舞草侧生小叶旋转的生态意义感性运动还是防御行为?
2022-02-15樊玥妮董梓豪邓子菁
樊玥妮,董梓豪,邓子菁,巩 岩,2*
(1.山西师范大学,山西太原030092;2.山西师范大学现代文理学院,山西临汾041004)
跳舞草学名舞草或电报植物(Codariocalyxmo⁃torius),为豆科蝶形花亚科舞草属植物,直立小灌木,三出复叶,顶生叶较大(3~7 cm),0~2个侧生小叶(1~2 cm),植株可供观赏及药用[1]。舞草原产亚洲和北澳洲[2],广泛分布于整个南亚及东亚和东南亚的部分地区,在我国主要分布在福建、江西、广东、四川、贵州、云南、台湾等地区[3]。

图1 舞草的(A)顶生大叶;(B)侧生小叶;(C)花型;(D)主要分布范围(图片整理自印度生物多样性开源门户网站:https://indiabiodiversity.org/)Fig.1(A)Large terminal leaves,(B)lateral leaflets,(C)flower typeand(D)main distribution range of C.motorius.(image from India Biodiversity Portal)
1881年,达尔文详细描述了舞草复叶闭合运动具有类含羞草的昼夜节律性(Circadian rhythm)[4]。而次昼夜节律(Ultradian rhythm)指周期短于24 h的节律现象,常用于医学或神经科学[5]。舞草复叶的侧生小叶在一定的温度和光照下,可垂直于复叶叶柄的方向做出近180°的摆动(上下张合)或360°旋转(“自由泳”或“蝶泳”),展现出肉眼可见的次昼夜节律运动,周期约2~5 min(见补充材料1)。舞草侧生小叶基部的叶枕[6]是其主要运动“器官”,环状对称性及围绕中央输导组织发达的晶鞘纤维(富含草酸钙结晶)是其重要的解剖学特征[7-10]。科研人员已从电生理学[8,11-13]和细胞生物学[14-15]等角度对舞草侧生小叶运动的内在机制进行了深入研究,对影响其运动节律的多种因素进行了细致描述(物理负重[16-17]、化 学 试 剂[7-8,18]、电 流[13,9-20]、无 线 电[21]、磁场[22]、光照[23-24]、温度[25]等),但对侧生小叶旋转运动的生态意义了解较少[26]。
Bünning最早认为舞草侧生小叶运动可能会精确感知明暗交错的时数,并在花形态建成过程中提供了最佳的重力条件[27],但相关研究缺乏数据支撑。Johnsson认为舞草侧生小叶运动在促进气孔水分运输和(或)优化光子捕获方面有积极作用[28]。而Lev-Yadun则认为该运动具有防御性质,可能为模拟某些植食性昆虫——特别是具翅节肢动物,以驱使其同类离开或吸引其天敌[29],降低被采食风险。通过对舞草相关研究进行整合,结合现有的理论知识,对舞草侧生小叶次昼夜节律行为的生态意义进行科学推测,以期为全面了解舞草侧生小叶运动的生态意义提供新思路。
1 舞草侧生小叶运动相关研究进展
1.1 舞草侧生小叶运动的结构基础
