典型城市森林旱季土壤团聚体稳定性与微生物胞外酶活性耦合关系
2022-01-07杨洪炳肖以华李明许涵史欣郭晓敏
杨洪炳 ,肖以华*,李明,许涵,史欣,郭晓敏
1. 江西农业大学林学院,江西 南昌 330045;2. 中国林业科学研究院热带林业研究所,广东 广州 510520
随着城市化的迅猛发展,伴随着大气氮沉降、CO2浓度升高、污染物负荷增加、城市热岛效应等对城市森林生态系统产生强烈的干扰(Sala et al.,2000;谢天等,2019)。城市化不仅对城市森林生态系统服务功能(植物生长维持、固碳、自然消减)产生重要影响(Zhao et al.,2016;吕娇等,2020),而且对森林土壤的理化性质、稳定性、微生物特性(谭雪莲等,2019)也产生重大影响。森林生态系统是陆地生态系统中最大的碳库,受气候、植物和微生物等调节(魏书精等,2013)。同时,大气氮沉降、磷富集、酸沉降等对森林生态系统土壤碳循环关键过程有深刻影响(Hansen et al.,2013)。
森林土壤团聚体是其土壤结构构成的基础,土壤团聚体稳定性受土壤发育母质、气候条件、植被类型、植物根系、微生物等影响(王清奎等,2005;刘均阳等,2020)。越来越多研究表明,林分类型的差异会影响其土壤理化性状(吕明亮等,2010),且其林龄对林分土壤物理性质影响不显著,而对林分土壤化学性质影响显著(孙利鹏,2018),因而导致其土壤团聚结构稳定性有所差异(Zoran et al.,2020)。土壤有机碳(SOC)和球囊霉素相关土壤蛋白(GRSP)是土壤团聚体保持稳定性的重要结合剂(Jing et al.,2021),土壤团聚体和土壤微生物是调控土壤有机质分解和积累的重要机制(Lin et al.,2019)。学者们对中国亚热带、暖温带及寒温带典型城市森林土壤团聚体均展开了大量研究,其中林立文等(2020)研究南亚热带人工林土壤团聚体稳定性特征,结果表明:米老排林(Mytilaria laosensisforest)>马尾松林(Pinus massonianaforest)。土壤微生物胞外酶活性反映微生物群落功能,且土壤酶活性易受气候条件、温度、土壤养分限制等影响(Burns et al.,2013;王理德等,2016)。目前,已有研究结果表明秦岭植物生长期的土壤中团聚体(0.25—2 mm)的 β-N-乙酰氨基葡萄糖酶(NAG)酶和微团聚体(<0.25 mm)的β-1, 4-葡萄糖苷酶(BG)酶是影响土壤团聚体稳定性的主要驱动因子(马寰菲等,2021),这方面也不乏有些研究,但城市典型森林土壤团聚体稳定性的主要驱动因子还不得而知。探讨土壤团聚体稳定性特征,其研究内容多集中在雨季和植物生长季,且较多分析团聚体尺度下不同粒径团聚体中土壤和微生物特性空间差异,以及不同粒径团聚体的土壤理化性质和微生物因素对土壤团聚体稳定性的影响已有较多学者深入研究,其初步结果表明微生物酶活性和土壤有机碳对土壤团聚体稳定性差异有较大贡献(吴雪里慧等,2020)。研究区域多集中在中亚热带,而对南亚热带森林土壤团聚体稳定性与酶活性相互作用关系研究较少,且缺乏对城市化背景下影响土壤团聚体稳定性的驱动因素的全面认识。
粤港澳大湾区人口众多,经济蓬勃发展,在国家发展大局中占据重要的战略地位,位于其中的广州市,城市化程度高,亟需对城市森林资源进行合理经营规划。针叶林、针阔混交林与常绿阔叶林这3种森林类型占比最大,为该区域代表性森林类型。因此,以广州市针叶林、针阔混交林、常绿阔叶林3种典型森林类型为研究对象,分析不同森林类型中土壤理化性质、微生物生物量和酶活性及其化学计量比特征,探讨与有机质相关的计算指标对土壤团聚体稳定性的影响,揭示极端干旱气候下城市森林土壤团聚体稳定性与酶活性的耦合关系,以期全面认识城市化背景下森林土壤团聚体稳定性的影响因子,为城市森林规划和经营管理提供理论依据。
1 材料与方法
