m6A表观遗传修饰及其调控机制研究进展
2022-02-15史源钧米思远
史源钧,米思远,俞 英
(中国农业大学动物科技学院,北京 100193)
畜禽拥有许多复杂的经济性状,近十余年来,研究者已通过全基因组关联分析等方式鉴定到了大量与这些经济性状密切相关的单核苷酸多态性(single nucleotide polymorphism,SNP)位点,但是这些位点大多数位于基因组的非编码区[1-3]。基因组的遗传信息通过何种方式影响畜禽的复杂性状仍未得到全面地阐明。生物中心法则指出,遗传信息由DNA流向RNA,最后流向蛋白质,在这一过程中往往伴随着复杂的调控模式。RNA作为遗传物质表达过程中最为重要的中间环节,有160余种RNA修饰被报道,这些修饰主要参与基因的转录及转录后调控过程[4],如N6-甲基腺嘌呤(N6-methyladenosine,m6A)、N1-甲基腺嘌呤(N1-methyladenosine,m1A)[5]及5-甲基胞嘧啶(5-methylcytosine,m5C)[6]等修饰(图1)[7]。其中,m6A在真核生物RNA修饰中丰度最高且具有较高的保守性,其最早在1974年被发现于真核生物mRNA及lncRNA中[8]。研究发现,m6A修饰广泛分布于人类细胞中超7 000个mRNAs及300个lncRNAs中[9]。据估计,分离出的RNA中m6A甲基化修饰的量占总腺苷量的0.1%~0.4%(即每条mRNA有3~5个m6A修饰位点),且主要在终止密码子、3′-端及内部长外显子中富集[10]。此外,近期研究发现,被修饰的RNA含有共有基序RRACH(R=A/G,H=A/C/U)[11]。目前,人类及模式生物上大量的研究已经表明,m6A广泛参与生长发育、代谢及炎症反应等相关过程[11-12]。作者就哺乳动物m6A研究的最新前沿动态进行了阐述,并重点探讨了m6A对于畜禽复杂经济性状的意义及相关应用前景。
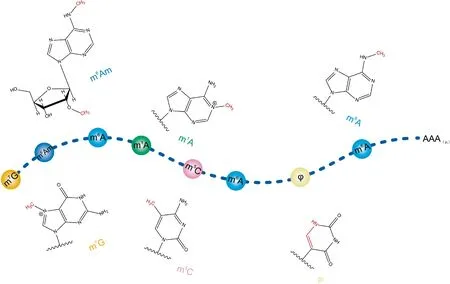
图1 RNA常见修饰方式及其常见位置(修改自[7])Fig.1 Overview of common nucleotide modifications on RNA (modified from[7])
1 m6A修饰的分子机制
m6A甲基化修饰是RNA中腺嘌呤上第6位的氨基发生甲基化的一种修饰方式,在生物体内是一个可逆的过程,在细胞核内由甲基转移酶复合物(writers)、去甲基化酶(erasers)共同作用,以调控m6A在RNA上修饰的位点及丰度,并由读取蛋白(readers)调节RNA可变剪接、出核运输、翻译及降解过程(图2)。
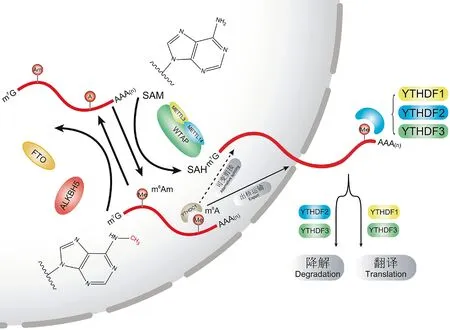