科学家们对舞草顶生叶叶枕的解剖学研究的开展较早[8-10],其结构与其他具有昼夜节律性运动的豆科植物类似[30]。位于舞草侧生小叶底部的叶枕直径约0.4 mm,长约3 mm。侧生小叶叶枕由中央核心(髓、导管和厚壁组织)向外依次分布着:厚角组织(2~3层细胞)、薄壁细胞(运动细胞,直径约10µm,长约15µm)和表皮(图2A左)。

图2 (A)叶枕组织切片显微图像;(B)叶枕细胞排列及运动模式示意图[31]Fig.2(A)Microscopic images of occipital tissue sections;(B)Schematic diagram of occipital cell arrangement and move‑ment mode
侧生小叶叶枕的环状对称性及围绕中央输导组织发达的晶鞘纤维(富含草酸钙簇晶)是重要的解剖学特征(图2A右)[7]。同时,侧生小叶叶枕细胞的细胞壁微纤丝排列成垂直于径向的环状结构(图2B),限制了运动细胞径向扩增,但允许其纵向延伸[32];其协调可逆的体积变化形成侧生小叶的椭圆运动[9,31]。舞草侧生小叶叶枕结构与顶生叶叶枕组织不对称具有明显差异[33],在淀粉粒有无、管胞分布、运动细胞大小等方面具有明显区别(见表1)。

表1 舞草复叶中顶生叶叶枕与侧生小叶叶枕的特征比较[7,31,34]Table 1 Comparison of the characteristics of pulvinus between terminal leaf and lateral leaflet of C.motorius
1.2 舞草侧生小叶运动模式
侧生小叶叶枕运动机制方面研究开展的较早[35-37],其运动细胞膨胀和收缩的生理学机制与气孔保卫细胞有许多相似之处[38],但运动模式却显著不同。处于图2B所示状态时,叶枕呈向上弯曲,顶部运动细胞呈收缩状态,底部为膨胀状态;经过半个周期,上部运动细胞膨胀,下部收缩,弯曲方向发生逆转。相邻的运动细胞或依次发生连续变化,或分区域发生变化,展现出的运动模式即为“上下张合”或“旋转”(补充材料1)。在一个运动周期,侧生小叶叶枕运动细胞可增长约20%[7],曲率数学模型验证了该运动模式合理性——运动细胞增长10%,即可实现120°的弯曲运动[31]。
1.3 舞草侧生小叶运动的生理过程
在一个运动周期中,侧生小叶叶枕上部运动细胞和下部运动细胞分别经历去极化和超级化过程[39],质子泵和离子泵以及调节运动细胞体积变化的水通道蛋白参与其中[40]。如图2B所示,质膜发生去极化时,外向K+通道和Cl-通道打开(内向K+通道关闭),渗透活性粒子排出细胞,水势改变,细胞失水收缩;随后,ATPases向内转运H+发生超极化,K+通过电压激活的K+通道重新进入细胞并吸水膨胀,直到某种膨压使质膜去极化[31]。去极化抑制剂安氟醚和超极化抑制剂钒酸盐阻断(或减缓)侧生小叶运动的实验很好地验证了上述结论[12]。电压敏感电极被用于检测运动细胞电势的周期性变化[8,11-13],侧生小叶运动过程中细胞生理变化已被清晰地描述,在一个运动周期中:胞外电位在+20 mV和-56 mV之间变化,膜电位则在-36 mV和-136 mV之间变化;胞外K+浓度和pH分别在9~18 mM和6.1~6.6之间变化[41];穿过质膜的水通量为3×10-7cm3/cm2·s;向下运动速度更快(~50 s),水通量可达到10-6cm3/cm2·s,叶枕体积约一半时间在发生变化(~150 s)[42]。
