羽衣甘蓝BoWRKY10基因克隆及其转基因烟草耐旱性研究
2022-02-08赵红启李鸿雁
赵红启,杨 勇,闫 艳,范 杰,李鸿雁
(1.黄淮学院园林中心,河南 驻马店 463000;2. 黄淮学院生物与食品工程学院,河南 驻马店 463000)
【研究意义】植物生长在不断变化的自然环境条件下,受到各种生物和非生物胁迫。为了提高生存率,植物必须通过进化和调节生长机制来适应这些恶劣的环境。作为主要的非生物胁迫,干旱是对植物的生长和发育产生不利影响,甚至对农业产量造成严重损失的重要环境胁迫因子之一。因此,研究与植物抗逆性相关的基因及其分子机制,对于植物抗逆力的遗传改良和人类社会的可持续发展具有重要意义。WRKY转录因子在调控机制中扮演着十分重要的角色,近几年备受关注。克隆WRKY相关基因,研究其调控机制,对于阐明植物转录因子的调控机制具有重要意义。【前人研究进展】转录因子(Transcription factors,TFs)是胁迫信号转导和靶基因表达的重要介质。TFs通过调节下游靶基因的表达水平,在保护机体免受应激相关损伤中发挥重要作用[1-2]。大量研究表明,TFs基因的过表达激活了一组靶基因,这些靶基因协同抵御非生物胁迫的不利影响。因此,TFs基因工程是提高作物抗逆性的有效途径。拟南芥(Arabidopsisthaliana)基因组中有1500多个TFs,约占所有基因的6%[3]。在这些TFs中,WRKY作为植物中最大的转录因子家族之一,在植物对干旱胁迫响应的调控网络中发挥重要作用4]。WRKY蛋白是一种新型的植物锌指转录调控因子,其N端含有7个由WRKYGQK组成的氨基酸序列。WRKY蛋白是由1个或2个WRKY保守结构域。WRKY-TF基因受生物和非生物胁迫诱导,参与植物胁迫响应[5]。WRKY-TFs是植物基因组中的一个大基因家族。拟南芥中有74个基因[4],水稻(OryzasativaL.)中有100多个基因[6],小麦(TriticumaestivumL.)中有119个基因[7],小白菜(BrassicacampestrisL.)中有46个基因[4]。WRKYs被认为是参与生物胁迫响应的关键TFs,这些实验数据为WRKY调控非生物胁迫响应提供了证据。此外,一些TFs对干旱胁迫特别敏感。例如,金钱草(DichondrarepensL)WRKY70基因通过调控精氨酸脱羧酶基因在抗旱和腐胺合成中发挥作用。转二穗短柄草(BrachypodiumdistachyonL.)的BdWRKY36基因对干旱胁迫的耐受性显著增强[8]。另外2个WRKY基因(ABO3和ThWRKY4)已被证明是干旱胁迫响应中的关键调控基因[9]。MuWRKY3基因在花生(ArachishypogaeaL.)中的过表达是通过减少MDA、过氧化氢和超氧阴离子的积累以及增加脯氨酸含量、可溶性糖含量和抗氧化的活性显著提高花生的耐旱性[10]。虽然一些TFs在模式植物中具有生理功能,但大多数TFs的功能尚不清楚,特别是在羽衣甘蓝(Brassicaoleraceavar.acephalaDC)等非模式植物中。【本研究切入点】WRKY家族基因在植物抗旱育种中具有广泛的应用前景,前期在拟南芥、小白菜和油菜(BrassicacampestrisL.)等十字花科中发现一些WRKY家族转录因子基因在植物逆境中起重要作用,但在羽衣甘蓝中鲜有研究。【拟解决的关键问题】本研究以羽衣甘蓝为材料,利用转基因技术探究BoWRKY10基因在羽衣甘蓝耐旱性中的生物学功能。探明在羽衣甘蓝中超表达BoWRKY10对羽衣甘蓝抗旱性的影响,明确该基因在羽衣甘蓝响应干旱胁迫的生物学功能其在旱育种中的价值。
