纤维素降解菌的筛选和培养条件的优化
2022-01-04陈诗瑶劳艳婷
陈诗瑶, 劳艳婷, 谢 苗
(1.福建农林大学生命科学学院, 福建福州350002;2.闽台作物有害生物生态防控国家重点实验室, 福建福州350002)
纤维素是植物细胞壁的重要组成成分,在大自然中分布很广,在食品、医药、建筑、废水处理以及造纸等行业中有着非常广泛的应用(罗成成等,2015)。 但作为一种重要的资源和能源,广泛存在的纤维素并没有得到有效地处理或利用,大部分都被直接焚烧或丢弃,这样的处理方式给环境和生态平衡带来了极大的破坏(张瀚罡,2019)。 因此,如何高效、无害地降解以及利用环境中的纤维素成为目前急需解决的问题。
目前对纤维素的主要处理方法有物理法、化学法和生物学方法,但物理法和化学法工艺复杂,成本较高且有一定的环境污染,所以在实际应用中有所限制。 自然界中存在众多高产纤维素酶的微生物,这些微生物能将自身产生的纤维素酶分泌至胞外且易于纯化,且对纤维素降解有较强的专一性(王盼星等,2018)。 此外,纤维素降解菌对环境的危害小,能实现对资源的循环利用(佟硕秋等,2020;文少白等,2010)。 因此,利用微生物降解是当前降解纤维素最有效、最经济、最环保、也是最热门的方法(乔健敏等,2019)。 自20 世纪以来,国内外已经报道了数千个产纤维素酶的菌株(贾丙志等,2008),如小刺青霉菌(简立燕等,2017)、多粘芽孢杆菌(Gorskaet al,2001)、泡盛曲霉(邢力等,2019)等。 尽管目前在土壤(吴越等,2018)、植物(顾挺等,2011)、动物(李君风等,2017)和粪便(张智等,2017)等内都有获得纤维素降解菌的相关报道,同时纤维素的生物学处理方法也得到了广泛关注,但由于其产酶数量及产酶活力都没有达到工业应用的要求、水解液中的产物会抑制微生物的发酵结果、部分菌株的耐受能力较低等原因,目前还难以运用到实际生产中(Daret al,2018;马泽林等,2017)。 因此,人们仍需寻找更稳定、更高效的纤维素降解酶来源。
厨余垃圾内有机物含量高、营养元素丰富,容易滋养病原菌等有害微生物,因此厨余垃圾的不当处理,会对环境造成严重污染(徐锐,2014)。 目前,对厨余垃圾的主要处理方法有焚烧、填埋、粉碎直排等非生物处理方法,好氧堆肥是主要的生物处理方法。 大部分的非生物处理方法经济效益低,容易造成资源浪费且严重破坏生态环境(孙孟喜,2017)。 而采用生物处理法是较为简单、易行、环保的方法(吴昊等,2011)。 研究表明,厨余垃圾内纤维素含量较高(郑旴等,2016),因此可能富含纤维素降解菌。 本研究通过刚果红水解圈直径初筛和测定酶活力等指标复筛,从经半年发酵的厨余垃圾中筛选出高效的纤维素降解菌,并使用响应面法优化纤维素降解菌的培养条件,分析了不同碳源浓度、培养时间和温度对菌株产酶活力的影响,以期得出菌株的最佳培养环境。