图2 m6A甲基化修饰机制Fig.2 The mechanism of m6A methylation
1.1 m6A甲基化酶复合物
研究显示,m6A甲基化主要由S-腺苷甲硫氨酸(S-adenosylmethionine,SAM)作甲基供体,通过大型甲基转移酶复合物(m6A methyltransferase complex,MTC)来实现整个甲基化过程。 MTC由甲基转移酶METTL3(methyltransferase-like 3)和METTL14(methyltransferase-like 14)组成的功能性异源二聚体为催化核心,由WTAP(Wilm’s tumour-1-associated protein)蛋白作其载体,最后由其他相关蛋白因子如KIAA1429、RBM15等共同完成m6A甲基化的过程[13]。在这个大型MTC中,METTL3可以催化SAM,将甲基由SAM转移至腺苷酸上;METTL14作为METTL3的同源物,可以稳定METTL3的空间构象保持其生物活性,并识别可甲基化的RNA,促进甲基化过程[14]。WTAP作为甲基转移酶复合物中第3个关键组成成分,募集由METTL3和METTL14组成的异源二聚体;同时还可以募集可甲基化的mRNA及lncRNA,并加速甲基化过程[15]。研究证明,在HeLa细胞中敲低METTL3,m6A修饰水平降低了30%;同样,敲低METTL14或WTAP,HeLa细胞整体m6A修饰水平均有不同程度的下降[9]。虽然三者甲基化活性相对偏低,但因其相结合形成稳定的复合物后可显著提升彼此的甲基化活性,由METTL3、METTL14及WTAP组成的MTC甲基化活性显著升高[16]。此外,METTL3和METTL14的同源物METTL16也可利用SAM单独完成m6A甲基化过程[11]。一般情况下,m6A甲基化过程需要在细胞核内完成,但在一些特殊情况下(如病毒的复制)也可在细胞质内完成[17]。
1.2 m6A去甲基化酶
m6A去甲基化酶主要功能是去除受m6A修饰RNA上的甲基基团。目前已被发现的m6A去甲基化酶只有ALKBH5(ALKB homolog 5)和FTO(又名ALKBH9,ALKB homolog 9) 2种。 由于ALKBH9最早被发现与肥胖有关,因此命名为FTO(fat mass obesity associate)[18],二者同属ALKB族蛋白,且在发挥作用时都需要氧气及二价铁的参与[19],但二者在去甲基化的机制上有所不同。ALKBH5主要在细胞核内富集,除去甲基化外还可调节mRNA的加工、代谢及出核运输[20],其去甲基化过程一步完成,不产生任何中间体[21]。