1 材料与方法
1.1 试验材料
羽衣甘蓝种质材料“叶牡丹”由黄淮学院园林中心提供。烟草NC89及pCAMBIA1301载体由黄淮学院生物与食品工程学院实验室保存。植物在温室中用混合营养土[m(蛭石):m(营养土)=1∶1]培养,培养温度为24 ℃/22 ℃(白天/黑夜),湿度为65%,光照周期为光照(L):黑暗(D)=16 h:8 h。
1.2 RNA提取与载体构建
用植物总RNA分离试剂盒(TaKaRa,大连,中国)从羽衣甘蓝的叶片中提取总RNA。根据试剂盒的说明,使用逆转录酶试剂盒(TaKaRa,大连,中国)合成第一链cDNA。从NCBI数据库中获得羽衣甘蓝WRKY10基因序列,利用Primer Premier 5.0分别设计了特异的正向和反向引物(表1)。扩增的PCR产物纯化后,被克隆到pMD18-T载体( TaKaRa,大连,中国)上并测序。
1.3 烟草转化与筛选
把测序后正确的序列与含有新霉素磷酸转移酶基因(Neomycin phosphotransferase gene, NPTII)和35S启动子(CAMV35S)的pCAMBIA1301双元载体相连。将pCAMBIA-BoWRKY10导入根癌农杆菌LBA4404中。根据Horsch等[11]的方法转化烟草。将T0转基因植株的种子收获并接种在含有卡那霉素(100 mg/L)的MS培养基上。用qPCR引物扩增BoWRKY10基因(表1)。采用定量PCR(qPCR)技术检测BoWRKY10在转基因植株中的表达。
1.4 试验设计
取适量野生型(WT)和转基因烟草种子于无菌2 mL离心管中,70%酒精消毒30 s,无菌水洗1次,10%(v/v)H2O225 ℃消毒 10 min,无菌水洗5~6次,点播于含有15% PEG4000的 1/2 MS培养基上,黑暗1 d后,28 ℃光照培养,每隔24 h 统计发芽率;正常萌发的种子移栽到 5%、10%和15% PEG4000的1/2 MS条件14 d后,统计根长,每个株系取12株;烟草于温室培养30 d后,15% PEG4000处理,24 h后取材料检测 BoWRKY10和相关胁迫响应因子的表达量,1周后测定SOD、POD、CAT活性和H2O2含量,每个株系取8株。在抗旱性试验中,每个株系取生长相似的5周龄植物30株,停止浇水14 d后复水,计算存活率。在干旱第7天时,取样测定H2O2含量以及超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性和相对含水量(RWC)、丙二醛(MDA)、脯氨酸、可溶性糖含量。所有测定按李玲的方法[12]进行。每个实验3个重复。

表1 引物序列及用途
1.5 实时定量PCR在烟草中的表达分析