1 材料与方法
1.1 试验材料
1.1.1 样品 福州某公司提供的已发酵6 个月的厨余垃圾。
1.1.2 培养基 羧甲基纤维素钠(CMC-Na)固体培养基:0.5 g KCl,0.5 g MgSO4,1.0 g K2HPO4,3.0 g NaNO3,5.0 g CMC-Na 和 20.0 g 琼脂,溶于 1 000 mL 蒸馏水;CMC-Na 液体培养基:0.5 g MgSO4,3.0 g NaNO3,1.0 g K2HPO4,0.5 g KCl 和 5.0 g CMC-Na,溶于 1 000 mL 蒸馏水;LB 液体培养基:5.0 g 酵母提取物,5.0 g NaCl 和 10.0 g 蛋白胨,溶于 1 000 mL 蒸馏水,pH调至7.4;以上培养基均121 ℃高压灭菌20 min。
1.2 试验方法
1.2.1 纤维素降解菌的培养及分离纯化 取10.0 g 样品加入90 mL 无菌水中,振荡30 min。运用梯度稀释法制备稀释度为10-1~10-5的样品稀释液。 在CMC-Na 固体培养基中分别进行平板涂布,28 ℃培养72 h。 使用三区划线法从稀释度为10-3~10-5且优势菌落分布均匀的平板中分离获取形态大小不一的单菌株,并接种于LB 液体培养基制成菌液并保存。
1.2.2 纤维素降解菌的初筛 从上述菌液中各取2.5 μL 分别点种于CMC-Na 固体培养基中心,28 ℃培养72 h,重复3 次。 培养完毕后加入适量1 g·L-1刚果红,均匀覆盖培养基表面染色30 min 后,用1 mol·L-1NaCl 溶液浸泡30 min 洗涤。 分别测量、计算每个纤维素降解菌周围的透明水解圈的平均直径,经比较筛选出降解能力较强的菌株。
1.2.3 纤维素降解菌的复筛 将初筛得到的菌株分别接种于50 mL 0.5%的CMC-Na 液体培养基中,28 ℃振荡培养72 h。 培养完成后进行CMC-Na 酶活力和滤纸酶活力测定,复筛出酶活力更高的菌株。
1.2.4 纤维素酶活力测定 采用二硝基水杨酸法(DNS 法),并在冯海玮等(2013)的方法上加以改进,测定还原糖含量。 首先取干燥的葡萄糖配制1 mg·mL-1葡萄糖溶液,再取7 只试管,分别按表1 加入各试剂,沸水浴5 min,立刻冷却至室温,并在540 nm 波长处测定吸光值(OD值),并以葡萄糖含量为横坐标,吸光值为纵坐标绘制葡萄糖标准曲线。
CMC-Na 酶活力测定,在段杰(2015)方法的基础上略有改进,各菌株取10 mL 菌液,4 ℃,4 000 r·min-1离心15 min,取2 mL 上清液于试管中,并加入2 mL 1% CMC-Na 溶液进行充分混合,60 ℃水浴30 min。 加3 mL DNS 试剂,沸水浴5 min,迅速冷却至室温,并定容至10 mL。 在540 nm 处测定各菌株吸光值,按葡萄糖标准曲线计算每个菌株的葡萄糖含量。 其中,对照组以2 mL 蒸馏水代替上清液。
