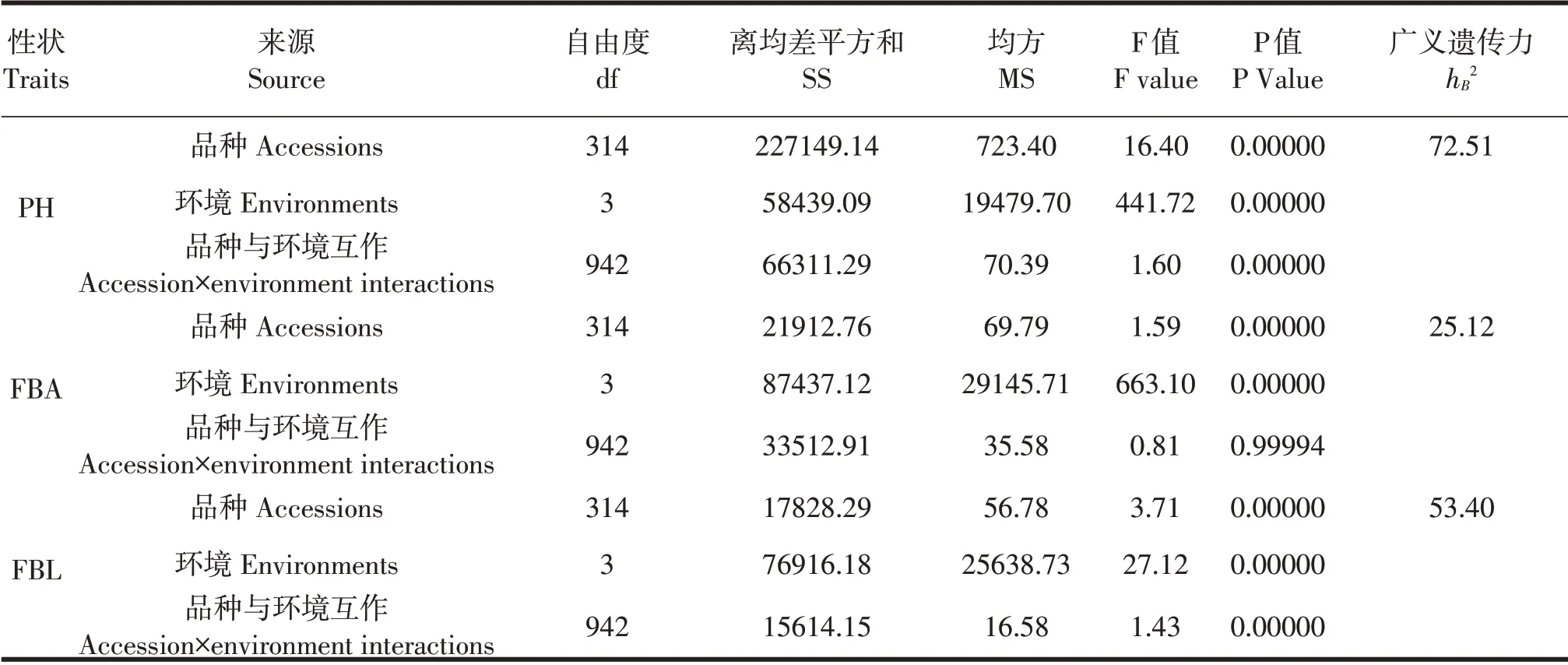
表3 陆地棉三个株型相关性状的方差分析Table 3 Analysis of variance of three plant architecture-related traits in four planting environments
2.4 陆地棉种质资源基于株型性状的聚类分析
聚类分析表明,315份陆地棉种质资源具有明显的类群关系。由图1A可知,当欧氏距离为30时,315份陆地棉种质资源被聚为Ⅰ(66份材料)、Ⅱ(25份材料)、Ⅲ(224份材料)三个大类群。由图1B可看出,当欧氏距离为15时,Ⅰ类群被划分为A(1份材料)和B(65份材料)两个亚群,Ⅱ类群被划分为C(18份材料)和D(7份材料)两个亚群;Ⅲ类群进一步划分为E(92份材料)和F(132份材料)两个亚群(图1B)。
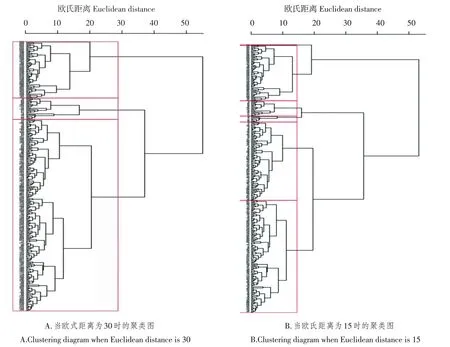
图1 陆地棉种质资源基于株型性状的聚类图Figure 1 Cluster diagram of upland cotton germplasm based on plant architecture related traits
为了明确三大类群陆地棉种质株型相关性状的差异性,对三个类群材料的株型相关性状进行类群之间的差异显著性分析,见表4。结果发现,类群Ⅰ、类群Ⅱ和类群Ⅲ材料的平均PH分别为(73.19±9.00)cm、(47.99±8.07)cm和(62.08±9.45)cm,三个类群之间的差异达到极显著水平(P<0.01)。类群Ⅰ、类群Ⅱ和类群Ⅲ材料的平均FBA分别为(62.34±8.39)、(59.50±8.09)和(61.95±8.21),且类群Ⅰ与类群Ⅲ之间的差异性不显著(P>0.05),但类群Ⅱ与类群类群Ⅰ、类群Ⅲ之间的差异均达到极显著水平(P<0.01)。类群Ⅰ、类群Ⅱ和类群Ⅲ材料的平均FBL分别为(13.34±6.21)cm、(9.30±4.69)cm和(12.72±6.11)cm,三个类群之间的差异达到显著水平(P<0.05),而且类群Ⅱ与类群类群Ⅰ、类群Ⅲ之间的差异均达到极显著水平(P<0.01)。变异度分析表明,PH在类群Ⅰ、Ⅱ和Ⅲ的变异系数分别为12.31%、16.82%和15.22%,FBA在类群Ⅰ、Ⅱ和Ⅲ的变异系数分别为13.46%、13.09%和13.26%,FBL在类群Ⅰ、Ⅱ和Ⅲ的变异系数分别为46.15%、50.38%和48.07%,整体从性状来讲,FBL的变异度较大,从聚类群来说,类群Ⅱ三个株型性状的变异度最大。

表4 陆地棉三个类群的株型相关性状的差异性分析Table 4 Significance analysis of plant architecture related traitss of upland cotton among three groups