用TRIzol试剂提取总RNA,取2 μg RNA合成cDNA。以表1所列的序列为引物,cDNA 为模板,用ABI7300实时荧光定量PCR系统(ABI,Foster City,CA,USA)进行qPCR,3次重复。反应程序为95 ℃10 s,94 ℃10 s,60 ℃25 s,40个循环。反应结束后分析荧光值变化曲线和融解曲线。每个反应3次重复,采用2-ΔΔCt算法分析结果。
1.6 统计分析
利用Excel 2016和SPSS 17.0中进行数据分析,并从3个独立实验中计算平均值±标准差。两组间比较采用t检验,P<0.05被认为是差异显著。
2 结果与分析
2.1 BoWRKY10基因转化烟草及阳性植株筛选
利用Primer 5.0软件设计带酶切位点的特异性引物扩增目的片段,构建pBIA-35S-BoWRKY10植物表达载体。利用农杆菌介导法将该重组质粒转化到烟草中,100 mg/mL Kan培养基上筛选出长势较好的烟草进行PCR检测。结果表明,在转基因烟草中检测到9个阳性株系。野生型烟草叶片中没有检测到目的条带。qRT-PCR分析表明,9个株系BoWRKY10基因均能过表达,但OE-3、OE-4和OE-6表达量较高(图1)。因此,以下研究选取这3个株系作为研究材料。
2.2 转基因烟草对干旱的耐受性分析
为研究外源BoWRKY10基因表达对植物干旱胁迫响应的影响,选择相对表达量较高的OE-3、OE-4 和OE-6转基因烟草T2代株系进行耐旱性分析,结果(图2-A)显示,转BoWRKY10基因的烟草比WT对干旱处理产生了一定的耐性。对5周龄的转基因植株和野生植株进行干旱处理, 干旱处理14 d后,3个转基因株系叶只有少许萎焉,而野生型烟草完全萎焉,复水后转基因株系恢复较好,转基因株系OE-3、OE-4 和OE-6存活率分别为60.6%、63.3%和61.0%,而野生型对照存活率仅为22.2%,转基因株系存活率显著高于野生型对照(图2-B) 。
2.3 干旱胁迫下转基因烟草种子萌发率及根长
种子在含15% PEG4000 1/2 MS培养基中萌发14 d后,3个转基因株系主根长显著大于野生型 (图3-A,3-B)。在正常1/2 MS培养基上,转基因株系OE-3、OE-4 和OE-6的发芽率和野生型对照没有明显差异(图3-C);而15%PEG4000培养基中,OE-3、OE-4 和OE-6萌发率分别为65.9%、75.4%和84.3%,野生型仅为22.5%(图3-D);与野生型对照相比,转基因株系萌发率分别提高了43.4%、52.9%和 61.8%(图3-D)。
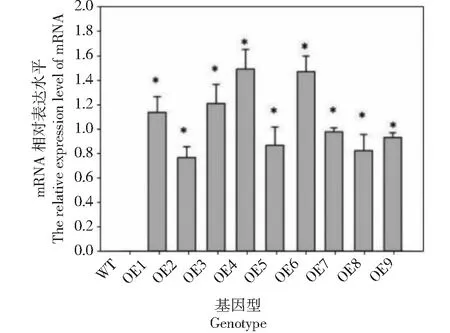
*表示与对照有显著差异(P<0.05),下同* denotes significant differences as compared with the control group(P<0.05), the same as below图1 转基因烟草中BoWRKY10基因过表达分析Fig.1 Overexpression analysis of BoWRKY10 gene in transgenic tobacco
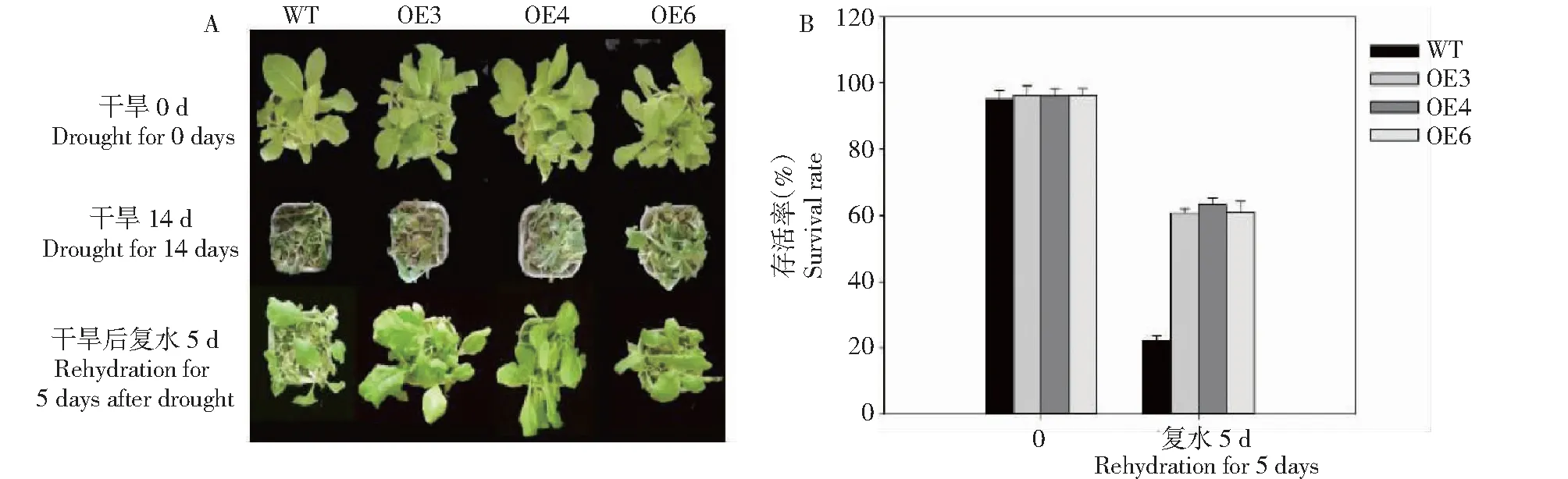