1.1 研究区概况
研究区域地处粤港澳大湾区中的广州市,样地区 域 经 度 113°23′— 113°50′E , 纬 度 23°11′—23°43′N,属于南亚热带季风气候区。旱雨季分明,一般4—9月为雨季,10—3月为旱季,雨季平均降雨量1462 mm,平均温度26.9 ℃,平均大气湿度80.3%;旱季平均降雨量349 mm,平均温度17.4 ℃,平均大气湿度72.1%。夏季高温多雨,光热充足,冬季低温少雨且干燥。其中 7—9月气温最高,最冷月在1月。土壤发育母质主要为花岗岩,土壤类型为黄红壤,土壤质地为砂壤土,土壤偏酸性。样地其他信息详见表1。

表1 广州市不同森林类型样地信息Table1 Information on sample sites of different forest types in Guangzhou
1.2 土壤样品采集与处理
在广州市选取具有代表性的3种林分类型,分别为针叶林、针阔混交林、常绿阔叶林。在3种森林类型中各自设置3个30 m×40 m样地,每个样地间相隔100 m以上,在每个样地中随机设置3个10 m×10 m样方,样方间最近距离大于4 m,避免边缘效应。
2020年12月,在上述样地的每个样方按“S”型随机选取5个样点,挖取剖面,对表层土(0—10 cm)和剖面土(10—30 cm)进行采样,将同一层土壤充分混合均匀,并剔除可见的枯枝落叶、动物残体、石块等杂质,再将土样分成两部分,一部分过2 mm筛装入样品布袋,另一部分自然风干。过筛的土样一部分存入4 ℃冰箱,用以测定土壤微生物生物量碳氮磷含量、土壤胞外酶活性。自然风干土,主要用来测定土壤团聚体粒径组成、土壤 pH和土壤有机碳、全氮与全磷等。取样的同时,用环刀法(环刀容积为100 cm3)测定土壤容重和含水量。
1.3 测定方法
土壤有机碳(Soil Organic Carbon,SOC)含量采用油浴锅重铬酸钾法测定(鲍士旦,2000),土壤全氮(Total Nitrogen,TN)含量采用凯氏定氮法测定(Sáez-Plaza et al.,2013),全磷(Total Phosphorus,TP)含量测定在紫外可见分光光度计(岛津 UV2405)上完成。土壤 pH测定参考标准《NY/T 1377—2007》,采用电位计法(PHS-3C型),重复3次。土壤微生物量碳氮磷含量采用熏蒸法来测定(Xiao et al.,2016)。在酶标仪(Themo)上,对β-1, 4-葡萄糖苷酶(β-1, 4-glucosidase,BG)、几丁质酶(Chitinase,CHI)、酸性磷酸酶(Acid phosphatase,AP)、过氧化氢酶(Catalase,CAT)活性进行测定,重复3次。
土壤团聚体组成测定参考Li et al.(2019)和吴梦瑶等(2021)方法,并进行一定改动,即取200 g左右土样,测定前挑除石块、细根,将所有土样过8 mm筛,在套筛中振荡5 min,采取干筛法筛取5、2、1、0.5、0.25 mm规格间粒径团聚体,用0.01 g精准度电子秤称取各个粒级的质量,记录数据并计算土壤团聚体粒径组成比例,依此配成50.00 g土样,用以水稳性团聚体的测定。水稳性团聚体实验采用手动湿筛法,即水面浸没土样,超过筛子水面2/3处。振荡前,土样浸泡5 min,然后上下振荡5 min,振荡频率为每分钟30次,拉升高度5 cm,用喷壶将土样转入铝盒,105 ℃烘干至恒质量,记录数据。
1.4 数据处理
在Microsoft Office Excel 2020软件上完成数据的计算处理,进一步利用IBM SPSS 26.0软件对数据进行图表绘制和统计分析,在SmartPLS 3软件上构建土壤团聚体稳定性驱动的最小二乘法路径模型。采用 Kruskal-Wallis秩和检验方法对不同森林类型土壤容重、pH、含水率、SOC、TN、TP含量以及它们之间的化学元素计量比进行显著性差异检验,并采用单因素方差分析对土壤微生物生物量、酶活性及其化学元素计量比进行显著性差异检验,同时采用典型相关分析法分析土壤理化性质、养分、微生物生物量及酶活性的相关关系。显著性水平均设定为α=0.05。对表征土壤团聚体稳定性的指标影响因子采用逐步回归法进行分析。图表中数据均表示为平均值±标准偏差。
