不同林龄杉木人工林根际效应及养分特征1)
2022-01-06黄章翰李维扬纪小芳冯明崔莉娜鲁建兵姜姜刘光正
黄章翰 李维扬 纪小芳 冯明 崔莉娜 鲁建兵 姜姜刘光正
(江苏省水土保持与生态修复重点实验室(南京林业大学),南京,210037)(江西省林业科学研究院)
根际是指受植物根系影响数毫米范围内的土壤区域[1],也是土壤—植物根系—微生物3者间物质转化与能量流动的场所[2],对生态系统的养分动态分布与循环起着重要作用[3-4]。植物通过根系分泌物的输入,以及根表皮脱落物的裂解,改变根际微生物酶的数量与活性[5],加之根系吸收的影响,使根际养分状况在浓度、形态与分布等方面发生变化[6-7],即存在明显的“根际效应”[5],由此深入认识根际生态过程对养分分配的影响具有重要意义。
人工林经营过程中,根际与非根际土壤特性的差异决定了根际养分特征[8],直接影响植物的生长状态。随着人工林林龄的增加,林分结构、凋落物形成与分解、根系周转与代谢都会发生相应的变化[9-10],从而影响根际周围的养分循环过程。不同养分的根际效应随林龄变化往往表现出不一致的变化规律。叶功富等[11]研究木麻黄人工林土壤有机质和全氮的根际效应随着的发育先减少后增加,马尾松人工林的建立不改变土壤氮根际效应的变化趋势,但有机质的根际效应随林龄的增加显著增加,可能受到光合作用的限制[12],孙涛等[13]发现由于根际微生物种类与数量的影响,樟子松林地土壤有效养分的根际效应对林龄的响应趋势明显不同于土壤全量养分。因此,对人工林根际效应随林龄变化的规律并无一致的结论,其机理仍不清楚。
杉木(Cunninghamialanceolata)是我国南方的速生针叶用材树种,因其生长周期短、经济价值高等特点而被广泛栽植,全国杉木人工林栽培面积达到1.7×107hm2,占全国人工林面积的24%[14]。随着栽植面积扩大、多代连载,以及不合理的经营措施,杉木林出现了土壤地力衰退、初级生产力下降等诸多问题[15-16],尽管已有多名专家学者对此展开研究,目前仍尚未弄清杉木人工林地力退化的内在机制,从根际效应角度研究不同生长阶段杉木人工林土壤养分循环特征,可为杉木人工林可持续经营提供新思路。目前,杉木人工林根际养分研究集中在根际与非根际土壤养分含量及有效性的对比[17]、土壤养分计量比特征[18]以及营林措施对杉木林根际养分的影响[19],尚未结合土壤养分与酶活性之间的关系,深入探讨不同生长阶段植物根际对土壤养分与酶活性的潜在影响。有鉴于此,本文以江西省官山林场10、25、45年生杉木人工林为研究对象,分析不同林龄杉木人工林根际与非根际土壤养分及酶活性状况,以期揭示林龄背景下杉木根际对土壤养分与酶活性的作用关系,为杉木人工林的土壤养分管理提供科学依据。
1 研究区概况