1.4 舞草侧生小叶运动的信号通路及模型验证
植物细胞的运动或体积调节通常与Ca2+和磷脂酰肌醇信号(IP3)的节律变化密切相关[15,43]。Ca2+可能是去极化的触发因素,Ca2+的流入和Cl-的流出使质膜去极化并向外开放定向K+通道[44]。多种Ca2+通道抑制剂已被证明可以影响舞草侧生小叶次昼夜节律运动周期[14-15]。IP3通过与受体结合可触发细胞内部钙库的释放[44],多种IP3的激动剂和拮抗剂(如Li+[45-46]、新霉素[14,47]等)已被证明可影响侧生小叶运动周期(cAMP可能未参与调节其运动[7])。钙振荡已经在许多物种中被描述和分析,并被用于建模[48-51]。Chen[46]将叶枕简化为一个由50个环(每个环50个细胞)堆叠而成的圆柱体,此模型下Ca2+波于不同方向的扩散系数不同,可同时产生顺时针和逆时针的Ca2+波,Ca2+耦合可形成环绕圆柱体的波;结果表明:该Ca2+振荡模型与舞草侧生小叶运动宏观表现相匹配(图3从左到右依次为周期性、准周期和非周期性运动,从上到下分别为时间序列图和相位图)。
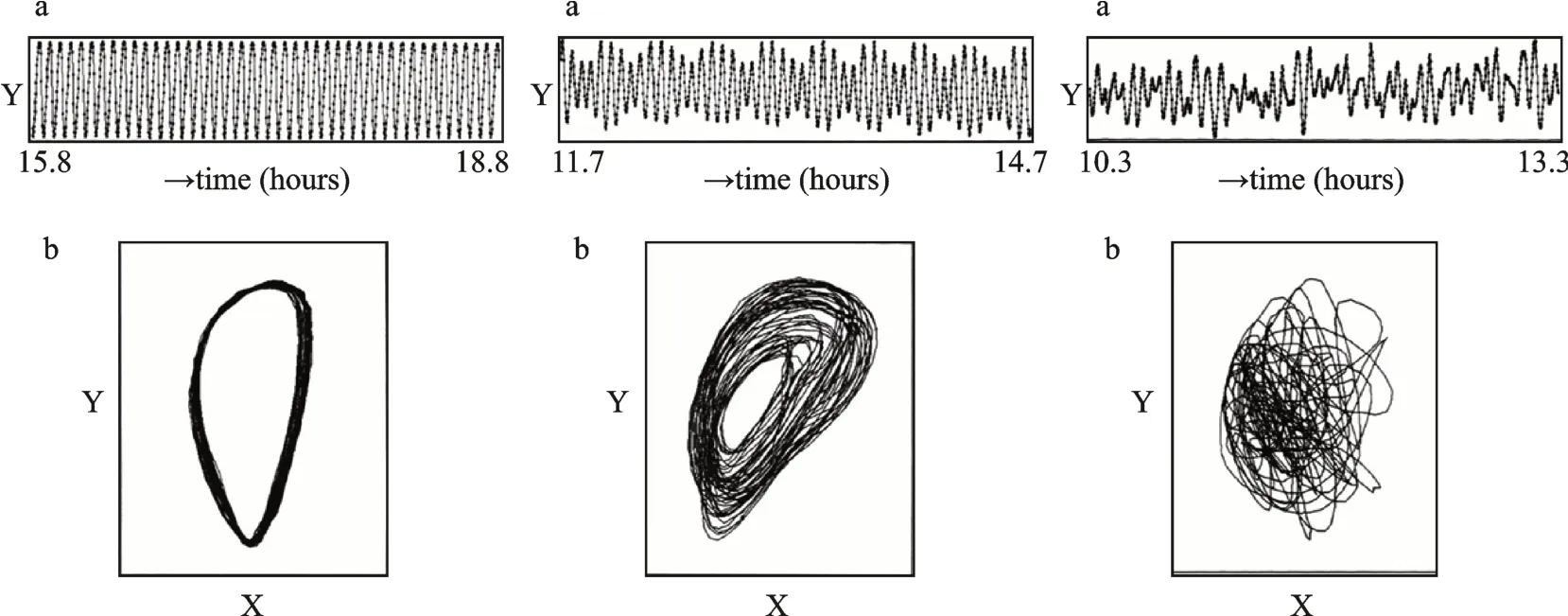
图3 舞草侧生小叶离体叶枕置于去离子水示意图(实验条件为:温度:24~28°C;相对湿度:20%;视频数字化系统记录:6:00~18:00[52])Fig.3 Schematic diagramof isolated leaf pillows of lateral leaflets of dancing grass placed in deionized water(The experimental con‑ditions are:temperature:24~28°C;Relative humidity:20%;Recorded by video digital system:6:00~18:00)
2 舞草侧生小叶对环境因子的响应
由于舞草侧生小叶这种奇特运动模式受到多种环境因子影响,一直让科学家和自然学家着迷。Bose[18]和Mitsuno[8]课题组分别开展了多种物理(光照、温度、负重等)和化学物质(能量转移抑制剂、电子传递抑制剂、解偶联剂和巯基封闭试剂等)对舞草侧生小叶运动的影响研究。
2.1 对光照的响应
舞草侧生小叶在恒定光照(有光或无光)条件下均表现出昼夜节律[53],这与顶生叶的昼夜节律表现不同[7]。舞草侧生小叶离体叶柄在蒸馏水中可以连续振荡多日[7],且在夜间(光照/黑暗条件下为12:12 h)剪取的叶柄会摆动较长时间(≥20 h),而清晨剪取叶柄则摆动时间较短(≤10 h)[31]。可见,侧生小叶的次昼夜节律运动与昼夜节律既区别又有联系,光/暗循环可以调节侧生小叶运动。当光照水平低于20µmol·m2·s-1时,侧生小叶的运动周期近乎恒定[23];在完全黑暗时,运动周期略有下降,但仍可持续约24 h[8](运动停止后,给予光照即可恢复,推测与组织内能量状态有关[23])。侧生小叶在黑暗中的运动一般呈现规律的椭圆或上下运动,无规运动偶有发生[42]。