滤纸酶活力测定,在段杰(2015)方法的基础上略有改进:各菌株取10 mL 菌液,4 ℃,4 000 r·min-1离心15 min,取2 mL 上清液加入50 mg 干燥滤纸条于试管中振荡充分混合,60 ℃水浴30 min。 取出滤纸条,加入3 mL DNS 试剂,沸水浴5 min,迅速冷却至室温,并将体积定容至10 mL。 在540 nm 处测定各菌株吸光值,按葡萄糖标准曲线计算每个菌株的葡萄糖含量。 其中,以2 mL 蒸馏水代替上清液作对照组。 上述条件下,定义1 mL 酶液在60 ℃条件下,每分钟水解纤维素产生的葡萄糖的μg 数为1 个酶活单位,以U·mL-1表示。

式中:A表示计算得出的葡萄糖含量(mg);K表示稀释倍数;V表示粗酶液体积(mL);T表示反应时间(min)。
1.3 培养条件的优化
1.3.1 不同碳源浓度对菌株产酶的影响 分别接种1 mL 菌液在CMC-Na 浓度为0.5%,0.7%,0.9%,1.1%,1.3%,1.5%,1.7%,1.9%和 2.1%的 50 mLCMC-Na 液体培养基中,在30 ℃,180 r·min-1条件下培养72 h 后,在540 nm 波长下测定菌液吸光值,计算其CMC-Na 酶活力。
1.3.2 不同培养时间对菌株产酶的影响 取1 mL 菌液加入50 mL 0.5%的CMC-Na 液体培养基中,30 ℃,180 r·min-1条件下培养,每隔 6 h 在 540 nm 波长下测定菌液吸光值。 从第 48 h 开始记录数据、绘制曲线,并计算其CMC-Na 酶活力。
1.3.3 不同培养温度对菌株产酶的影响 取1 mL 菌液加入50 mL 0.5%的CMC-Na 液体培养基中,分别在 20、25、30、35、40 和 45 ℃,180 r·min-1条件下培养 72 h,培养结束后在 540 nm 波长下测定吸光值、绘制曲线,并计算其CMC-Na 酶活力。
1.3.4 响应面试验优化培养条件 在以上3个单因素试验的基础上,采用Design-Expert 8.0.6 软件中Box-Behnken 试验设计,选择碳源浓度(A)、培养时间(B)、培养温度(C)对菌株的培养条件进行优化,以菌株经培养后用DNS 法所测得的540 nm 吸光值作为响应值。 试验因素水平见表2。
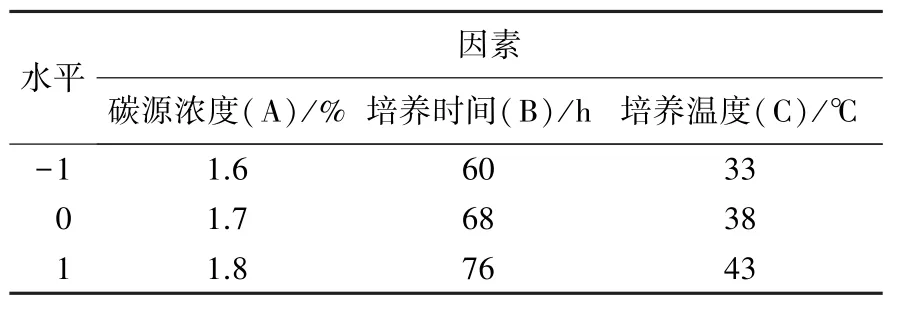
表2 响应面分析因素水平表Table 2 Independent variables and levels of the response surface analysis experiment