为了探讨与有机质相关的一些重要指标与土壤稳定性的关联性,计算微生物熵碳(qMBC)、氮(qMBN)、磷(qMBP),土壤-微生物化学计量不平衡性指标和元素阈值。其中微生物熵碳是微生物量碳与有机碳含量的比值,微生物熵氮是微生物量氮与全氮含量的比值,微生物熵磷是微生物量磷与全磷的比值。土壤-微生物化学计量不平衡性以土壤养分碳氮磷之比与土壤微生物生物量碳氮磷之比的比值来表示(Cimc/Nimc,Cimc/Pimc,Nimc/Pimc)。阈值元素比(马寰菲等,2021)是一个与有效有机质的实际元素组成不同的参数(Sinsabaugh et al.,2010),可用以探讨不同森林类型土壤微生物调节元素利用效率的方式,用此指标(TC꞉N、TC꞉P、TN꞉P)将测定的微生物生物量和生物酶活性联系起来,阐明森林土壤养分限制情况。参考 Sinsabaugh et al.(2009)并进行一定的改动,计算公式如下:
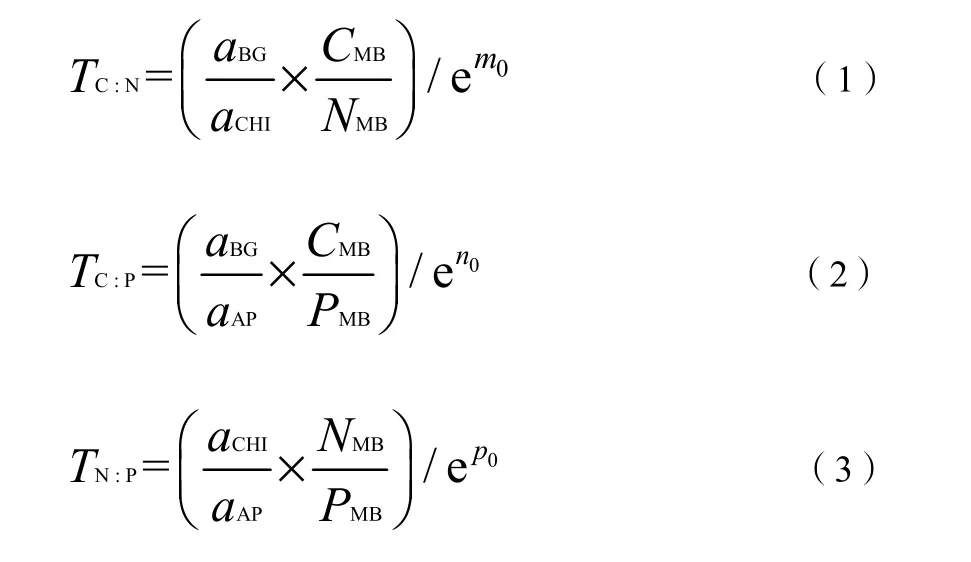
式中:
aBG——β-1, 4-葡萄糖苷酶活性;
aCHI——几丁质酶活性;
aAP——酸性磷酸酶活性;
CMB、NMB、PMB——土壤微生物生物量碳、微生物生物量氮、微生物生物量磷含量;
m0——lnaBG对 lnaCHI的标准主轴回归分析所得到的方程截距;
n0——lnaBG对 lnaAP的标准主轴回归所得方程的截距;
p0——lnaCHI对 lnaAP的标准主轴回归分析得到方程的截距。
在R语言中SMATR程序上完成标准主轴回归分析(Standardized Major Axis Tests,SMA)。
土壤团聚体稳定性特性分为机械稳定性和水稳性,采用平均重量直径(mean weight diameter,DMW)、几何平均直径(geometric mean diameter,DGM)、质量分形维数(fractal dimension of weight,Dm)指标来评价土壤团聚体结构稳定性。参考汪三树等(2013)和Yang et al.(1993),计算公式如下:

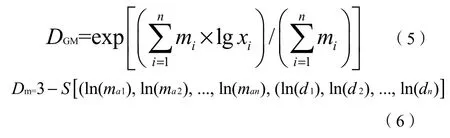
式中:
xi——筛分出来的任一粒径范围团聚体的平均直径;
wi——任一粒径范围团聚体的重量占土壤样品干质量的百分数;
n——土壤样品数量;
mi——各粒级团聚体质量。
∑——求和所有粒径范围的团聚体数量,包括通过最小筛孔的那组团聚体;
mai——各粒级按从小到大排序的团聚体质量进行累加的累计质量;
di——各粒级的平均粒径;
S——进行斜率计算。
2 结果与分析
2.1 不同森林类型土壤理化性质特征
2.1.1 不同森林类型土壤物理性质特征
从表2可知,3种森林类型的土壤容重在表层土中1.31—1.57 g·cm−3,土壤容重在剖面土中为1.47—1.62 g·cm−3。土壤容重随演替阶段的正向发展呈显著递减趋势(P<0.05),且在不同土层内,针阔混交林和常绿阔叶林土壤容重随土层深度增加而显著增加(P<0.05)。3种森林类型表层土含水率为 10.11%—17.6%,剖面土含水率为 10.12%—16.96%。不同演替阶段森林土壤含水率在不同土层中呈现一致的规律,即从演替阶段针叶林到针阔混交林呈下降趋势,从演替阶段针阔混交林到常绿阔叶林呈显著上升趋势,但在同森林类型下,其不同土层土壤间含水率不呈显著性差异。3种森林类型的表层土 pH为 4.01—4.31,剖面土 pH为 4.25—4.45。不同土层土壤 pH随演替阶段的正向发展均有显著上升的趋势,且同森林类型下不同土层间pH均呈显著性差异(P<0.05)。