试验研究区位于江西省中部吉安市永丰县官山林场(115°17′~115°56′E,26°38′~27°32′N),属中亚热带湿润气候,年均气温18 ℃,年均降水量1 627.3 mm,年均无霜期279 d。主要土壤类型为红壤。官山林场自然条件优越,适宜松、杉、竹及多种阔叶树生长,是江西省的林业重点产区[20]。场内杉木面积分布广泛,其它植物种主要有湿地松(Pinuselliottii)、毛竹(Phyllostachysedulis)、楠木(Phoebebournei)、枫香(Liquidambarformosana)等。
2 材料与方法
土壤样品于2018年7月在江西省永丰官山林场中采集,分别选取10、25、45年生3种林龄的杉木人工林样地,样地土壤类型、立地条件一致,杉木种源一致,林分基本状况见表1。每个样地设置3个20 m×20 m的样方,在每个样方内随机选择3株杉木,取其根际与非根际土,分别混合均匀,装于无菌袋中,置于便携式冰箱内。具体方法:根际土壤(RS)取样采用抖落法,去除土壤表面的动植物残体,顺着树干基部挖开根系集中分布的土层(0~15 cm),轻抖去大块不含根系的土壤,用小毛刷将黏附在细根表面上的土壤收集至无菌袋中,为根际土[21];非根际土壤(BS)取样在离树干基部50 cm外进行,在每个样方内随机挖取3个0~15 cm深土壤混匀,为非根际土[11]。将土样带回实验室后风干过2 mm筛,用于土壤养分和酶活性的测定。

表1 样地基本特征
2.1 测定方法
土壤有机碳(SOC)采用重铬酸钾氧化法测定;活性有机碳(LOC)采用高锰酸钾氧化比色法测定;全氮(TN)采用元素分析仪测定;速效氮(AN)采用碱解扩散法测定;全磷(TP)采用酸溶-钼锑抗比色法测定;速效磷(AP)采用氟化铵-盐酸浸提法测定;土壤的pH采用电位法测定[22]。
土壤蔗糖酶采用3,5—二硝基水杨酸比色法测定,酶活性以37 ℃下培养24 h后1 g土消耗葡萄糖的毫克数表示[23];脲酶采用水杨酸钠—二氯异氰尿酸钠比色法测定,酶活性以37 ℃下培养2 h后1 g土释放铵态氮的微克数表示[24];酸性磷酸酶采用对硝基磷酸苯二钠比色法测定,酶活性以37 ℃下培养1 h后1 g土水解生成对硝基苯酚的毫克数表示[25]。
2.2 数据处理
土壤养分与酶活性的根际效应(ER)采用如下公式进行计算[26]:
ER=[(Crs-Cbs)/Cbs]×100%。
式中:Crs和Cbs分别表示土壤养分与酶活性指标在植物根际与非根际土壤中的数值大小。当ER>0时表现为根际正效应,ER<0时表现为根际负效应。
土壤养分有效性的计算,本文将活性有机碳与惰性有机碳(土壤有机碳与活性有机碳的差值)比值作为土壤有机碳活度,即表示土壤有机碳的高活性[27];将速效氮与全氮比值和速效磷与全磷比值的百分率用于表示土壤氮、磷养分的有效性[28-29]。
运用SPSS25.0软件对数据(平均值±标准差)进行统计分析。用双因素方差分析和最小显著差异法(α=0.05)比较各个变量的差异显著性。运用线性回归拟合分析土壤养分与酶活性之间的关系。