而FTO在细胞质内的去甲基化过程中主要通过分步氧化的方式完成:首先将甲基(m6A)氧化成羟甲基(N6-羟甲基腺苷,hm6A);之后再将其进一步氧化成甲酰基(N6-甲酰基腺苷,f6A);最后在多种酶的作用下还原成腺苷[22]。这种方式从动力学角度来看反应更容易进行,FTO通过分步氧化甲基,不断加大修饰基团与腺苷之间的极性,更利于修饰基团的脱除。此外,FTO还可以调节前体mRNA的剪切[23-24]。
1.3 m6A读取蛋白
m6A读取蛋白主要由YTH域蛋白组成,包括2个亚型:YTHDFs(YTH domain family proteins)和YTHDCs(YTH domain containing proteins)。YTHDFs主要包括3种蛋白:YTHDF1、YTHDF2和YTHDF3;YTHDCs主要包括2种蛋白:YTHDC1和YTHDC2[25]。此外,除YTH域蛋白外,m6A读取蛋白也包括eIF3和hnRNPs等多种蛋白。除了少部分读取蛋白在细胞核内发挥作用外,大多数作用场所在细胞质内。对于YTHDFs蛋白而言,不同的YTHDF蛋白会结合到不同的m6A修饰区域上[26],并影响下游被修饰RNA的可变剪接、翻译、降解及RNA稳定性[27]。 从实际功能上看,YTHDF1、YTHDF3、YTHDC2及eIF3蛋白可诱导经m6A甲基化修饰的mRNA进行翻译。但由于m6A甲基化在终止密码子附近富集,远离翻译起始位点[28],m6A对mRNA翻译的促进作用仅存在于可环化的RNA中[29],其原因可能是由于mRNA发生环化后,终止密码子和起始密码子相邻导致m6A可促进mRNA的翻译。 此外,YTHDF2、YTHDF3、YTHDC1及YTHDC2可将m6A诱导至P-body[30],并募集CCR4-NOT复合物,将转录本腺苷酸化并降解[31]。在热应激情况下,YTHDF2可转移至核内,通过限制去甲基化酶FTO的去甲基化过程来维持体内m6A甲基化水平的稳定[32]。由于被m6A修饰的mRNA区域通常缺乏二级结构,致使YTHDC1与hnRNP有利于与其结合从而发生选择性剪切[33-34]。
2 m6A修饰的检测技术
为检测m6A修饰整体及特异位点的丰度,研究者基于液相色谱、质谱、免疫沉淀及测序等多种技术手段开发出了定量、定点的m6A甲基化修饰检测方法(图3)。考虑到试验的高效性及准确性,目前针对m6A的定量分析主要有Dot Blot法[35]、液相色谱-质谱联用(LC-MS)及基于液质联用优化出的高效液相色谱-串联三重四极杆质谱技术(LC-MS/MS)[36]等方法。m6A整体丰度的改变只能在一定程度上说明m6A修饰发生变化,但若是在检测中发现整体丰度无明显变化并不能说明m6A修饰无变化,有可能是部分修饰位点发生变化而整体丰度变化不明显[37],因此,需要在后续针对m6A修饰的位点及单基因m6A修饰的变化进行进一步检测。