1.4 优化条件的检验
根据1.3.4 分析所得最佳培养条件,设计对应、可行的条件数值进行重复试验,比较菌株经培养后的540 nm 处吸光值以及回归方程模拟推测的最大吸光值,并计算其相对误差,以检验优化是否成功。
2 结果与分析
2.1 纤维素降解菌的分离和筛选
2.1.1 初筛结果 刚果红染色后即形成以菌株为中心的透明水解圈。 菌株纤维素降解能力越大,分解纤维素越多,即透明水解圈直径越大。 初筛后获得3 个优势菌株为1、5、10 号,其水解圈平均直径分别为10.50、16.50、13.33 mm。
2.1.2 复筛结果 根据葡萄糖标准曲线,计算出1、5、10 号菌株产生的葡萄糖含量。 并通过公式(1)计算CMC-Na 酶活力和滤纸酶活力(表3)。

表3 CMC-Na 酶活力及滤纸酶活力测定结果Table 3 The results of CMC-Na enzyme activity and filter paper enzyme activity test
CMC-Na 酶活力与滤纸酶活力大小均为5 号>10 号>1 号。 因此,选择 5 号菌株、10号菌株进行后续培养条件优化的研究。
2.2 纤维素降解菌培养条件的优化
2.2.1 不同碳源浓度对菌株酶活的影响 如图1 所示,5 号菌和10 号菌在低浓度CMC-Na 培养基中,酶活力都随着碳源浓度的上升而逐步上升。 当碳源浓度达到1.7%时,5 号菌和10 号菌的CMC-Na 酶活力都达到最高值,分别为212.625 和49.657 U·mL-1。 到达最高值后,两菌株的酶活力都随着碳源浓度的增加而快速下降。
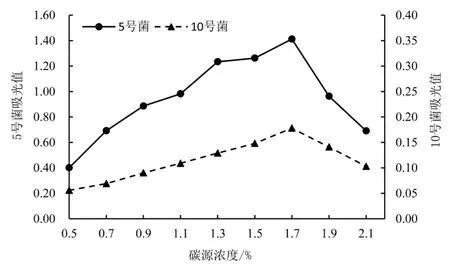
图1 不同碳源浓度对菌株酶活的影响Figure 1 Effect of different carbon source concentrations on enzyme activity
2.2.2 不同培养时间对菌株酶活的影响 如图2 所示,在培养48 h 后,5 号菌的酶活力随着培养时间的增加而平稳上升,当培养时间达到 72 h 时,CMC-Na 酶活力达到最高值172.742 U·mL-1。 在培养时间达到54 h 前,10 号菌株的产酶能力很弱,酶活力都很低,这可能是因为10 号菌的繁殖能力相对较弱;54 h 后,随着培养时间增加,菌株酶活力逐步升高,当培养时间为66 h 时,其CMC-Na 酶活力达到最高值48.996 U·mL-1。 到达最高值后,两菌株的酶活力都随着培养时间的进一步增加而呈缓慢下降趋势。
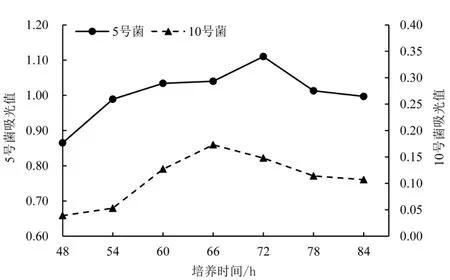
图2 不同培养时间对菌株酶活的影响Figure 2 Effect of different culture times on enzyme activity