2.1.2 不同森林类型土壤养分特征
3种森林类型土壤SOC、TN与TP质量分数在表层土中分别为20.11—43.07、1.11—2.09、0.13—0.18 g·kg−1,SOC、TN 和 TP含量在剖面土中分别为 7.12—19.87、0.49—1.11、0.08—0.16 g·kg−1(表2)。在不同演替阶段森林中不同土层间土壤有机碳含量和碳磷比均呈显著性差异(P<0.05),而土壤全氮和全磷含量及碳氮比与氮磷比在针叶林和常绿阔叶林土壤中不同土层间都呈显著性差异(P<0.05)。3种森林类型土壤N/P均值均较低,且C/P值较高,表明区域森林植物生长易受磷养分限制。整体上,不同森林类型各土层间的土壤养分含量及化学计量比均是表层土明显大于剖面土,表明土壤有机碳、全氮和全磷表聚效应明显。

表2 不同演替阶段城市森林土壤理化性质特征Table 2 Physical and chemical properties of urban forest soils at different successional stages
2.1.3 不同森林类型土壤团聚体稳定性特征
3种森林类型的0—10 cm土壤团聚体机械稳定性和水稳性的DMW范围分别为2.499—2.844 mm、1.670—2.091 mm,10—30 cm土壤团聚体机械稳定性和水稳性的DMW范围分别为2.688—3.267 mm、0.877—1.443 mm(图1)。从DMW特征来看,常绿阔叶林的10—30 cm土层土壤团聚体机械稳定性显著高于针阔混交林,而针阔混交林的10—30 cm土层土壤团聚体水稳性高于其他两种森林。3种森林类型的土壤团聚体水稳性均随土层深度增加而降低,而土壤团聚体机械稳定性无明显变化。3种森林类型0—10 cm土层土壤团聚体机械稳定性和水稳性的DGM分别为1.135—1.227、0.901—1.005 mm,10—30 cm土层土壤团聚体机械稳定性和水稳性的DGM分别为 1.211—1.312、0.679—0.852 mm。从DGM特征来看,针阔混交林土壤10—30 cm土层几何平均直径大小显著大于其他2种森林(P<0.05)。3种森林类型0—10 cm和10—30 cm土层土壤机械稳定性的Dm分别为2.359—2.422、2.555—2.635,同样地,土壤团聚体水稳性的 Dm依次为 2.345—2.378、2.653—2.765。数据统计分析得出,不同森林类型中同土层和同森林类型中不同土层土壤团聚体机械稳定性的 Dm之间均无显著性差异,但针叶林和常绿阔叶林中土壤团聚体水稳性的 Dm在土层间均呈显著性差异(P<0.05)。综合DMW、DGM和 Dm指标来看,同森林类型下,仅常绿阔叶林土壤团聚体机械稳定性在土层间呈显著性差异,而 3种森林类型土壤团聚体水稳性在土层间均呈显著性差异(P<0.05)。

图1 不同森林类型中土壤团聚体稳定性指标特征Fig. 1 Characteristics of soil agglomerate stability indicators in different forest types
2.2 不同森林类型土壤酶活性和微生物生物量碳氮磷及其化学计量比特征
2.2.1 不同森林类型土壤酶活性及化学计量比特征
BG和CHI活性在不同森林类型中不同土层间均不具统计学意义上的显著性差异,AP和CAT活性在不同森林类型中不同土层间变化规律基本一致(图2)。AP活性仅在马尾松林和针阔混交林中土壤表层土间呈显著性差异(P<0.05),CAT活性在不同土层间呈现一样的规律,即针叶林与针阔混交林和常绿阔叶林中土壤 CAT活性间呈显著性差异(P<0.05)。马尾松林土壤中BG和CHI活性在不同土层间呈显著性差异(P<0.05),针阔混交林土壤中不同酶活性在土层间均无显著性差异,常绿阔叶林土壤中仅有 CHI活性在土层间呈显著性差异(P<0.05)。在不同森林类型土壤表层土中,各酶活性大小均呈现为PF>BF>MF,而在其剖面土中各酶活性大小无明显差异。