2.5 陆地棉株型性状的区域聚类及差异性分析
不同区域来源的陆地棉种质聚类分析发现(图2),当欧氏距离为20时,来自USA的材料被单独划分为一个类群,其余区域的材料被划分为一个类群;当欧氏距离为15时,来自USA和NIR的材料分别聚为一个单独类群,来自YZR、YRR、FSU和NSER区域的材料合计被聚为一个类群。上述结果表明,根据株型性状来划分的话,来自美国的棉花材料其遗传背景较为独特,并且与西北内陆棉区的材料遗传背景有较大差异,亲缘关系较远;而西内陆棉区材料与长江流域、黄河流域、前苏联及北方特早熟棉区的遗传背景差异较大,亲缘关系较远。这说明在今后我国棉花的株型改良育种中,首先要重点利用美国材料做亲本,以改良本土品种的株型结构,西北内陆棉区棉花在株型育种中要重点关注长江流域、黄河流域、前苏联及北方特早熟棉区的材料并加以利用。

图2 陆地棉种质基于株型性状的区域聚类图Figure 2 Regional clustering diagram of upland cotton germplasm based on plant architecture related traits
对陆地棉种质的株型性状进行不同区域间的差异性分析发现,PH在六个不同环境材料间呈现出显著性(P<0.05)差异,且NIR环境的材料的PH大于其他五个环境的材料,差异达到了极显著性水平(P<0.01),来自USA和YRR的材料的PH大于NSER的材料,差异也达到了极显著水平(P<0.01);FBA在六个环境的表现是:USA>NSER>YZR>NIR>FSU>YRR,但差异未达到显著水平;FBL在六个环境的表现是:USA>NSER>YZR>NIR>YRR>FSU,差异同样未达到显著水平(见表5,图3)。

表5 陆地棉不同区域材料的株型相关性状的差异性分析Table 5 Significance analysis of plant architecture related traitss of upland cotton from different regions

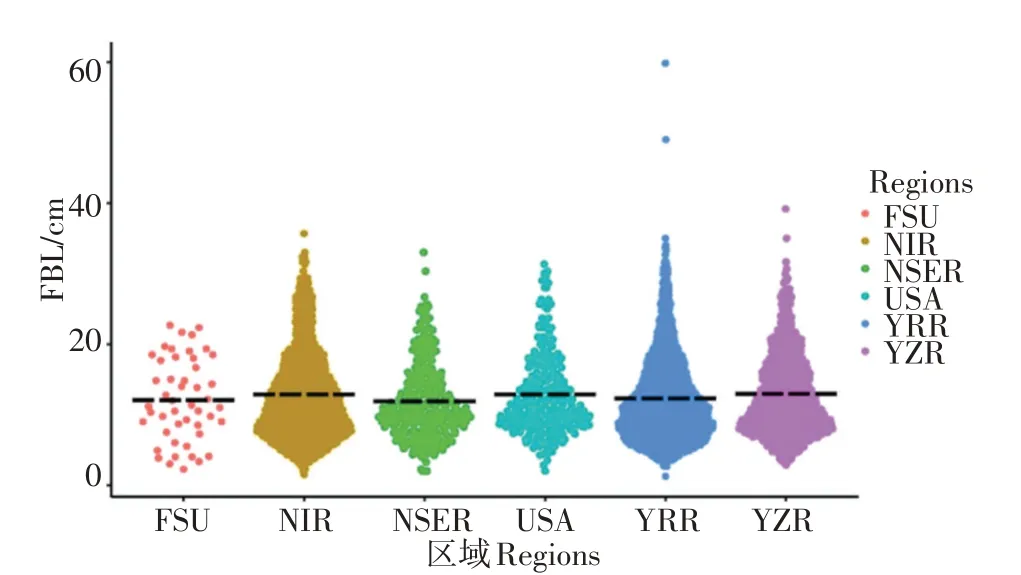
图3 陆地棉种质株型性状的区域差异性比较图Figure 3 Comparison of regional differences in plant architecture related traits of upland cotton germplasm
3 讨论与结论
3.1 丰富的种质资源是棉花遗传育种的基础
种质资源是棉花遗传育种的基础[10]。本研究中所利用的315份陆地棉种质是收集的不同区域来源的陆地棉品种/系,既有前苏联、美国等国外来源的材料又有我国四大棉区,即西北内陆棉区、长江流域棉区、黄河流域棉区以及北方特早熟棉区的材料,这些材料具有广泛的株型变异、丰富的遗传背景,为研究陆地棉株型相关性状提供了材料基础。在田间目标性状鉴定时,试验又设置了多点多环境三重复的种植与数据采集,为株型相关性状的精准鉴定提供了重要保障。