2.2.3 不同培养温度对菌株酶活的影响如图3 所示,在低温培养条件下,5 号菌株的酶活力随着培养温度的升高而稳定增加。 当培养温度达到 40 ℃时,菌株的CMC-Na 酶 活 性 达 到 最 高 值177.100 U·mL-1。 在低温培养条件下,10号菌的活性逐渐增加,当温度达到35 ℃时, 其 CMC-Na 酶 活 性 达 到 最 高 值47.940 U·mL-1。 到达最高值后,两菌株的酶活力都随着培养温度的升高而持续下降。
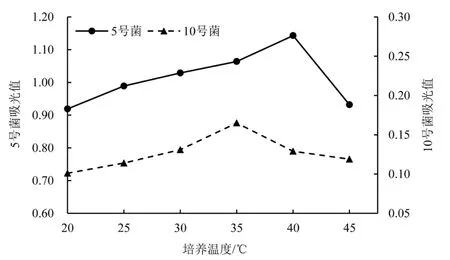
图3 不同培养温度对菌株酶活的影响Figure 3 Effect of different culture temperatures on enzyme activity
2.2.4 响应面试验优化 根据以上3 个单因素试验的试验结果,以菌株在540 nm 处的吸光值为因变量,碳源浓度、培养时间、培养温度为自变量,选用3 因素3 水平的Box-Behnken 进行试验分析(表4、表5)。
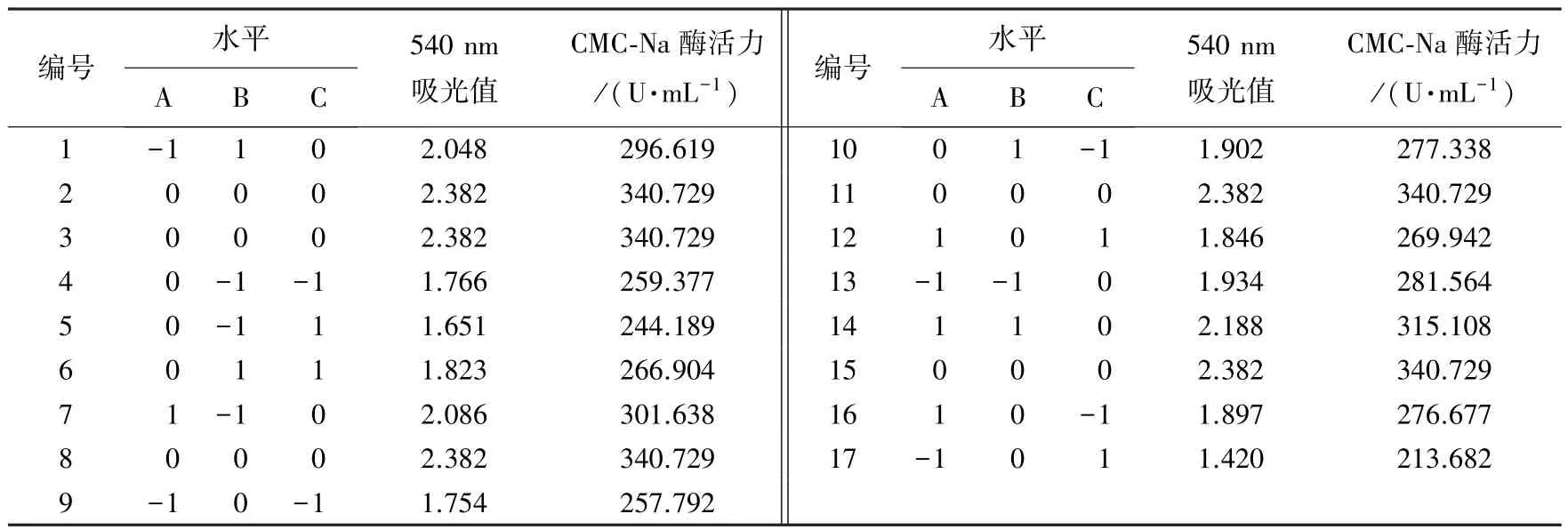
表4 5 号菌响应面试验设计及结果Table 4 The results of response surface methodology for bacteria No.5
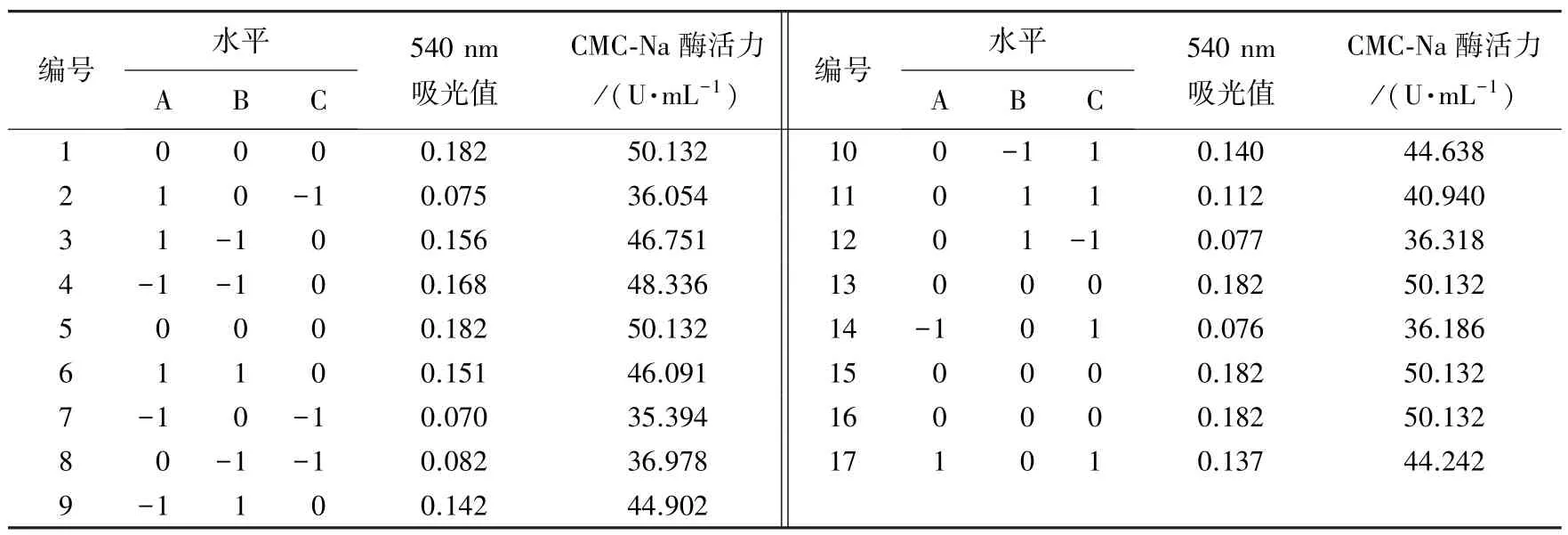
表5 10 号菌响应面试验设计及结果Table 5 Design and results of response surface methodology for bacteria No.10
分别对碳源浓度(A)、培养时间(B)、培养温度(C)3 个影响因素的试验数据进行回归拟合(表6、表7),得到两个菌株经培养后540 nm 处吸光值的二次多项式回归方程。
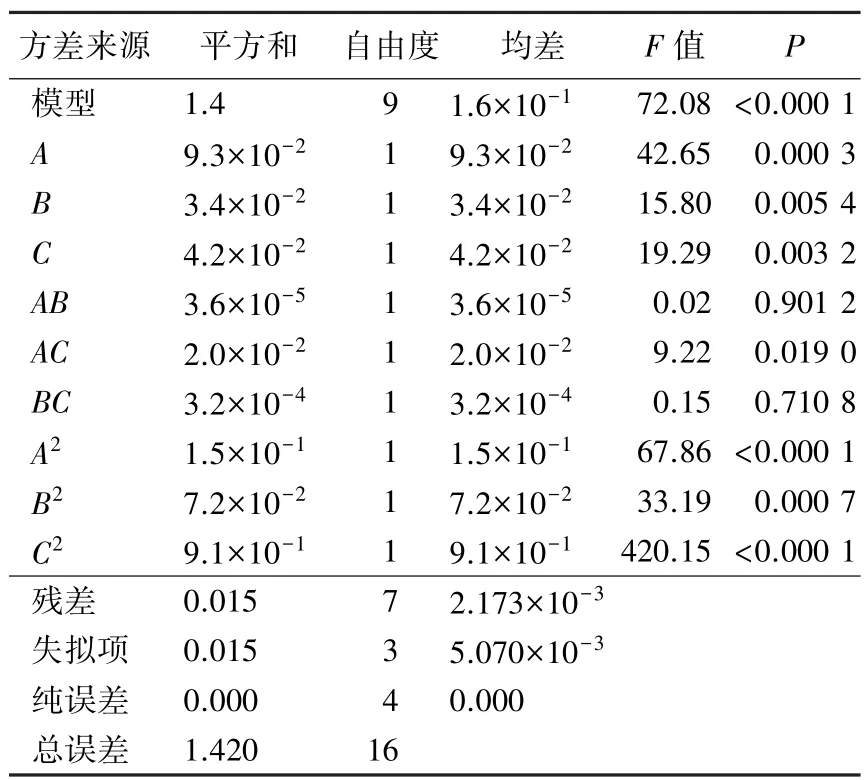
表6 5 号菌回归方程的显著性检验Table 6 Significance analysis of the regression model of bacteria No.5
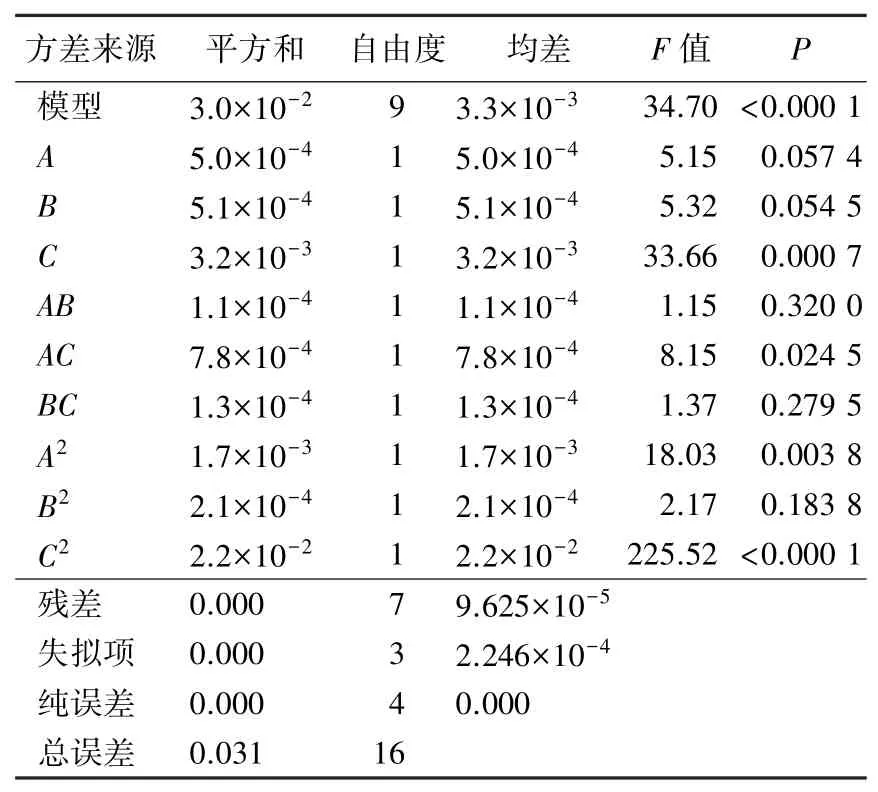
表7 10 号菌回归方程的显著性检验Table 7 Significance analysis of the regression model of bacteria No.10