图2 不同森林类型和不同土层酶活性差异Fig. 2 Soil enzyme activities varied among different forest types and different soil layers
由图 3可知aBG/aCHI、aCHI/aAP在 PF和MF土壤各土层间均呈显著性差异(P<0.05)。各土层土壤的aBG/aAP、aBG/aCAT、aCHI/aCAT、aAP/aCAT在不同森林类型之间均无显著差异。除了PF的aBG/aCHI、aBG/aAP与 aBG/aCAT在土层间呈显著性差异(P<0.05),其他在土层间均无明显差异。各土层土壤的aBG/aCHI随森林正向演替有降低的趋势,而其aCHI/aAP随森林正向演替有升高的趋势。

图3 不同森林类型土壤酶活性化学计量比特征Fig. 3 Chemometric ratio characteristics of soil enzyme activity in different forest types
2.2.2 不同土层间微生物胞外酶活性相关性分析
从偏相关分析结果上看(表 3),土壤表层土样品中BG酶活性和AP酶活性间呈极显著线性正相关(r=0.579,P<0.01),CHI酶活性与AP酶活性间呈显著线性正相关(r=0.479,P<0.05),而土壤剖面土样品中所有酶活性之间均不呈现显著线性相关。同种酶活性在不同土层间仅有BG酶活性和AP酶活性是显著正相关的,其他酶活性之间相关性不高(r=0.079—0.244)。CHI活性与BG活性相关性不大,BG和CHI活性与AP活性相关性很大,表明土壤微生物BG和CHI酶活性可能共同促进了AP活性,使微生物更易受磷限制。较剖面土壤酶活性之间的相关性,表层土的土壤酶活性之间呈较强相关关系,微生物获取碳能力的酶(BG)与获取氮的酶(CHI)可能共同促进了获取土壤磷的酶(AP)活性,而剖面土中酶活性之间相关性不大。
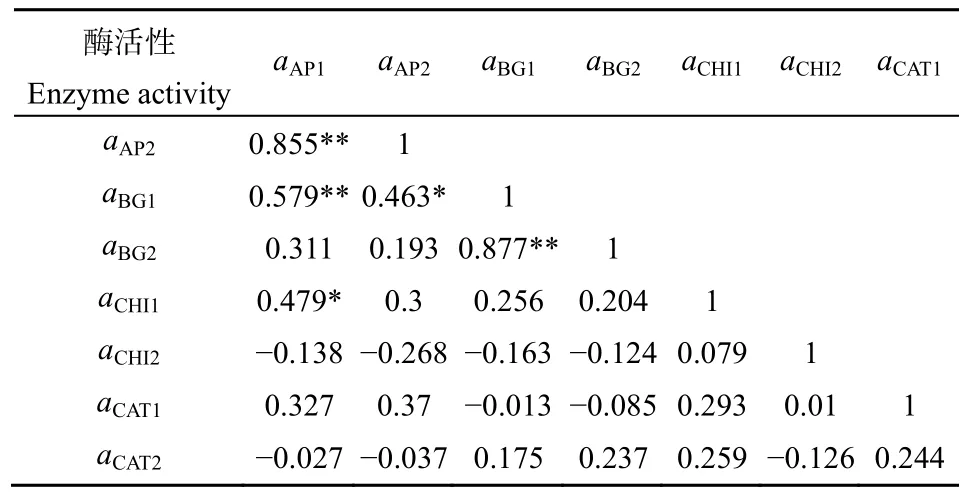
表3 城市森林不同土层土壤胞外酶活性间Pearson相关性Table 3 Pearson correlation between extracellular enzyme activities in soils of different soil layers in urban forests
2.2.3 不同森林类型土壤养分与微生物生物量碳氮磷相关参数特征
从表4可知,在3种森林类型下,微生物生物量碳氮磷含量在同土层间无显著性差异,而同种森林类型下微生物生物量磷含量在不同土层间呈显著性差异。同森林类型下,针叶林和常绿阔叶林中土壤微生物生物量碳氮比和微生物生物量碳磷比在其土层间均呈显著性差异(P<0.05)。针叶林土壤微生物熵碳在其土层间呈显著性差异(P<0.05),常绿阔叶林土壤微生物熵碳氮磷在其土层间均呈显著性差异。针叶林和常绿阔叶林的土壤-微生物化学计量不平衡性指标在土层间均呈显著性差异(P<0.05),表层土的 Cimc/Nimc和 Cimc/Pimc随正向演替而显著增大,而剖面土呈先增大后减小。3种森林类型仅有针阔混交林的 Cimc/Nimc、Cimc/Pimc和Nimc/Pimc在其土层间均无显著性差异,且不同森林类型中同土层土壤的Nimc/Pimc之间也无统计学意义上的差异。

表4 不同森林类型土壤-微生物生物量碳氮磷及化学元素计量比特征Table 4 Characteristics of soil-microbial biomass carbon, nitrogen, phosphorus and chemical elemental stoichiometry ratios in different forest types