3 结果与分析
3.1 不同林龄土壤养分的根际效应
由表2和表3可知,除土壤氮外,不同林龄杉木根际与非根际土壤养分质量分数呈一致的变化规律,其中根际土壤养分分质量分数均高于非根际土壤,表现出在根际的富集现象。由于杉木根系对不同养分吸收速率的不同,土壤碳、氮、磷的根际效应随林龄变化表现出不同的趋势。根际与非根际土壤中,SOC和LOC量均随林龄增加显著增加,其根际效应都表现为先增加后减少的趋势,后期趋于稳定,SOC的根际效应强度(30%~60%)大于LOC(10%~20%);随林龄增加,根际土壤TN和AN量先增加后减少,而非根际土壤TN和AN量显著增加,土壤氮的根际效应亦呈现先增加后减少的趋势,但后期明显下降,根际效应对TN和AN的影响一致(均在20%~60%);根际与非根际土壤的TP和AP量总体表现为随林龄增加而增加的趋势,其根际效应持续增强,但TP的根际效应(100%~150%)远大于AP(10%~20%)。
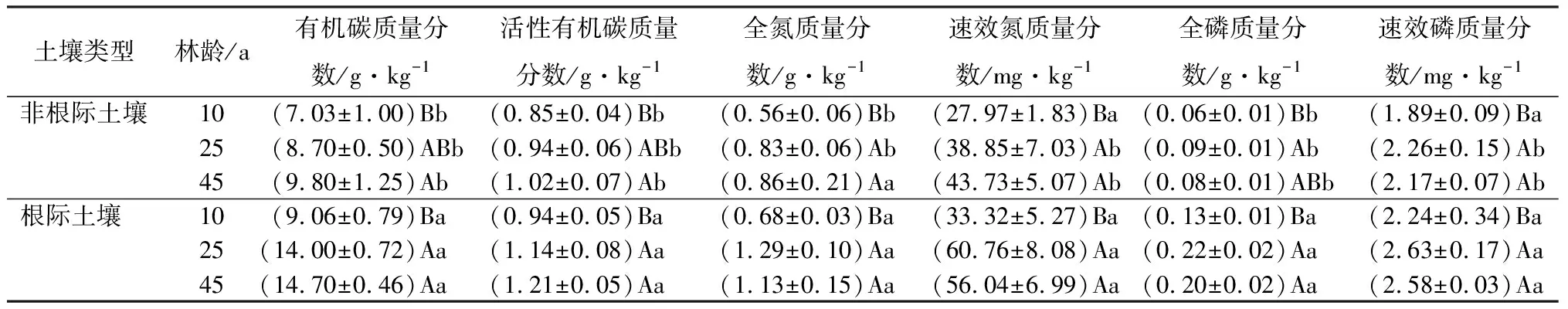
表2 不同林龄杉木人工林土壤养分质量分数

表3 不同林龄杉木人工林土壤养分根际效应
表4可以看出,根际SOC活度随林龄增加而显著减少,且与非根际间的差异逐年显著,说明根际最终促进了ROC的积累;林龄与根际对土壤氮有效性无显著影响;土壤磷有效性随林龄变化趋势与SOC活度相同,且根际土壤磷有效性均显著低于非根际土壤,表明杉木根系倾向于吸收AP,导致AP的根际效应表现最小,杉木生长可能受磷限制;土壤w(C)∶w(N)∶w(P)生态计量比在3个林龄组中均无显著差异,但根际对w(C)∶w(P)与w(N)∶w(P)具有显著作用;此外,杉木土壤整体呈现酸性,随着林龄增加,酸性增强,且根际土壤pH均显著低于非根际土壤。
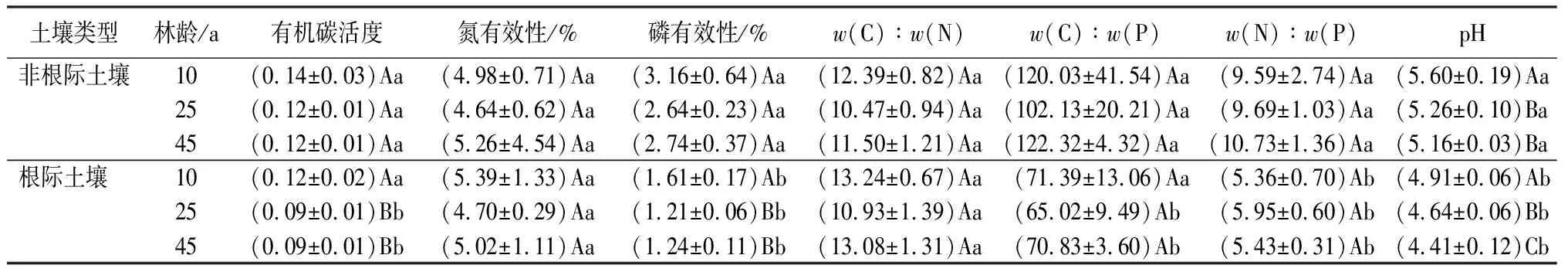
表4 不同林龄杉木人工林土壤养分有效性与化学计量比
3.2 不同林龄土壤酶活性的根际效应
土壤酶主要来源于植物根系和微生物活动,并参与土壤养分循环过程。由表5和表6可知,土壤碳、氮、磷转化酶(蔗糖酶、脲酶、酸性磷酸酶)随林龄变化的规律趋同于相应养分,不同林龄根际土壤酶活性均高于非根际土壤。其中,蔗糖酶与酸性磷酸酶的根际效应均随林龄增加而增加至25 a后趋于稳定(分别为15%~30%和50%~70%),脲酶的根际效应则呈现与土壤氮相同变化趋势,范围在150%~250%。
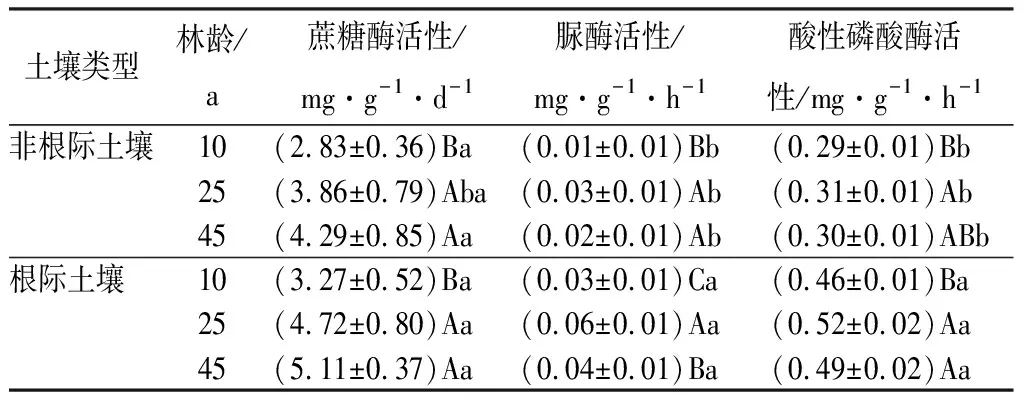
表5 不同林龄杉木人工林土壤酶活性

表6 不同林龄杉木人工林土壤酶根际效应
3.3 土壤酶活性对养分及根际效应的影响