在光照水平为0.1 nmol·m2·s-1的昏暗条件下(24°C),施加中等强度光照(5 nmol·m2·s-1)可以降低侧生小叶振幅,使运动周期减少43%[24],且总是朝着光(近轴或远轴)的方向摆动;这种运动可在长时间(10 min)中等强度(5 nmol·m2·s-1)或短时高强度光照(12 nmol·m2·s-1)暴露后停止[24]。舞草侧生小叶叶枕可同时作为向光运动的感受器和执行器[42],其次昼夜节律优化光合作用与蒸腾作用的能力具有合理性[23]。
2.2 对温度的响应
与舞草顶生叶昼夜节律运动不同,侧生小叶的运动周期强烈依赖于温度(温度系数为2.0[54]或2.5[55])。在环境温度为15°C时,运动周期约为8.7 min;而32°C时,运动周期则变为2 min[55];在距离叶枕5 mm处施加45°C热源(持续30 s)对侧生小叶运动影响较小[55]。两组冷暖交替实验(0°C保持25 s和70°C保持20 s[54]、19.8°C保持4 min和20.3°C保持3 min[31])均发现侧生小叶运动相位具有同步变化趋势(冷-延迟和暖-提前),推测叶枕为温度传感器[54]。
2.3 对负重的响应
物理负重可以明显改变舞草侧生小叶运动节律[16-17]。小叶平均重量约为1.5 mg,负重3~12 mg时运动周期增加10%,且振幅降低[17];20 mg负重可使周期增长约16%,振幅降低50%;25 mg负重可使侧生小叶运动完全停止[18]。针对负重对舞草顶生叶昼夜节律影响的研究未见报道,但负重对紫叶酢浆草(Oxalisregnellii)叶片昼夜节律运动周期没有影响[17]。舞草侧生小叶次昼夜节律运动具有不同于昼夜节律叶片运动的独立潜在机制,次昼夜节律运动调控机制对外部受力敏感,而昼夜节律则对受力不敏感或存在压力补偿[31]。
2.4 对电和磁的响应
Bose[18]和Dutt[13]率先发现直流电可以刺激舞草侧生小叶运动。侧叶顶端施加10 s的直流电(10~100µA),可观察到相位延迟[56]。电脉冲施加在侧生小叶叶枕位置可抑制其运动,暂停运动后一段时间可自发重新开始[19],外部电流信号可能作用于电压门控通道而引起叶枕细胞内外电位的变化[20]。无论静态磁场还是动态磁场,均匀磁场还是非均匀磁场,都可以影响小叶节律[31]。50 mT静态磁场可以使侧生小叶运动减慢,运动周期增加10%,这种效应是通过叶枕组织中的电过程介导的[22]。27 MHz电磁波(≥8 W/cm2)可以改变舞草侧生小叶运动的振幅、周期和相位[21],这与Ca2+浓度[57]或ATP浓度[58]效应相关。
3 生态意义
植物对环境的适应性极为复杂,可通过运动、形态变化以及挥发性物质对环境做出敏感反应,为了生存、生长和繁殖,它们进化和“学习”抵抗各种环境胁迫。运动细胞对环境的响应似乎难以界定主动和被动[59],将复杂行为(智力)与植物联系起来的概念在学术界仍是极具争议的话题[60],但其行为背后存在复杂的利弊权衡(生态意义)却是不争的事实。舞草侧生小叶运动究竟是对非生物因子的感性运动[10,27-28]还是对生物因子的防御行为[29]?感性运动的对象是谁?感受对象间是否存在相互作用及量化关系如何?起源如何?