目前,对于m6A位点的定位检测主要有基于免疫沉淀技术及测序技术的m6A测序[9](m6A-seq)、m6A免疫沉淀测序[10](methylated RNA immunoprecipitation sequencing,MeRIP-seq)、m6A单核苷酸分辨率交联与免疫沉淀定位法[38](methylation individual-nucleotide-resolution crosslinking and immunopre-cipitation,miCLIP)等技术。而针对单基因的检测主要是甲基化RNA免疫共沉淀实时荧光定量PCR(methylated RNA immunoprecipitation quantitative Real-time PCR,MeRIP-qPCR)和利用m6A位点进行特异性切割及放射性标记后再进行连接辅助萃取和薄层层析方法(site-specific cleavage and radioactive-labeling followed by ligation-assisted extraction and thin-layer chromatography,SCARLET)2种,但二者存在着不同的优缺点。MeRIP-qPCR操作简便但精度不足,无法定位到单碱基;SCARLET可定位到单碱基水平但其操作繁琐,不适合大范围使用[39]。近几年,研究者开发出了一系列不需要抗体的单基因检测技术,如基于单碱基延伸和连接的qPCR法[40](single-base elongation- and ligation-based qPCR amplification method,SELECT)和FTO辅助的化学交联法[41](FTO-assisted chemical labeling method,m6A-SEAL)等。此外,单分子实时测序(single molecule Real-time seqencing,SMRT-seq)[42-43]和单分子纳米孔测序[44]也为RNA修饰的检测提供了新的可能。
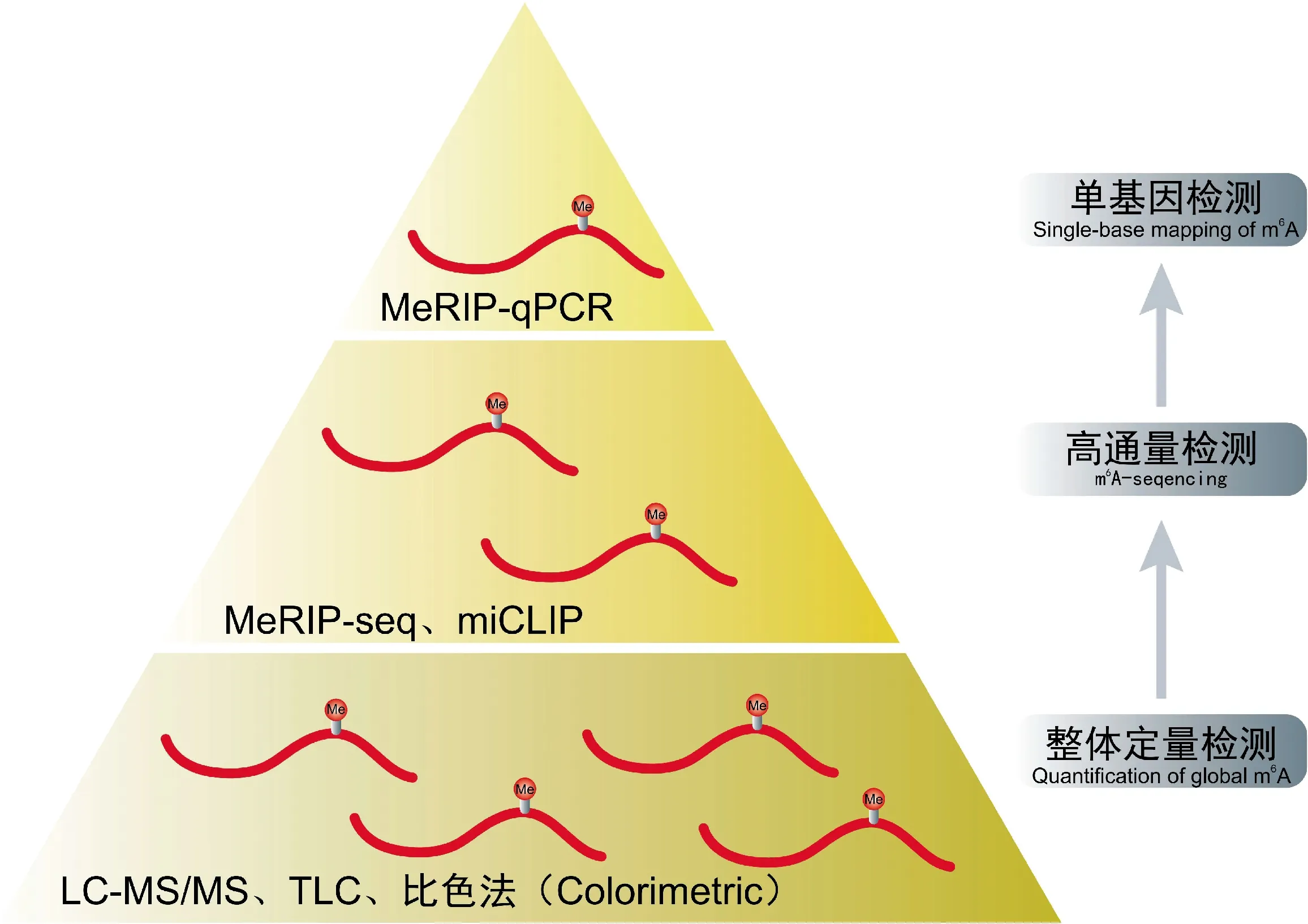
图3 m6A主要检测方法Fig.3 The main detection techniques of m6A
3 m6A修饰在动物复杂性状方面的研究
3.1 m6A调控动物生长发育