元素阈值在不同森林类型土壤中不同土层中呈现不同的变化规律(表 5)。在表层土中,针阔混交林的 TC꞉N数值显著大于其他森林,针叶林的TC꞉P数值显著大于针阔混交林(P<0.05),常绿阔

表5 不同森林类型中不同土层元素阈值比特征Table 5 Characteristics of elemental threshold ratios in different soil layers
叶林的TN꞉P数值显著大于针阔混交林(P<0.05)。而在剖面土中,常绿阔叶林的TC꞉N数值显著大于其他森林(P<0.05),针叶林的TC꞉P数值显著大于其他森林(P<0.05),常绿阔叶林的TN꞉P数值显著大于针叶林(P<0.05)。TC꞉P在同一森林类型中的两土层间呈显著性差异(P<0.05)。
2.2.4 土壤微生物与土壤团聚体稳定性相关性分析
典型相关法结果表明(表6),剖面土中PMB与土壤团聚体机械稳定性特征(DMW和DGM)之间呈显著相关(P<0.05),表层土中土壤资源(SOC、TN与TP)和微生物生物量及酶活性与团聚体机械稳定性特征(DMW、DGM和Dm)相关性较弱。剖面土TP和CMB与土壤团聚体水稳性特征(DMW、DGM和Dm)呈显著相关(P<0.05),而表层土中土壤资源和土壤微生物生物量及酶活性与土壤团聚体水稳性特征(DMW、DGM和Dm)相关性均较弱。
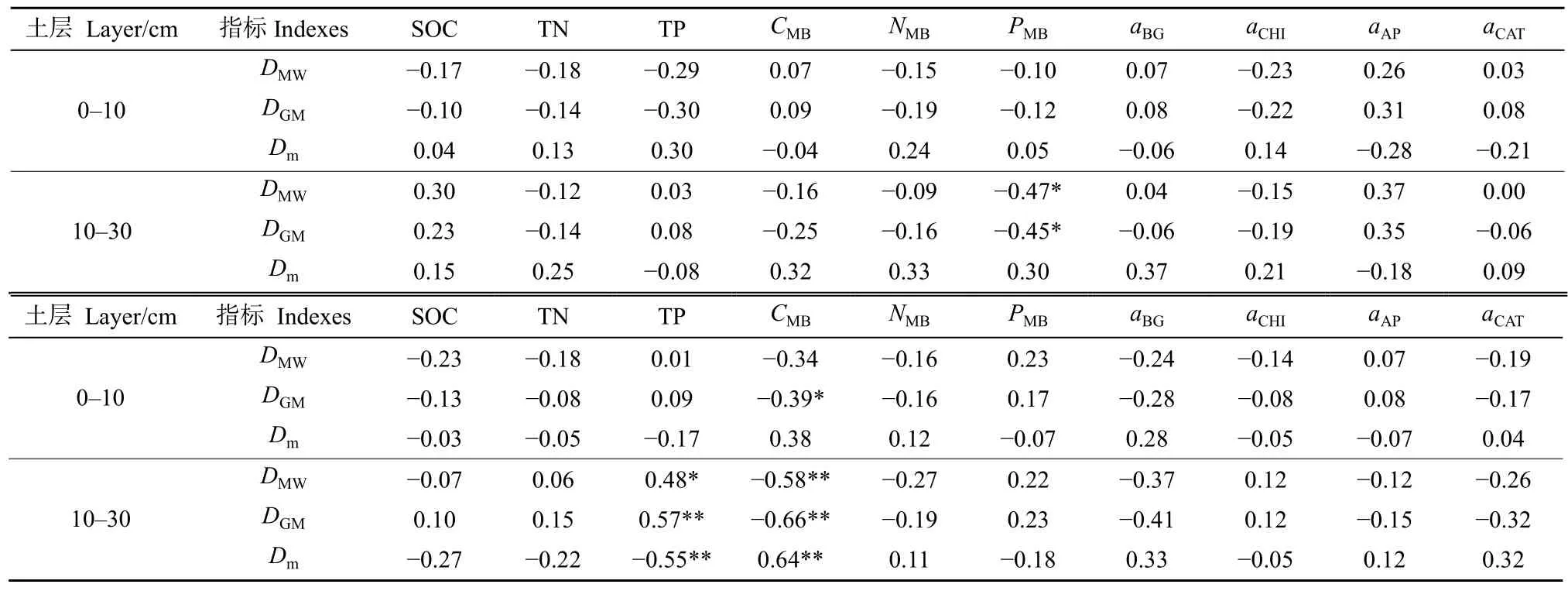
表6 不同土层土壤资源和微生物相关指标与土壤团聚体稳定性指标相关性分析Table 6 Correlation analysis of soil resources and microbial related indicators with soil agglomerate stability indicators in different soil layers
2.3 不同森林类型土壤团聚体稳定性及其影响因素
从路径模型结果来看(图4),土壤理化性质与土壤团聚体稳定性是正关联,但关联程度不高,而微生物熵与土壤团聚体稳定性是负关联的,且关联程度相对较高。此外,微生物生长效率和酶活性与土壤团聚体机械稳定性均是负关联的,与土壤团聚体水稳性均是正关联的,而微生物生物量和元素阈值与土壤团聚体机械稳定性是正关联,与团聚体水稳性是负关联。森林类型和土层的差异解释土壤理化性质变化 63.5%,以及分别解释微生物生物量及酶活性的变化为 71.0%和55.2%,而土壤团聚体机械稳定性整体变化被土壤理化性质、微生物生物量、酶活性等解释的程度为 13.3%,土壤团聚体水稳性整体变化被它们解释的程度为40.0%。
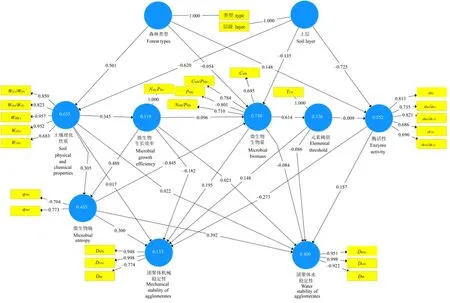
图4 土壤团聚体稳定性驱动因素偏最小二乘法路径模型Fig. 4 A partial least squares path model for the drivers of soil agglomerate stability
从逐步回归分析结果来看(表7),在0—10 cm土层土壤中,对土壤团聚体机械稳定性(DMW)有显著负影响的有微生物熵磷、微生物生物量碳氮磷(R2=0.734,P<0.05),而对土壤团聚体水稳性(DMW)有显著正影响的有C/N元素阈值(TC/N)和显著负影响的微生物熵碳(R2=0.651,P<0.05)。在10—30 cm土层土壤中,对土壤团聚体机械稳定性中的平均重量直径有显著负影响的有微生物熵磷和微生物生物量氮(R2=0.682,P<0.01),对其还有显著正影响的有TC꞉N(P<0.01),微生物生物量碳和pH对土壤团聚体水稳性有显著负影响(R2=0.513,P<0.01)。