由图1可知,土壤碳、氮、磷养分质量分数均与相应酶活性(蔗糖酶、脲酶、酸性磷酸酶)呈极显著正相关,且在根际土壤中的关系更为明显。故进而分析根际酶活性对养分根际效应的影响发现(图2),除TP外,土壤养分的根际效应均与根际酶活性呈显著正相关,说明根际对养分的富集是通过促进根际土壤酶分泌实现的,而TP的根际效应与酸性磷酸酶活性关系不显著,表明存在其它复杂的关系对根际TP积累产生影响。

*表示P<0.05的显著性;**表示P<0.01的显著水平。
4 结论与讨论
4.1 不同林龄杉木人工林养分根际效应的变化
研究发现,SOC和LOC的根际效应表现为正效应(ER>0),根际促进了土壤碳积累,随林龄增加根际效应先增加后减少,这与刘顺等[17]对陈山红心杉的研究结果相似,分析原因,杉木前期处于速生阶段,土壤呼吸速率大,以CO2形式释放到大气中的碳量多,而补充其来源的凋落物分解相对较慢[18],有机碳的消耗大于归还,从而造成10 a杉木土壤有机碳含量最低。随着林龄增加,林下生态系统逐渐完善[14],加速凋落物的分解速率,输入土壤有机碳的含量也随之增加,但不同于非根际土壤,植物除通过细根周转向根际土壤输入碳外,其根系在生长期会向根际分泌各种次生代谢物质,其中90%作为LOC化合物[30],为土壤微生物提供丰富的碳源,以提高微生物胞外酶活性及其对根际碳的固定能力[31],导致土壤碳的根际效应逐年增强。当林分成熟后,杉木生长相对变慢,根际的碳转化酶活性缓慢增加,根际养分积累速率下降,促使杉木SOC和LOC的根际效应呈现减少趋势。另一方面,虽然根际效应增加LOC量,但因其不稳定性,能在短时间内迅速分解,导致LOC随林龄增加速度小于ROC的增加速度,最终促进ROC在根际的积累,因此SOC的根际效应强度(30%~60%)显著大于LOC(10%~20%)。Liu et al.[32]研究发现,根际菌丝的胶结作用有利于土壤团聚体形成,进而促进根际对土壤碳的贮存。

*表示P<0.05的显著性。
土壤氮质量分数由凋落物归还和植物吸收之间的平衡所决定,其根际效应随林龄增加,表现先增加后减少趋势,根际效应对TN和AN的影响一致(均在20%~60%),分析原因可能是由于植物根际不断从外界摄取营养元素,氮养分在土壤中向根际迁移速度较快[8],根际氮输入速率大于非根际,造成TN的根际效应显著增加,同时植物根系数量与微生物量增加[33],根际氮的周转速率加快,底物TN的富集引起脲酶的根际效应大幅度增强(150%~250%),从而维持土壤TN向AN的转化[34],因此TN和AN对根际效应的响应具有趋同性。然而,相比于同纬度江西千烟洲亚热带人工林土壤氮水平的0.86 g/kg[35],研究区25、45 a杉木根际氮质量分数(1.28、1.13 g/kg)均高于千烟洲人工林,表明杉木25 a后对土壤氮养分的需求趋于稳定,根际氮富集可能通过底物反馈抑制脲酶活性,减缓凋落物分解与有机氮矿化过程[36-37],导致根际对土壤氮的富集效应明显降低,但该解释缺乏直接证据。另外,有研究报道,南方针叶树种在生长发育过程中会出现氮素流失的现象[9-10],与本研究结果不符,这可能和不同研究区的林分、气候、土壤类型等因素有关[10,38],本研究区林分结构一致,土壤氮有效性稳定,根际诱导脲酶活性的提高,增加对土壤氮的固持作用。