图2 干旱胁迫下转基因烟草的表型和存活率Fig.2 Phenotype and survival rate of transgenic tobacco under drought stress
2.4 干旱胁迫下BoWRKY10过表达对烟草生理特性的影响
转基因植株与野生型(WT)植株在处理前脯氨酸和MDA含量差异不显著。干旱胁迫后,转BoWRKY10基因植株和WT植株的脯氨酸和MDA含量增加,且转BoWRKY10基因植株的脯氨酸含量显著高于WT植株(图4)。相比之下,转基因BoWRKY10株系的MDA含量显著降低。在干旱胁迫后,BoWRKY10转基因植株和WT植株的可溶性糖含量增加,但BoWRKY10转基因植株的可溶性糖含量显著高于WT植株。干旱处理7 d后,BoWRKY10过表达株系的RWC显著高于WT。
干旱胁迫下转基因烟草叶片中SOD、POD、CAT活性显著高于对照,而转基因植株中的H2O2水平较低(图5)。此外,在MS培养基上培养2周的幼苗,移栽到含PEG4000(mol/L,15%)的MS培养基上1周,检测结果与干旱胁迫结果相似。表明,在干旱胁迫下,BoWRKY10的表达通过增加3种重要抗氧化酶的活性影响ROS水平。
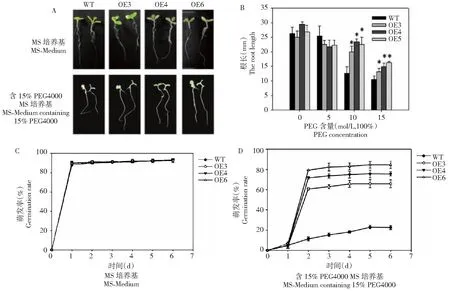
图3 不同胁迫条件下过表达BoWRKY10基因烟草植株的根长和萌发率Fig.3 Root length and germination rate of tobacco plants overexpressing the BoWRKY10 gene under different stress conditions
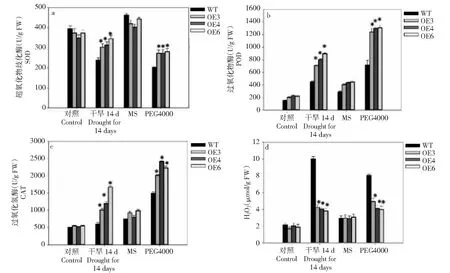
**表示与对照有极显著差异(P<0.01),下同** denotes significant differences as compared with the control group (P<0.01), the same as below图5 正常和干旱渗透胁迫条件下野生型(WT)和转基因烟草中抗氧化酶活性和H2O2积累Fig.5 Three antioxidant enzyme activities and H2O2 accumulation in WT and transgenic tobacco under normal and drought/osmotic conditions
2.5 BoWRKY10调节干旱胁迫下应答基因的表达
以2周龄对照和转基因烟草(OE-3)植株为材料,PEG4000(m/mol/L,15%)处理1周,NtSOD、NtAPX、NtCAT、NtGST、NtSPSAt和NtNCED1等6个ROS相关的胁迫响应基因进行qPCR检测。与对照植物相比,当用PEG4000时,转基因植物中所检测的应答基因均显著上调(图6),表明BoWRKY10在烟草中的表达通过诱导一些ROS相关基因和胁迫响应基因的表达,增强了烟草的抗旱性。
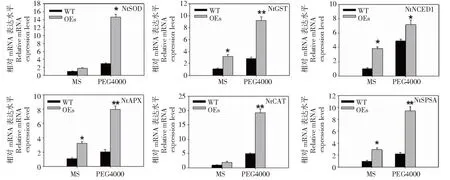