表7 土壤团聚体稳定性表征指标影响因子逐步回归模型Table 7 Stepwise regression model of influence factors of soil agglomerate stability characterization index
3 讨论
3.1 不同森林类型土壤理化性质及土壤团聚体稳定性差异分析
3种典型森林土壤容重随正向演替而减小,随土层深度增加而增加,这与其他学者研究结果一致(赵维娜,2016;魏强等,2019;王晓荣等,2020)。土壤含水量随正向演替在土层上呈现一致规律,即针叶林到针阔混交林显著降低,针阔混交林到常绿阔叶林增加,而其不同土层间无显著性差异,这与王晓荣等(2020)研究结果相似。不同土层土壤pH在森林类型间均表现为PF 微生物在土壤有机质的形成与分解中起关键作用,其生物量大小反映土壤有机碳同化与矿化能力大小(Li et al.,2019)。土壤酶活性与土壤养分密切相关(戴凌等,2014),酶活性大小与土壤养分循环速率和微生物利用元素效率关系密切(冯秀秀,2020)。干旱背景下,土壤养分和水分是影响土壤酶活性的主要因子(黄龙等,2021)。土壤微生物生物量碳(CMB)、氮(NMB)、磷(PMB)与对应参与土壤碳氮磷循环的酶活性紧密相关,CMB在不同土层中沿正向演替均是先降低后增加,NMB在不同土层间表现不同规律,PMB在不同土层土壤中是先增加后减小,呈“单峰”状,这可能是CMB、NMB与PMB含量是土壤碳氮磷含量的极少部分,其变化规律同土壤养分变化规律相似,受微生物吸收并转化土壤碳氮磷的能力影响。本研究中土壤微生物群落获取碳相关酶(BG)、获取氮相关酶(CHI)、获取磷相关酶(AP)与清除毒害微生物物质相关酶(CAT)活性随演替正向发展在土壤剖面土间均无显著性差异。值得注意的是,AP和CAT活性在0—10 cm土层土壤上显著呈现PF>MF(图 2),这与戴凌等(2014)对长沙市旱季中不同森林类型土壤酶活性研究结果相反,这可能是由于气候差异以及 pH沿演替方向升高,降低了森林土壤CMB和NMB含量,抑制了部分微生物活性。亚热带森林土壤受磷限制已成共识,其土壤生态酶化学计量比能够反映不同森林土壤之间磷养分限制强度(张星星等,2018)。4种酶活性在不同森林类型间响应规律不同,可能是由于酶活性易受磷限制,以及测定酶活性的方法存在差异。土壤酶活性结果与史丽娟等(2020)关于中亚热带典型马尾松林、湿地松林(Pinus elliottii)、马尾松(Pinus massoniana)-木荷(Schima superba)针阔混交林土壤酶活性研究结果基本相似;但与其关于酶活性化学计量比结果相反,可能由于所选的与N循环相关酶种类存在差异。这说明酶活性化学计量特征受森林类型显著影响,气候因素对酶活性及其化学计量比有较大影响。 从 PLS-SEM 可知,影响森林土壤团聚体稳定性主要是微生物胞外酶活性和微生物熵(图 4),这与马寰菲等(2021)对秦岭不同海拔植物生长期研究结果相似,但本研究发现微生物胞外酶活性对土壤团聚体机械稳定性影响更大,微生物熵对土壤团聚体水稳性影响更大,这也许是由于土壤团聚体对其团聚体内酶活性有物理隔离保护作用(钟晓兰,2015),加之植物根系对土壤团聚体的穿插缠绕作用不同,微生物生存的微环境发生变化,微生物胞外酶活性可能影响土壤碳氮化学转化过程与土壤微生物量以及多糖、有机质等胶结物质的分解,进而影响土壤团聚体的形成与稳定。从土壤团聚体稳定性的逐步回归分析结果(DMW)来看,0—10 cm和10—30 cm土层受环境因子影响有所不同,但两土层土壤团聚体稳定性均受到微生物生物量与微生物熵的负影响,同时土壤团聚体稳定性受土壤微生物对养分需求(元素阈值)的正影响。