在过去的研究中,土壤磷根际效应随林龄变化结果并不一致,刘顺等[17]研究发现,随林龄增加,陈山红心杉土壤磷的根际效应长期表现为负效应(ER<0),而本文土壤磷的根际效应为正效应(ER>0)结果与杨阳等[39]对荒漠草原区优势种的研究结果一致,根际效应持续增强,分析原因主要是本研究区土壤平均磷水平为0.13 g/kg,与同纬度江西千烟洲人工林土壤的0.11 g/kg相近[35],同属低纬度缺磷地区,杉木为了适应低磷胁迫,根系会分泌大量质子和有机酸,以促进难溶性矿物磷的分解与吸收[40],提高土壤磷质量分数的同时,造成根际pH值显著低于非根际,另一方面,随着林龄增加,凋落物分解产生的酸性物质增加,土壤愈加酸化,促使酸性磷酸酶活性显著增强[41],有效促进土壤有机磷转化为无机磷[42],以供植物吸收利用,在pH值和酸性磷酸酶的协同作用下,土壤磷的根际效应随林龄增加而稳定增强。此外,根际对TP的富集效应(100%~150%)远大于AP(10%~20%),分析原因,尽管凋落物分解与有机酸淋溶增加了土壤磷质量分数,然而,当土壤磷有效性低于2.0%,则说明磷素利用率低[29,43],TP难以转化为AP,本研究中根际土壤磷有效性随林龄增加显著减少,且均低于2.0%,表明该地区磷有效性低,生态系统长期缺磷,产生的AP能被植物快速吸收,导致AP的根际效应表现最小。
土壤化学计量特征反映土壤有机质组成和营养供应状况,是表征土壤内部碳、氮、磷元素循环的重要指标[44]。本研究土壤w(C)∶w(N)=11.93均值与全国土壤平均值(11.90)[10]相当,不同林龄根际与非根际土壤w(C)∶w(N)变化趋势不显著,表明碳、氮作为结构性成分,紧密相关,二者在积累和消耗过程中存在同步性关系,验证了不同生态系统土壤C∶N相对稳定的结果[45]。土壤w(C)∶w(N)=91.95与w(N)∶w(P)=7.79均高于全国平均水平(61.00、5.20)[10],说明该地区微生物在有机质矿化过程中的磷释放较差,杉木磷受限可能加剧,另外较高的w(C)∶w(N)与w(N)∶w(P)也反映了磷有效性越低的问题[44],因此,在杉木人工林经营过程中应适当增施磷肥,提高磷素活化能力,维持土壤与植物的良性养分循环。
4.2 不同林龄杉木人工林根际效应与酶活性的关系