5 号菌的回归方程为:吸光值=2.380+0.110A+0.066B-0.072C-3.000×10-3AB+0.071AC+9.000×10-3BC-0.190A2-0.130B2-0.470C2。 10 号菌的回归方程为:吸光值 = 0.180 +7.875×10-3A-8.000×10-3B+ 0.020C+ 5.250×10-3AB+0.014AC-5.750 × 10-3BC-0.020A2-7.050×10-3B2-0.072C2。
在菌株吸光值回归模型的显著性考察中,当P<0.05 时,说明模型差异显著,P<0.01 时,表示模型差异极显著(张学彬等,2019)。 根据表6 可知,在5 号菌的回归方程中,一次项A、B、C,二次项A2、B2、C2均达到极显著水平;根据表7 可知,在10 号菌回归方程中,一次项C,二次项A2、C2达到极显著水平。 由此可得,两个回归模型拟合度都较好,预测值较为准确,可采用回归方程计算值模拟试验结果,进行预测和研究分析。
根据P值的比较结果可以得出,3 个因素对5 号菌酶活力影响程度的顺序为:碳源浓度>培养温度>培养时间;10 号菌为:培养温度>培养时间>碳源浓度。 其中两种菌株的交互作用项中均为AC的P值最小,表示碳源浓度和培养温度的交互作用对两种菌株酶活力的影响最显著。
由响应面试验法分析得出,5 号菌最佳培养条件为碳源浓度1.73%,培养时间69.96 h,培养温度37.78 ℃,预测吸光值为2.407;10 号菌最佳培养条件为碳源浓度1.72%,培养时间63.41 h,培养温度38.91 ℃,预测吸光值为0.186。
2.2.5 响应面分析 根据所拟合的响应面的形状,分析3 种因素对菌株酶活力的影响。 等高线的形状呈椭圆形表示两因素作用显著,呈圆形则与之相反。 3D 交互图曲面图则根据形成的平面坡面陡峭度来判断显著性,坡面越陡峭,因素的交互作用显著,否则与之相反(李佳桥等,2016)。
由图4~图6 可知,5 号菌AB 交互作用的等高线近似圆形,交互曲线较为平滑,两者的交互作用不显著,而AC、BC 交互作用的等高线虽然都是椭圆,但AC 的交互曲线较为陡峭,因此,AC 两元素间的交互作用最为显著。 由图7 ~ 图 9 可知,10 号菌 AB、AC、BC 交互作用的等高线都为椭圆,但AC 的3D 交互图曲面先上升后下降且最为陡峭,因此,AC 两元素间的交互作用最为显著。 综上,碳源浓度(A)和培养温度(C)的交互作用对5 号菌和10 号菌的酶活力影响最显著,这与2.2.4 分析结果一致。
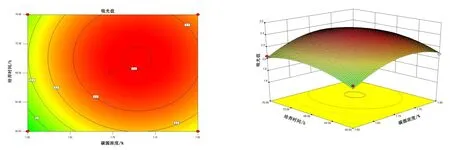
图4 5 号菌碳源浓度、培养时间交互作用对菌株酶活的影响Figure 4 The interaction effects of carbon source concentration and culture time on the enzyme activity of bacteria No.5
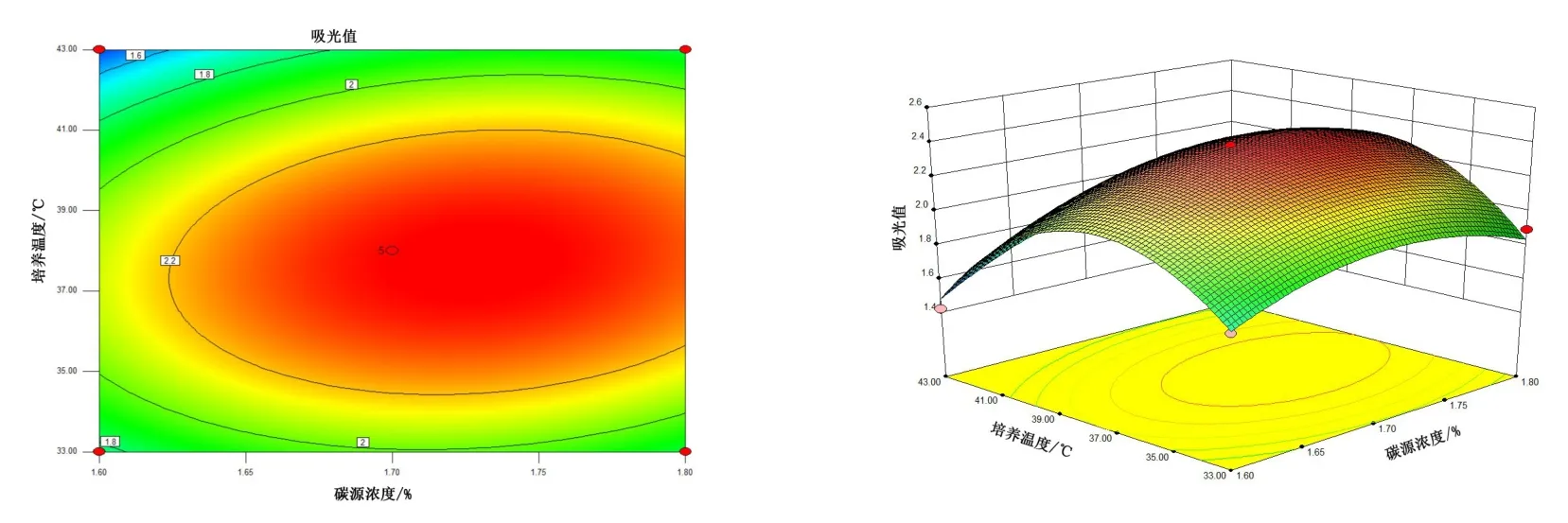