由于m6A在生物体内分布极为丰富,近年来关于m6A对动物体生长发育影响的研究也越来越多。研究表明,m6A甲基化修饰对脂肪的形成起到了关键的调节作用,在脂肪组织中m6A去甲基化酶FTO及读取蛋白YTHDF2均可通过介导ATG5、ATG7转录本m6A修饰丰度影响其转录本丰度和脂肪前体细胞自噬过程,最终调节脂肪组织的形成[45]。在家养动物上,断奶仔猪饲粮中添加支链氨基酸(branched-chain amino acids,BCAA)可显著下调腹侧、背侧及肝脏脂肪组织中m6A的总体丰度,从而影响其脂质代谢及沉积[46]。在肌肉发育的不同阶段m6A修饰也发挥了重要的调控作用,如在骨骼肌发育相关通路中大量蛋白转录本均受到不同程度m6A修饰的调控,随着骨骼肌的发育,m6A读取蛋白胰岛素样生长因子2 mRNA结合蛋白1(insulin-like growth factor 2 mRNA-binding protein 1,IGF2BP1)表达量在不断降低,为了证明该过程中m6A修饰发挥了关键作用,研究人员在成肌细胞中分别敲降IGF2BP1以及m6A读取蛋白METTL14,发现肌细胞出现了类似表型变化[47]。此外,在动物表皮mRNA的m6A修饰差异同样显著影响到皮肤组织及动物绒毛的发育[48-49]。
研究发现,动物性情对动物经济性状及动物福利有显著影响[50],在近期对神经发育及疾病的研究中同样证明了m6A对大脑发育、衰老及阿尔兹海默症有重要影响。据报道,5XFAD阿尔兹海默症小鼠3′-端非编码区m6A水平显著低于野生型对照小鼠,而这些差异甲基化区域内的基因参与了在阿尔茨海默症发病过程中受到影响的生物过程,如突触传递和离子转运调节等[51],因此,在今后m6A修饰对畜禽神经系统发育的影响同样值得关注。
3.2 m6A对于畜禽繁殖的影响
在畜禽繁殖及发育中同样发现m6A发挥了重要作用。研究表明,m6A甲基化修饰及其相关调控因子在诱导猪多能干细胞(porcine induced pluripotent stem cells,piPSCs)分化中发挥了重要的调控作用。由于Janus激酶2(Janus kinase 2,JAK2)和细胞因子信号抑制剂3(suppressor of cytokine signaling 3,SOCS3)转录本的3′-端存在m6A修饰,METTL3缺失会导致JAK2和SOCS3转录本m6A水平降低。由于这2种基因编码的蛋白在JAK2-STAT3通路中发挥了重要作用,其表达量的下降进一步导致了JAK2-STAT3通路的沉默,从而阻断Kruppel样因子4(kruppel like factor 4,KLF4)和SRY转录因子2(SRY-box transcription factor 2,SOX2)基因的转录,而这2种基因编码的蛋白在维持细胞多能性及自我更新中发挥了重要作用,其缺失会显著影响piPSCs的自我更新并诱导其发生分化[52]。