本研究发现,林龄是影响杉木人工林生长状态的重要因素,杉木不同生长阶段根际与非根际土壤养分状况不同,分析原因主要是不同发育阶段植物的吸收与归还不同,以及根际与非根际间养分富集速率的差异所造成的。相关分析结果显示,土壤碳、氮、磷含量均与相应酶活性呈极显著正相关,与前人的研究结果相似,徐秋芳等[31]研究证明土壤蔗糖酶与有机碳关系最为密切,其活性表征了土壤有机碳的累积和释放易溶性碳素的能力,刘淑英等[34]人的研究中,土壤脲酶与TN、AN相关关系显著,与本研究结果一致,这反映了脲酶将土壤有机氮水解转化为有效态氮的作用,是植物氮源的直接驱动力,另外研究表明,酸性磷酸酶的酶促反应能释放土壤中可供植物吸收的无机磷[46],尤其是林木生长受到磷限制时,其活性通常会维持在较高水平[47],以利于提高土壤磷有效性,保证林木正常生长。除TP外,土壤养分的根际效应均与根际酶活性呈显著正相关,说明在典型的低效林区中,根际酶的分泌是调控根际促进养分积累的关键,值得注意的是,酸性磷酸酶对土壤磷的促进作用主要体现在AP方面,尽管TP含量与酸性磷酸酶活性关系密切(P<0.01),但TP根际效应与酸性磷酸酶活性呈微弱的正相关,TP随林龄的根际富集速度大于酸性磷酸酶活性的增加速度,表明存在其他因素对根际TP积累产生影响,崔莉娜等[48]人的研究证明,杉木根际存在一定的丛枝菌根真菌,通过侵染植物根系,促进土壤磷聚集和作物吸收,李蓉等[49]人的研究中发现,由于杉木长期处于低磷环境,根际中的溶磷细菌含量较高,补充土壤磷素的同时产生致酸物质,降低土壤pH值,促使酸性磷酸酶活性进一步提升,协同改善杉木土壤磷环境。具体而言,10年生杉木处于生长旺盛期,对养分需求量大,此时林分相对稀疏,凋落物归还养分不足以被植物吸收,导致土壤养分出现盈亏,由此,养分根际效应表现较低;25年生杉木林分逐渐郁闭,林内竞争较大,此时的杉木根系汲取养分的范围更加宽广,使得土壤养分向根际发生迁移,同时根际酶活性显著高于非根际,根际养分输入速率大于非根际,造成此时的养分根际效应显著增强;45年生杉木人工林处于成熟林阶段,木材生长相对缓慢,对养分的需求量相对减少,根际碳积累速率下降,造成此时的土壤碳根际效应减弱,而该阶段土壤氮根际效应减弱与根际脲酶活性降低有关,不同于土壤碳、氮,此时的杉木仍处于磷匮乏状态,根系优先吸收AP,导致土壤磷有效性较低,需通过刺激微生物分泌更多酸性磷酸酶,以提高根际对土壤磷的促进效益。
本研究得出林龄对土壤养分和酶活性具有显著影响,仅蔗糖酶活性在根际与非根际间无显著差异,随林龄增加,根际与非根际土壤养分质量分数、酶活性均呈增加趋势,土壤酸性增强;在林龄和根际共同影响下,SOC和LOC的根际效应随林龄增加呈现先增加后减少的趋势,二者均与蔗糖酶活性显著相关,根际土壤ROC比例大于非根际;TN和AN的根际效应25 a最大,与脲酶活性变化规律一致,造成45 a杉木根际对土壤氮的富集效应减弱;TP和AP的根际效应随林龄增加而增加,但TP的根际效应远大于AP,酸性磷酸酶活性与AP根际效应显著相关。因此,在杉木人工林经营管理中,应长期注意磷养分的添加管理。根际对土壤养分和酶活性的具体影响机理还需结合根际微生物状况深入研究。