孙利鹏(2018)研究北温带半湿润半干旱地区天然次生林,结果表明表层土壤团聚体稳定性主要受微生物氮和碳含量等因素影响,本研究结果与其研究结果相似,但本研究探讨了两种团聚体稳定性特征(表7),结果表明其团聚体的水稳定性还受酸性磷酸酶活性和pH影响。胡琛等(2020)研究表明,林分是土壤胞外酶活性及其生态化学计量比的重要影响因素,其对土壤有机碳和pH值的影响是调控土壤酶活性变化的主要因素。因此,森林类型对土壤酶活性有重要影响,土壤物理因素和微生物酶活性可影响土壤团聚体稳定性。不同森林类型土壤团聚体稳定性(机械稳定性和水稳性)在0—10 cm土层深度无显著差异,可能是由于自然因素和人为因素对土壤质量的影响主要集中在表层土上,加之城市化趋同效应引起的。 土壤团聚体稳定特性在不同土层中所受影响有所差异,或许可以从以下几点来解释,(1)土壤有机碳组成和团聚体结构中有机碳量受到气候变化的显著影响(Su et al.,2020;孙筱璐等,2018),土壤团聚体力学性质发生变化,且水稳性团聚体分布受影响较大。(2)根据生态化学计量学理论(Ecological Stoichiometry,EST),森林类型和土层对土壤养分元素限制影响程度不同,微生物可能受元素限制的影响,进而调节元素利用效率。较高的元素阈值TC꞉N数值伴随着更高的C利用效率和更低的N利用效率,较高TC꞉P数值伴随着更高的C利用效率和更低的P利用效率等(Sinsabaugh et al.,2013;Agumas et al.,2021)。本研究结果表明:针阔混交林在表层土中有更高的的 C利用效率和更低的N利用效率,而在剖面土中有更低的C利用效率和更高的N利用效率,且在两土层中有更高的P利用效率。常绿阔叶林在两土层中均有更高的N利用效率和更低的P利用效率,表明针阔混交林较其他两种森林,微生物生物量中有相对较少的 CO2排放和更多的碳固持,以及相对较高的对磷元素的有效固持和较少的磷转化。事实证明,微生物 C利用效率对SOC有非常大影响(Wang et al.,2021),其间接影响了不同森林类型土壤团聚体特征,这与微生物活动受P限制条件相呼应,说明微生物群落通过调整元素阈值也可以影响土壤团聚体稳定性。(3)P限制。亚热带森林在氮沉降背景下土壤和酶活性更易受磷限制,且其土壤P的有效性较低(王全成等,2021)。Jian et al.(2021)发现较低的酶促 C꞉P[ln(BG)꞉ln(AP)]和酶促N꞉/P[ln(NAG)꞉ln(AP)]的获取率表明了P限制,亚热带人工松林可能受到强烈的P限制。本研究区在N沉降背景下将更突显P的限制作用,且不同森林类型中的不同土层土壤微生物酶活性对磷的限制响应不同,以对土壤碳固持和碳循环过程产生不同的作用效应。在P限制地区,今后可选择P分配能力更强的树种进行栽培,或通过基因技术来选育P利用效率更高的树种,以降低P对植物和微生物的限制强度,提升森林土壤质量,并改善土壤结构。 (1)广州市不同典型森林中土层间土壤团聚体的质量分形维数(Dm)随正向演替均无明显变化,而土壤团聚体机械稳定性的平均重量直径(DMW)、几何平均直径(DGM)在表层土中随正向演替而降低,在剖面土中基本呈升高趋势。土壤团聚体水稳定性的DMW、DGM在两土层中均随正向演替而先升高后降低,其在演替阶段和土层间的变化趋势主要受微生物生物量和微生物胞外酶活性的影响。 (2)微生物通过调控微生物生物量和调节碳氮元素利用效率,可使演替后期受磷限制状况得以改善,进而对不同演替阶段森林旱季土壤团聚体的不同稳定性特性产生影响。 (3)土壤团聚体机械稳定性的主要影响因子有微生物熵磷(qMBP)和微生物生物量氮(NMB),土壤团聚体水稳定性的主要影响因子有酸性磷酸酶活性(aAP)和微生物生物量碳(CMB)。土壤微生物熵和微生物胞外酶活性是影响土壤团聚体稳定性的主要驱动因子。3.2 不同森林类型土壤微生物生物量与酶活性及化学计量比特征
3.3 影响土壤团聚体稳定性因素分析
4 结论