3.1 对非生物因子的感性运动
3.1.1 光、热和水
植物一般都采用向上的叶片运动(低速生长)作为逃避不利生长条件的机制,触发偏下性生长的信号包括气态激素乙烯、低光照强度和高温刺激[61]。向日葵向光生长可以优化光合作用,获得最佳光照条件和传粉条件,受植物自身生理节奏及外部环境刺激共同调节[62]。含羞草复叶张合速度与环境状况(光照条件[63]和营养状态[64])密切相关,存在对获取光照与降低被采食风险之间的利弊权衡:资源有限时,复叶打开速率较快——此时的主要矛盾为能量缺乏,被捕食风险则为次要矛盾;反之亦然。正所谓生态学是自然的经济学,成本与收益的权衡始终贯穿始终[65],内因与外因相互作用产生了个体行为,形成最佳的生存策略[66]。
舞草侧生小叶叶枕可以作为光照[42]和温度[54]的感受器和执行器,作为光合作用和蒸腾作用“调制器”存在理论上的可能性[10,28],但缺乏实验验证。同时,针对光合作用的主要原料——组织内外CO2和H2O含量对侧生小叶运动影响的研究未见报道,叶枕(和/或侧生小叶)作为感受器的感受行为还有待深入发掘。CO2可诱导拟南芥气孔密度和大小等的改变[67-68],水分胁迫可使植物个体自然振动频率降低15%[69];而叶片运动可以促进空气湍流,降低叶表面的温度[70]。既然侧生小叶旋转是为达到最佳生长条件,过程中势必协调光合、蒸腾及呼吸速率等,对舞草侧生小叶周围边界层多种理化指标及运动响应的定量研究,有助于全面理解其生态意义,对理解其他植物行为同样具有参考价值。
3.1.2 电、磁和力
舞草在印度被称为“电报植物”,侧生小叶旋转运动表现与“旗语通讯”形态相似。事实上,相邻植株间的确广泛存在着地上和地下的“无线通信”[71]。近期发现不同番茄个体可通过土壤进行根际间快速的电信号传递[71-72]。电刺激可诱导植物离子通道和离子转运激活、基因表达、酶系统激活、电信号传递、加速伤口愈合、修复细胞损伤、影响植物生长及植物运动等[73-74]。此外,熊蜂接触花朵引起的电势波动可引导其它熊蜂的授粉活动[75],那么除了啃食或化学物质外,植物个体间及植物与动物间是否存在空气介导的电信号通讯还有待发现[71]。舞草侧生小叶叶枕存在对电流的响应已被证实[20],舞草之间及舞草与周围动物之间是否存在地上或地下电信号通讯还不得而知,有待进一步开展研究。
舞草侧生小叶对磁场[22]和电磁波[21]均有响应行为。由于抗磁各向异性粒子或分子(重力感应细胞中的淀粉颗粒或生物膜中的有序磷脂层[31])的存在,叶片运动过程中在空间上产生不均匀的梯度场,进而会发生趋磁弯曲运动[76],但磁场方向是否对舞草侧生小叶运动节律起决定作用不得而知[21]。植物可通过调控基因表达和表型(植物生长发育、结构组成等)对不同磁场进行感知并快速响应[77],定性定量研究其响应关系——尤其是磁场逆转对植物基因的影响,在遗传和进化等领域都具有十分重要的现实意义[77]。舞草兼具昼夜节律和次昼夜节律运动表现,同时具有明显感磁运动能力,在上述领域应当能够扮演非常重要的角色。
植物对声音振动的刺激响应最近引起人们的关注[78]。植物在不同水平(基因水平、细胞水平和代谢水平等)对振动做出响应被证明可增加农作物产量[79]、增强植物对病原体的免疫力[80]或诱导增强干旱胁迫响应[81];植物可以从风(或虫鸣)引起的振动中识别出昆虫幼虫的咀嚼声[82]或授粉蜂的嗡嗡声[83],并做出具体响应——防御反应或释放花粉,这种现象格外让人着迷。民间流传舞草可随音乐节奏舞动虽未被证实,但其侧生小叶节律运动的确对外部机械压力敏感[31],对触摸和振动均有响应行为[2]。