此外,在正常生殖细胞的发育过程中m6A甲基化修饰同样起到了重要的调控作用,如m6A对卵泡的发育过程(外层颗粒细胞对卵泡的发育、成熟及排卵)有重要的调控作用[53]。而在颗粒细胞中m6A存在高丰度的修饰,且修饰丰度会随着卵泡的发育发生动态的变化,在颗粒细胞增殖、类固醇分泌及卵泡发育过程中,颗粒细胞内参与m6A甲基化修饰的基因均出现显著的差异表达[54]。
3.3 m6A对畜禽热应激的影响
近几十年来,全球变暖导致世界各地出现了持续性高温及频繁的极端炎热天气,由于奶牛、猪及家禽等家养动物受热应激的影响较大,给养殖业带来了巨大的损失。研究发现,转录组m6A甲基化水平会随着热应激的发生而出现显著变化,而KEGG通路富集分析显示,这些差异m6A修饰的基因主要富集在Wnt、转化生长因子-β(transforming growth factor-beta,TGF-β)等与应激响应和脂质代谢相关的通路[55]。m6A甲基化调节了热休克蛋白(heat shock proteins,HSPs)基因的表达,而m6A修饰的HSPs转录本的差异表达可能取决于m6A位点及其丰度[56],一方面,随着热应激的发生,m6A甲基化相关调控因子表达量升高,导致热休克蛋白(HSP70、HSP90、HSP110)转录本m6A甲基化水平和表达量出现明显上升,从而影响动物对于热应激的反应[57];另一方面,热应激提升了部分mRNA的5′-端非编码区m6A甲基化修饰丰度,增强了mRNA的翻译。值得注意的是,在此过程中m6A甲基化读取蛋白YTHDF2从细胞质内几乎全部迁移至细胞核内发挥作用,且在热应激发生后,YTHDF2蛋白表达水平与HSP70出现了同步上调,细胞m6A甲基化丰度增加了近4倍[32]。此外,热应激会影响畜禽的生产性状,当仔猪发生热应激反应后,肝脏及腹部脂肪组织中m6A甲基化相关调节因子的表达水平出现显著上调,参与脂肪代谢相关基因的表达量出现了显著上升。在此过程中,肝脏及腹部脂肪组织中脂肪代谢基因转录本可能受到m6A甲基化的调节,从而影响了仔猪后续脂肪的沉积[58]。同时,生物为了应对热应激,在生命早期可通过m6A去甲基化酶FTO影响脑营养因子(brain-derived neurotrophic factor,BDNF)转录本m6A甲基化修饰丰度,进而影响其表达量,从而改变生物后期对热应激的反应[59]。而BDNF作为体内含量最多的神经营养因子,其广泛存在于中枢及周围神经系统中[60],对生物神经系统发育、精神疾病及某些癌症均有重要影响[61-62]。
3.4 m6A对于炎症反应及癌症的调控