图5 5 号菌碳源浓度、培养温度交互作用对菌株酶活的影响Figure 5 The interaction effects of carbon source concentration and culture temperature on the enzyme activity of bacteria No.5
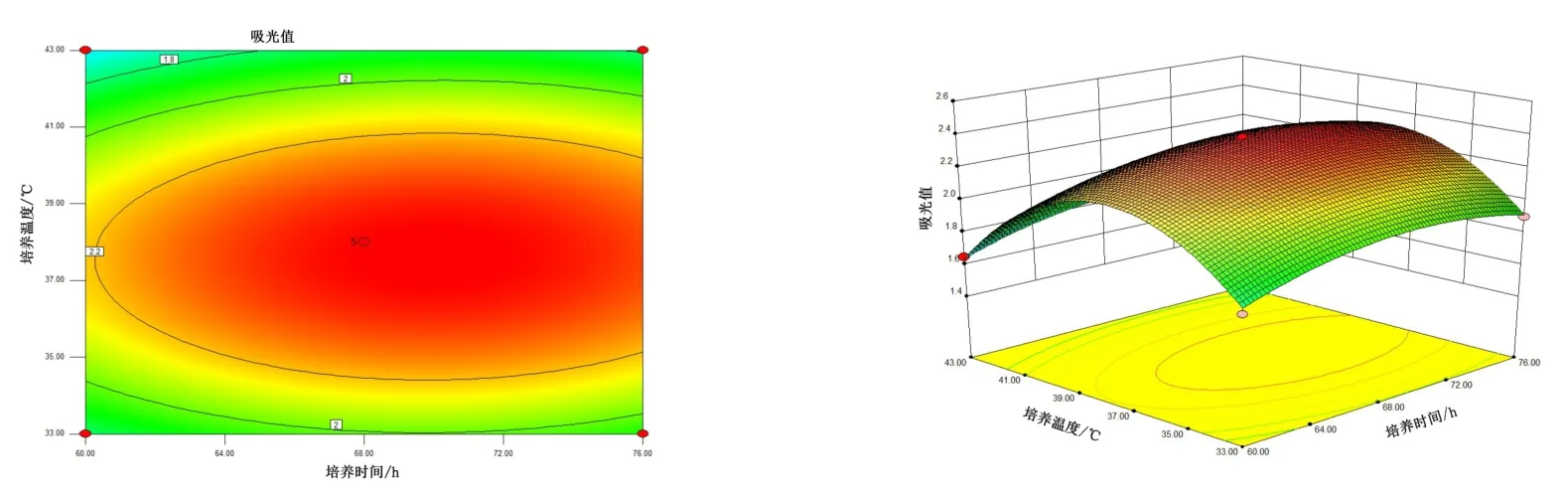
图6 5 号菌培养时间、培养温度交互作用对菌株酶活的影响Figure 6 The interaction effects of culture time and culture temperature on the enzyme activity of bacteria No.5
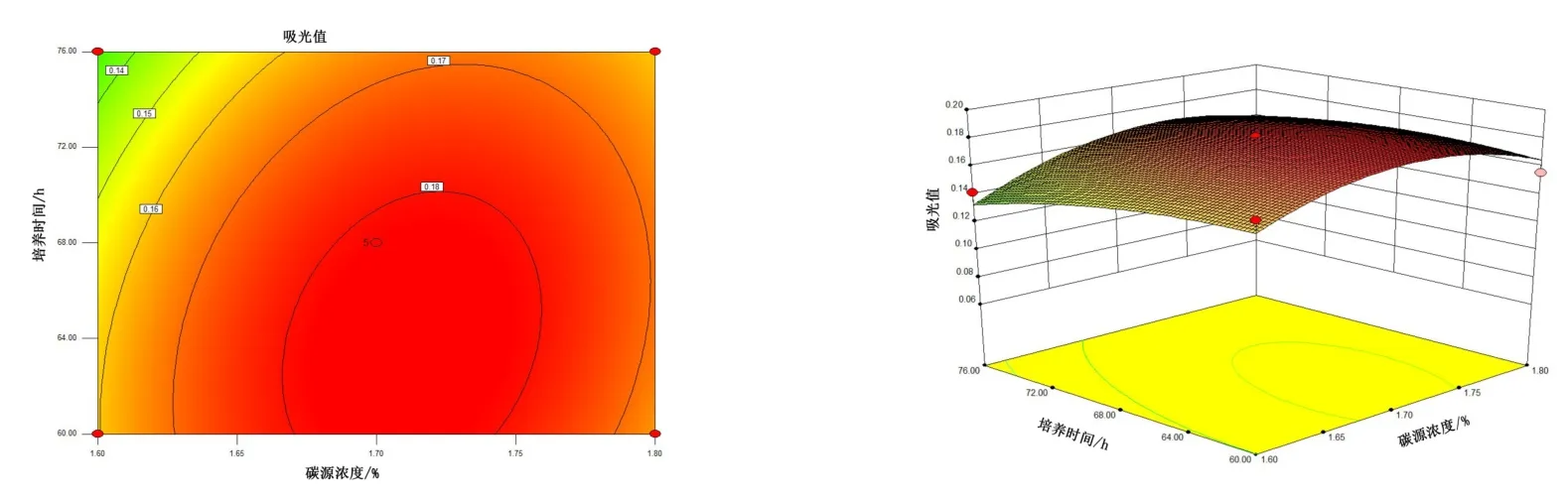
图7 10 号菌碳源浓度、培养时间交互作用对菌株酶活的影响Figure 7 The interaction effects of carbon source concentration and culture time on the enzyme activity of bacteria No.10
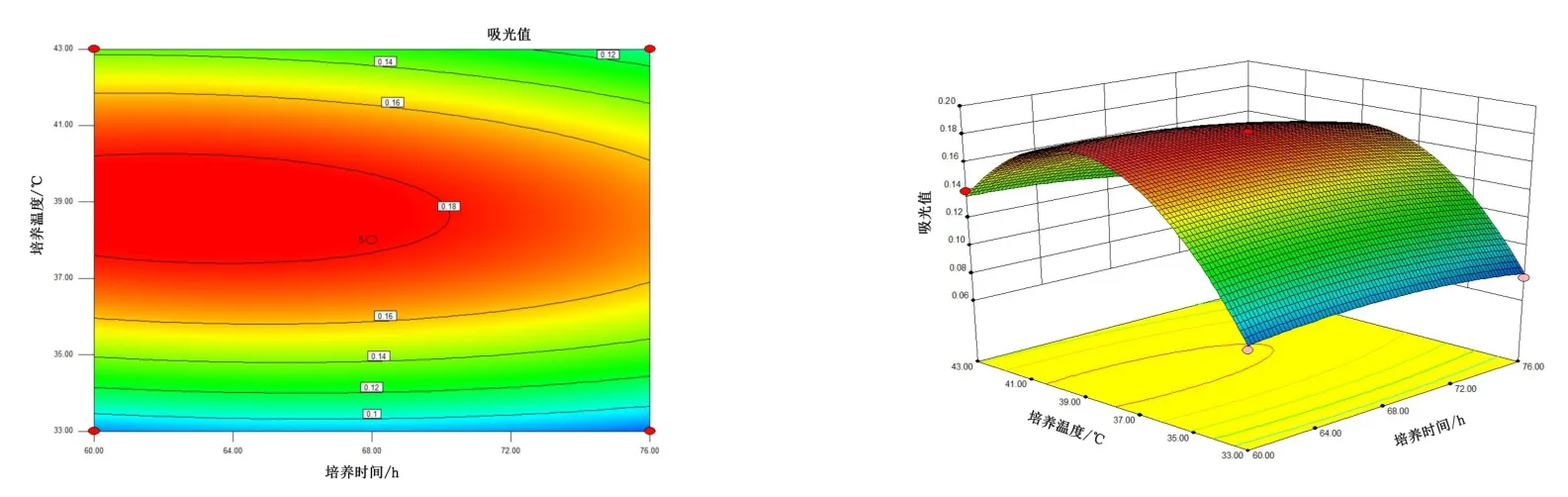
图9 10 号菌培养时间、培养温度交互作用对菌株酶活的影响Figure 9 The interaction effects of culture time and culture temperature on the enzyme activity of bacteria No.10
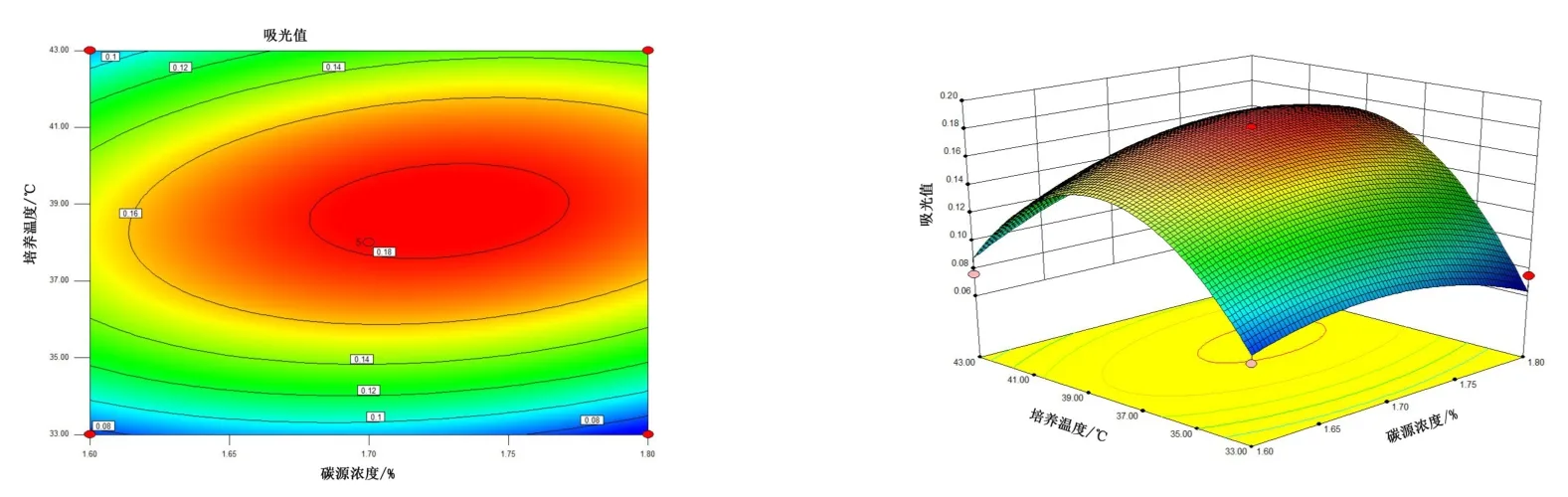
图8 10 号菌碳源浓度、培养温度交互作用对菌株酶活的影响Figure 8 The interaction effects of carbon source concentration and culture temperature on the enzyme activity of bacteria No.10
2.3 优化条件检验
为检验响应面分析结果的准确性,根据试验的操作性和可行性进行进一步的结果检验,设计5 号菌在CMC-Na 浓度为 1.73%,培养时间 70.0 h,培养温度37.8 ℃的条件下进行培养;10号菌在CMC-Na浓度为1.72%,培养时间为63.5 h,培养温度39.0 ℃的条件下进行培养。 运用回归方程模拟预测5 号菌在540 nm 处吸光值为2.407,10 号菌为0.186。 而在优化条件下培养后,5 号菌实际测得的吸光值为2.389,10 号菌测得的吸光值为0.182(表8)。 由此可知,试验观测值与模型预测值较为接近,两菌株的检验结果与预测值相对误差均小于2.5%。 这表明模型预测性良好,所得经优化的菌株最适培养条件稳定合理,客观可行。