图6 干旱处理下野生型(WT)和 BoWRKY10转基因烟草胁迫响应相关基因表达Fig.6 Expression of stress response genes in WT and BoWRKY10 transgenic tobacco under drought conditions
3 讨 论
WRKY-TFs是植物中最大的转录调控家族之一,具有多种功能环境胁迫下的发育和生理功能。近年来,在水稻[6]、大豆[13]、棉花[14]、短梗草[8]、玉米[15]和小麦[7]等植物中发现了大量的WRKY蛋白。随着研究的深入,发现植物WRKY 转录因子对不同的胁迫环境均有应答。已有研究发现,过表达TaWRKY46增强了转基因拟南芥植株的渗透胁迫耐受性,其转基因拟南芥植株在含甘露醇的1/2 MS培养基中具有较高的发芽率和较长的主根[7]。番茄SlWRKY39是拟南芥AtWRKY40的同源基因,它能被盐、干旱及ABA等相关胁迫因子诱导表达[16]。但Liu等[17]对棉花研究表明,转基因植株中GhWRKY25的过表达降低了抗旱性。本研究表明,当把羽衣甘蓝的BoWRKY10基因克隆到烟草中,其过表达增强了转基因烟草的抗旱性;在干旱胁迫下,转基因株系的根长、发芽率及存活率均显著高于野生型。
在干旱胁迫下,ROS的过量产生会导致氧化损伤,通过激活活性氧清除酶进行解毒[18-19]。MDA浓度是膜脂过氧化的一个参数,反映了响应程度。因此,丙二醛含量通常被用作非生物胁迫下植物受损的参数[20]。抗氧化酶系统,如POD、SOD和CAT酶,通过将ROS保持在阈值水平来保护受胁迫的植物,这些酶的活性表明植物在非生物胁迫下的抗胁迫能力[21-22]。本研究中,与野生型烟草相比,转基因烟草在干旱胁迫下膜损伤程度较低,表现为电解质渗漏、ROS和MDA积累减少,表明活性氧清除系统的增强能力与BoWRKY10过表达植株的抗旱性增加密切相关,即BoWRKY10通过激活ROS清除系统提升烟草抗旱能力。
在干旱胁迫下,一些胁迫相关基因表达水平上调或下调。例如,GmWRKY16的过表达使胁迫相关标记基因RD29A、RD22、KIN1和LEA14的表达显著上调[23]。WRKY17s在其他植物中的负调控也有报道。在盐胁迫下,转基因拟南芥CmWRKY17中AtRD29A和ATDREB2的表达水平显著下调[24]。此外,HaWRKY76的过表达通过调节RAB18、RD29A和RD29B的表达增强了转基因拟南芥的耐旱性[25]。本研究表明,在干旱胁迫下,NtSOD、NtAPX、NtCAT、NtGST、NtSPSAt和NtNCED1等6个基因表达上调。推测BoWRKY10可能通过直接或间接调控干旱胁迫相关基因来调控干旱胁迫。在今后研究中,对BoWRKY10蛋白与DNA结合位点及其BoWRKY10蛋白在其他环境胁迫中功能等方面进行研究,探索其在基因调控中作用机理及其生理功能。
4 结 论
过表达羽衣甘蓝BoWRKY10基因烟草显著提高烟草在干旱胁迫下成活率、萌发率及抗氧化酶活性,增加可溶性糖含量和减少MDA含量;NtSOD、NtAPX、NtCAT、NtGST、NtSPSAt和NtNCED1等6个ROS相关的胁迫响应基因表达上调。本研究表明BoWRKY10基因与干旱胁迫相关。这对培育羽衣甘蓝新品种、探索基因调控机理等具有理论与实践意义。