越来越多的研究表明,m6A修饰在炎症反应中发挥着至关重要的作用。引起炎症反应的病原因素主要有病毒和细菌两大类,如许多编码抗病毒蛋白的干扰素刺激基因(interferon stimulating genes,ISGs)的转录本被m6A修饰,m6A的修饰促进某些ISGs的翻译,增强干扰素的抗病毒作用[63];但也有研究表明,病毒自身的m6A甲基化修饰可通过抑制双链RNA(double-stranded RNA,dsRNA)的形成,削弱Rig样受体(Rig-like receptors,RLRs)对病毒的识别,从而促进病毒的免疫逃逸[64]。此外,在某些DNA病毒感染下,m6A调节因子hnRNPA2B1可识别病毒DNA促进m6A修饰,从而触发先天免疫反应[65]。在对家畜的相关研究中发现,猪流行性腹泻病毒受到大量m6A修饰的调控,宿主甲基化酶METTL3、METTL14及去甲基化酶FTO对病毒RNA的m6A修饰起到了重要的调节作用,读取蛋白YTHDF可通过影响病毒RNA的稳定性从而影响病毒的复制,该研究证明在病毒感染后,宿主通过下调去甲基化酶FTO使宿主m6A修饰丰度提高[66]。近期对于2019新型冠状病毒(COVID-19)的研究发现,新型冠状病毒具有m6A修饰,且病毒m6A丰度降低,可促进其与RIG-Ⅰ的结合,并增强宿主细胞下游先天免疫信号通路和炎症基因表达,且新型冠状肺炎重症患者m6A甲基化酶显著偏低[67]。
细菌引起的机体炎症反应可引起机体m6A水平的改变。当细菌感染小鼠角膜后,角膜组织可通过提高甲基化酶METTL3的表达量而提高其整体甲基化水平,从而起到抗感染的作用[68]。除此之外,在动物肠道中也有相似发现,肠毒性大肠杆菌感染后,机体可通过“FOXO6-METTL3-m6A-GPR161信号通路”提高METTL3基因的表达水平,从而提高m6A修饰丰度,促进肠防御素的表达[69]。
在对癌症的研究中发现,m6A修饰对肺癌、胃癌、胰腺癌、肝癌、急性髓系白血病等多种癌症都有重要影响[70]。针对m6A去甲基化酶FTO及甲基化酶METTL3在急性髓系白血病中的异常表达,已有课题组开发出了一系列小分子靶向药物,使m6A的研究具有临床意义[71-73]。
3.5 m6A修饰调控蛋白与畜禽经济性状关系
研究表明,畜禽m6A修饰丰度的高低对其经济性状会产生较大影响,m6A修饰对畜禽繁殖、生长和抗应激等多方面均有重要意义,部分相关研究结果见表1。
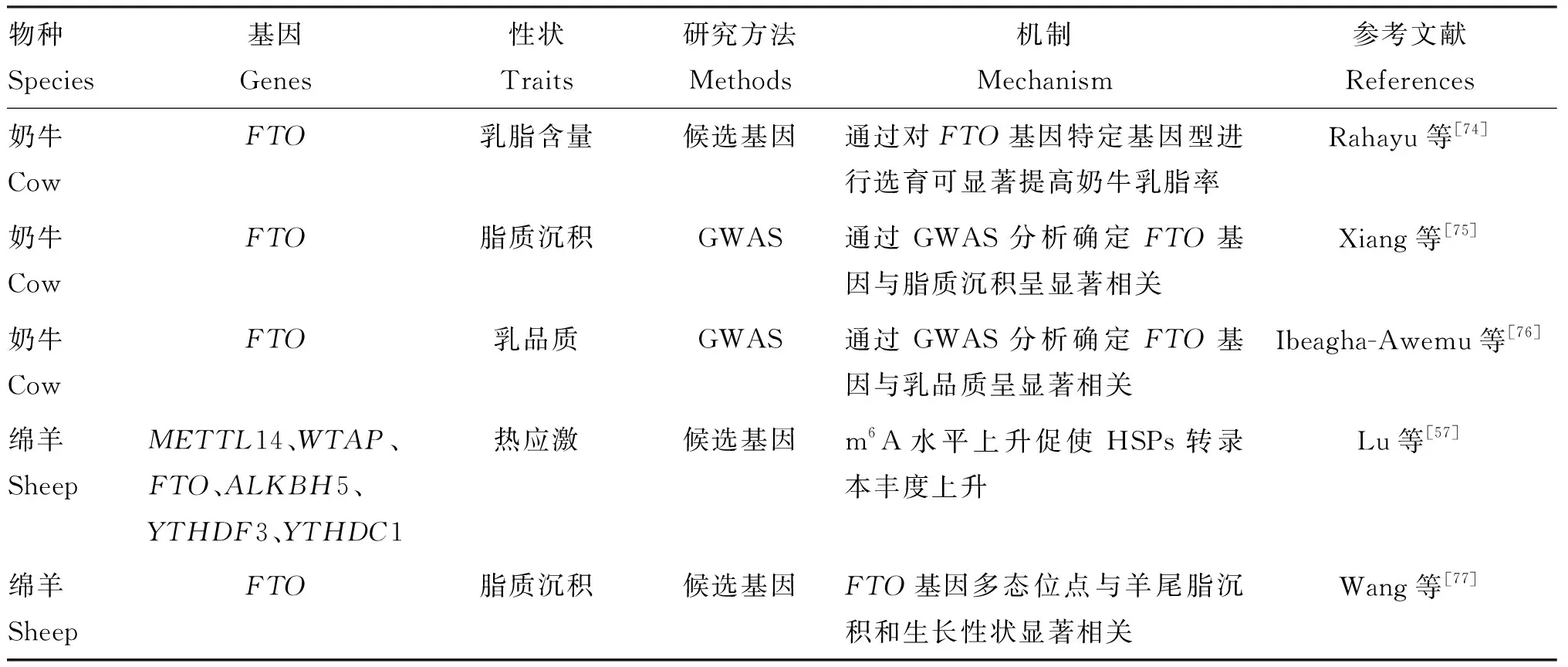
表1 m6A修饰与畜禽经济性状的关系
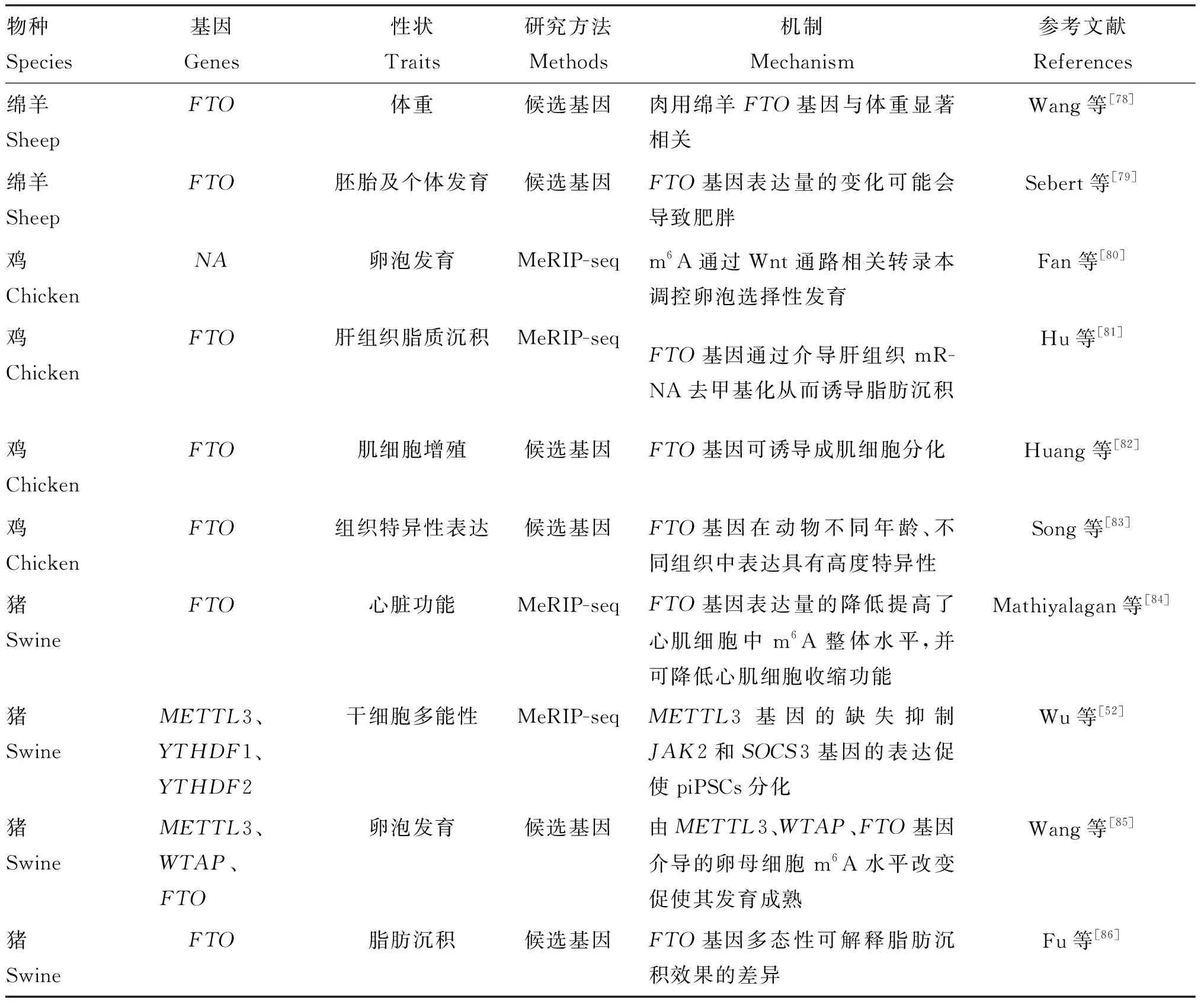
续表
4 m6A甲基化修饰与其他表观修饰之间的联系