3.2 株型相关性状的鉴定与分析评价是棉花株型遗传育种的关键
表型性状的鉴定分析与评价是作物种质资源高效利用的基础[11]。本研究中发现,在陆地棉的三个株型相关性状中果枝长度的变异度最大,而果枝夹角的变异度最小,表明果枝长度性状的以均数为准变异程度大,离散趋势强;相反,果枝夹角性状则以均数为准变异程度小,离散趋势弱,这说明果枝长度性状中更容易挖掘具有极端差异性状的材料。通过计算三个性状的广义遗传力得知,株高和果枝长度的广义遗传力相对较大,而果枝夹角的广义遗传力相对较小,这说明株高和果枝长度性状可以相对稳定地遗传给后代,相反果枝夹角则难以稳定地遗传,且易受田间环境因素的影响,这一研究结果与SU等[9]对陆地棉株型性状的研究结果基本一致。相关性分析发现,三个株型相关性状之间存着在极显著的相关性,这说明在棉花生产中控制棉花株高时很容易引起果枝长度和果枝夹角的同步调控,因此可借助于化学调控的方法实现株高、果枝夹角和果枝长度性状的变化来同步塑造棉花株型结构的目的。
3.3 不同区域材料的聚类及差异性分析为棉花株型遗传育种提供重要参考依据
对不同区域材料基于株型性状进行聚类分析发现,当欧氏距离为20时,来自美国的材料被单独从315份材料中划分出来而作为一个单独类群,其余区域的材料被划分为一个类群;但当欧氏距离为15时,来自美国和西北内陆棉区的材料分别各自聚为一个单独类群,而来自我国长江流域、黄河流域、北方特早熟棉区以及前苏联的材料被聚为一个类群,这说明根据株型结构来划分的话,美国棉花遗传背景较为独特,相较与西北内陆棉区的材料其遗传背景差异较大,亲缘关系较远,这可能由于美国较好的光热条件和相对松散稀疏的栽培模式,造成在长期的人工选择驯化中材料的株型结构较为高大松散。西内陆棉区材料与长江流域、黄河流域、前苏联及北方特早熟棉区的遗传背景差异较大,亲缘关系较远,这说明西北内陆棉区长期以来采用“矮”“密”“早”“膜”的栽培技术导致材料的株型结构更加趋向于紧凑型,与我国其他几大棉区的材料在株型结构方面发生了根本性的差异。通过对不同区域材料的株型相关性状进行差异性分析时发现,西北内陆棉区的材料株高大于其他五个区域的材料,且差异达到了极显著性水平(P<0.01);而相比其他区域的材料,美国材料的果枝夹角相对较大,果枝相对较长,但未达到差异显著性水平。上述研究结果说明,今后我国棉花的株型改良育种过程中首先要重点利用美国材料做亲本,以改良本土品种的株型结构;而在本土种质资源的利用中要综合考虑六个棉区材料的株型结构,相互引进,相互开发利用,有效促进我国棉花株型育种取得重要进展,选育出高产、适宜轻简化栽培和机械采收的棉花新品种。综上所述,对棉花表型性状的鉴定分析与评价可有效实现棉花种质资源高效利用,对促进棉花株型遗传育种具有重要意义。