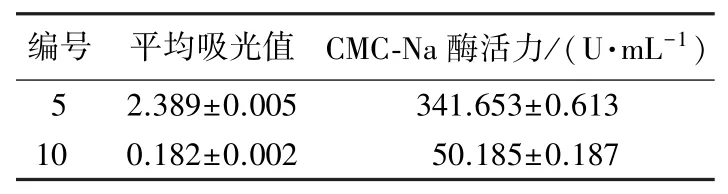
表8 优化条件检验Table 8 The results of the optimum condition test
3 结论与讨论
本研究以发酵半年的厨余垃圾为原材料,运用羧甲基纤维素钠培养基分离纯化,并使用刚果红染色法、CMC-Na 酶活力以及滤纸酶活力检测进行初筛和复筛,最终成功筛选获得5 号菌和10 号菌两株优势纤维素降解菌。
单因素试验表明,两菌株的CMC-Na 酶活力都随着碳源浓度的上升而上升,并在碳源浓度为1.7%时达到最高值,此后开始快速下降。 这可能是因为在碳源浓度过低时,菌株的营养不足,导致菌体生长缓慢或不良;在碳源浓度过高时,虽然菌株的能量、碳源充足,但会使菌体生长过于旺盛,导致发酵液粘度过大,氧气供应不足,使得产酶量降低(张粲,2014)。 两菌株的CMC-Na 酶活力也随着培养时间的增加而上升,5 号菌培养72 h 时,酶活力达到最高值;10 号菌培养66 h 时,酶活力达到最高值。 此后,两菌株的酶活力都随着培养时间的进一步增加而缓慢下降。 这可能是因为菌株的产酶活力会随着其进入对数生长期而逐渐升高,在进入稳定期后,其产酶活力达到峰值。 此后,培养基中的营养物质不断被消耗,菌株代谢产物也不断积累,从而导致菌株生长缓慢甚至死亡,其产酶活力大小也呈现出下降趋势(张悦等,2018)。 两菌株的CMC-Na 酶活力同样随着培养温度的升高而升高,5 号菌的培养温度达到40 ℃时,酶活力达到最高值;10 号菌的培养温度达到35 ℃时,酶活力达到最高值。 当温度继续升高,两菌株的酶活力都开始下降。 这可能是因为过低或过高的培养温度会降低菌株的活力,影响其繁殖效率,从而导致产酶能力下降,酶活力降低(冯海玮等,2013)。
为了进一步优化两菌株的培养条件,根据单因素试验的结果,又进行了响应面试验优化,所得模型预测性良好,所得到的菌株最适培养条件稳定合理,客观可行。 结果表明,两种菌株的条件优化结果与单因素试验结果相比较,最适培养时间都缩短了,即菌株的培养条件经优化后不仅能得到更高的酶活力,同时还提高了产酶效率。 崔秀秀等(2016)利用此方法优化耐冷纤维素降解菌,使其CMCase 酶活较优化前提高2.96 倍、杨培新等(2015)利用此方法优化分离得到的1 株产纤维素酶真菌,使其发酵产纤维素酶活性提高了15.02%,也都证明了使用响应面法优化菌种培养条件是可行、有效的。
本研究从厨余垃圾中筛选出两株优势纤维素降解菌,并通过响应面法优化了两菌株的培养条件,使菌株有了更高的产酶效率及酶活力,在降解纤维素应用方面有很好的潜力。