除m6A对于RNA代谢的调控外,其他表观修饰形式与m6A之间同样存在紧密联系,如磷酸化、类泛素化修饰对m6A相关蛋白因子的调控、m6A对非编码RNA的调控及组蛋白修饰与m6A之间的互作。研究发现,在m6A对RNA调控的基础上,m6A去甲基化酶ALKBH5及读取蛋白YTHDF2也存在翻译后的类泛素化修饰调控。一方面,经类泛素化修饰后的YTHDF2对于m6A修饰的RNA具有更强的亲和力,可促进其降解[87],而m6A去甲基化酶ALKBH5在经磷酸化及类泛素化修饰之后活性显著降低,并使DNA损伤修复相关蛋白表达量提高,减小了活性氧对DNA造成的损伤[88];另一方面,m6A不仅在mRNA中发挥作用,同时还可影响非编码RNA的剪接、折叠及成熟。在lncRNA中,m6A修饰可通过改变RNA构象增强其与异质性核糖核蛋白C(heterogeneous nuclear ribonucleoprotein C,HNRNPC)的结合能力,并促进其成熟[34]。在细胞对miRNA的加工中也有类似发现,RNA结合蛋白HNRNPA2B1(heterogeneous nuclear ribonucleoprotein A2/B1)可特异性识别m6A并与其所在RNA片段结合,诱导pri-miRNA成熟[89]。
m6A甲基化修饰与组蛋白修饰之间联系最为密切,组蛋白修饰作为经典的表观修饰途径之一,对于生物生长发育、疾病发生、炎症及癌症均有显著影响。研究发现,H3K36me3的富集峰与m6A在转录本中的富集峰在分布上趋于一致,H3K36me3可被METTL14识别并直接结合,随后在转录过程中促进甲基化酶复合物与RNA聚合酶Ⅱ结合,进而在转录过程中引导m6A在转录本中沉积[90]。m6A对组蛋白的修饰同样具有显著影响,如m6A水平下降可引起细胞中H3K27me2水平的整体下降[59];m6A甲基化读取蛋白YTHDF2能抑制白细胞介素6(interleukin 6,IL-6)基因启动子区H3K27me3的去甲基化过程,影响炎症因子的表达,从而影响细菌对宿主细胞的侵染过程[91]。上述研究不仅将m6A修饰与生物生命活动联系起来,更证明了m6A修饰与各种表观修饰之间的相互作用,丰富了表观遗传调节过程。
5 展 望
m6A作为RNA上丰度最高的表观修饰形式,在人类及模式生物中已经开展了大量的研究工作,m6A已被证实在生长发育、繁殖及免疫等众多生物过程中发挥着重要的调控作用。因此,m6A可能会影响畜禽生长发育等很多过程,m6A在畜禽复杂性状形成中所发挥的作用及m6A与其他表观修饰间的互作有待进一步深入探究。尽管目前m6A测序仍然存在费用昂贵、高通量测序所需的RNA起始量较多等缺陷,但相信在不久后的将来,随着测序技术的不断改进与提升,畜禽上m6A方面的数据将得到不断扩充,这将有助于今后对畜禽繁殖过程的解析及遗传改良策略的优化。