值得一提的是,在太空失重条件下,拟南芥叶片出现了自发的次昼夜节律运动(颤动),平均周期约为45 min[84]。太空实验认为重力不是维持植物正常生长和正常发育所必需的[85],但对其适应机制相当灵活。为弥补重力的不足,植物可增强其他的向性运动,如:向蓝光性[86]和超敏向湿性[86]。拟南芥叶片的次昼夜节律可能是为弥补重力缺失和昼夜周期缺失而产生的“潮汐节律”[87]。Fisahn等整合了1924-2012年间多种豆科植物昼夜运动节律与地表重力加速度之间的研究报告,发现上述数据与日-地-月之间运行关系呈现明显相关性,豆科植物昼夜运动节律同时具有明显日月引潮力引起的潮汐节律特征[88],且主要作用部位是叶枕[89-90],跟细胞内外水状态密切相关[88]。那么,舞草作为一种豆科植物,其侧生小叶叶枕会感知重力变化,其旋转运动是否是一种比其他豆科植物更精确的重力响应行为以及重力场和磁场是否具有协同作用[91],仍需进一步研究。
3.2 对生物因子的防御行为
植物面对复杂的外部环境并不是完全被动的,尤其在面对采食者时,其防御行为更是千变万化,协同进化的模式更是千姿百态[92]。尽管舞草侧生小叶旋转作为防御行为的研究没有数据支撑,但舞草虫害的确较少[93]。
3.2.1 躲避
拟南芥叶片遭受昆虫啃食后,会产生被啃食叶柄的快速变形-恢复运动(~10 min)和相邻叶片慢速向下运动(~15 min),产生或依赖于慢波电位进行传递,与谷氨酸受体样蛋白[94]和木质部密切相关[95]。被啃食叶柄的快速变形-恢复运动与含羞草感性运动模式极为类似,此类运动可能具有共同的进化起源[95],侧生小叶运动是否也是进化过程中为抵抗昆虫啃食保留下来的防御机制有待进一步研究。
3.2.2 伪装行为
舞草侧生小叶旋转被认为是一种伪装防御行为——可能是模拟具翅节肢动物,以驱使同类离开或吸引其天敌[29],降低被捕食风险。Lev-Yadun将植物伪装防御分为模仿非植物型和模仿植物型[96]:前者通常伪装成对食草动物不具吸引力或厌恶(视觉、嗅觉、触觉等)的其他物体(石头、某种天敌、粪便或腐肉等)[97];而后者通过对自身生长状态的调整降低对食草动物的吸引力(失绿、营养不良、萎蔫、隐藏天敌或已受攻击等)[98]。无独有偶,赭色香碗豆(Lathyrusochrus)半透明的豆荚被认为是在模仿毛毛虫,可减少大型食草动物的采食或吸引天敌捕食真正的食草动物[99]。将植物行为的生态意义拓展到更高营养级具有十分重要的意义[100],但优先策略应该是抵抗植食性昆虫的取食偏好性和产卵偏好性。
3.2.2.1 抗取食偏好性
一般认为含羞草叶片的感触运动具有恐吓作用,但以舞草侧生小叶形态和运动速率来看,其恐吓效果存疑。植食性昆虫普遍存在着取食偏好性[101]——优先采食气味佳、口感好、营养丰富的植物种和个体。旋转运动着的叶片可能不是昆虫优先考虑采食的对象。风引起的叶片被动运动甚至被认为可以达到防御效果[102-103],叶片可运动性更强的植物被采食的风险更小,植株体附着无脊椎动物的种类和数量也越少。植株体叶片的可运动性(叶柄长短、叶片形状等)可能是防御效率和抗风损伤之间利弊平衡的结果[103]。Pantin认为气孔对湿度响应行为让人迷惑,难以区分主动运动和被动运动的界限[59]。舞草侧生小叶是否可以干扰昆虫移动或屏蔽昆虫通讯,从而达到抵抗昆虫采食的目的?
物理屏障是植物抵御害虫的第一道防线,几乎所有的植物都特化出抵抗食草动物某种结构或物质,广泛存在植物体表面的蜡质层、表皮毛或粘液被认为在阻止食草动物附着或移动方面具有重要作用[104]。那么,舞草侧生小叶的旋转是其所特化出来的一种结构,类似于风引起的被动运动,在一定程度上可以干扰昆虫附着或移动(向顶生大叶),达到让采食者“讨厌”的目的。
生物体在各层次或尺度(亚细胞过程、生物与环境的相互作用等)都遵循物理法则[105],基于生物力学的生物通讯也不例外。振动信号传递对植株、采食者和捕食者及其相互之间信号通讯起着至关重要的作用[106],如:寻找配偶或猎物[107]、躲避捕食者[108]、花粉释放[83]等。振动传递在叶脉及叶片不同区域传播效率不尽相同[109];叶片性状(叶面积、含水量、厚度等)对振动的传递起着至关重要的作用,叶脉密度可能是水分传递(生长)与振动传递(防御)权衡的结果[110]。植株振动消耗的阻尼来源一般为外部流体、软组织及向分支传递能量[70]。舞草侧生小叶叶枕具有软组织特征,其旋转运动存在潜在的消耗(或阻止)振动信号传递的功能,以切断采食者求偶信号或捕食者接近的振动信号;采食者这种状态下无法进行振动通讯,进入一个“振动屏蔽”区域而无法获益(或直接受害),经过长期进化将舞草从“食谱”上删除。
3.2.2.2 抗产卵偏好性
植食性昆虫产卵行为在昆虫与寄主的协同进化中起着重要作用[111]。产卵行为会与植物产生复杂的机械或化感作用,诱发植株产生一系列防御反应(次生代谢物的合成、防御蛋白的表达、营养分配的改变等[112-116])。
植食性昆虫普遍存在着产卵偏好性[117]——成虫通过多种感官对寄主种类、个体特质或具体部位等的选择性。草地螟(LoxostegesticticalisL.)偏好叶片长宽比高的植株个体[118],而棉铃虫(Helicoverpa armigeraHübner)则在掌状叶上产卵量较大[119];烟粉虱(BemisiatabaciGennadius)一般选择叶片背面产卵(防止雨水冲刷和阳光直射)[120];而眉斑并脊天牛(GleneacantorFabricius)[121]和毛萤叶甲(Pyrrhal⁃taviburniPaykull)[122]偏好群聚产卵(已被侵害的病态植株),广聚莹叶甲(OphraellacommunaLeSage)则喜好独居产卵(未被侵害的新鲜植株)[123]。有学者认为舞草侧生小叶运动是在模拟某些具翅节肢动物,以传递“已占勿扰”的信息[29],但具体哪些种类的具翅节肢动物偏好群聚或独居产卵还需要深入了解。细致调查昆虫行为偏好性对于理解舞草侧生小叶旋转生态意义和进化起源具有重要的现实意义。因此,舞草侧生小叶的旋转在视觉上为一些母代产卵所厌恶的一种形式。但是否或多大程度上降低产卵行为发生的概率,侧生小叶旋转时边界层挥发性成分是否发生了改变,以及如何实现能量收支和防御效率等的相互协调,这些都需要进一步探究。
4 总结与展望
舞草兼具昼夜节律和次昼夜节律运动,对多重(种)环境因子具有响应能力,还有许多未知等待解答。目前对舞草侧生小叶运动的研究主要集中在解剖学、电生理学、细胞生物学等方面,但在分子生物学、遗传学和生态意义方面的研究需要进一步深入。应用现代的分析监测仪器和先进的分子遗传学方法,融合动物行为学、生物力学及化学生物学等相关学科,对舞草侧生小叶运动发生前后体内外痕量理化性质的分析、侧生小叶叶枕缺陷突变体的诱导、跨食物链动物行为的试验分析、多学科理论及技术的交叉研究等,有助于全面了解舞草侧生小叶旋转的起源和进化意义。
影响舞草侧生小叶旋转运动的主导环境因子是非生物还是生物,究竟是感性运动、向性运动还是防御行为还不清楚。侧生小叶运动的自主节律和环境因子是否及如何发生交互作用,能量收支如何平衡有待解答。植物激素在此过程中是否发挥作用,其运动起源究竟是为躲避昆虫啃食、退化的攀援能力还是类生长介导的运动不得而知。植物长期进化形成防御行为具有极高的复杂性和动态性,许多新机制和新系统还有待发掘。开展对舞草侧生小叶运动全方位解读对于理解其它植物种行为的生态意义同样具有重要现实意义。
