茂兰喀斯特森林不同地形土壤酶与植物多样性的关系1)
2021-12-03廖全兰龙翠玲薛飞熊玲
廖全兰 龙翠玲 薛飞 熊玲
(贵州师范大学,贵阳,550001)
在森林土壤中,土壤酶是其重要的组成部分,其参与土壤有机物的转化、分解、合成等重要生态学过程,能调控森林生态系统的养分及能量循环[1-2]。植物群落可通过影响土壤酶活性来控制土壤养分的代谢,进而导致土壤养分含量的变化[3]。在森林生态系统中,人们对森林土壤酶的研究主要集中在以下几个方面:不同植被类型土壤酶活性研究[4-6]、土壤酶活性在不同植被演替阶段的研究[7-9]、土壤酶活性与植物多样性的研究[10-11]等。而地形能够引起环境中水热条件的重新分配,影响土壤养分、植物群落结构及土壤微生物组成[12]。因此,不同地形植被类型的差异对土壤酶活性也会产生一定的影响,探讨土壤酶活性与植物的相互作用及机制具有重要意义。
茂兰喀斯特森林是世界同纬度地区仅有的原生性较强的喀斯特森林[13],与常态地貌森林相比,茂兰喀斯特森林属于特殊的亚热带森林生态系统,其在生态环境,植物群落水平、垂直结构、演替更新动态,森林群落性质、外貌、区系组成,生态系统对周围环境的影响等方面均有显著不同[14]。茂兰喀斯特森林地貌复杂多样,其中峰丛漏斗、峰丛洼地、槽谷等是林内分布极广的主要地貌类型[15]。在茂兰喀斯特森林土壤-植被连续系统中,有研究对土壤微生物及土壤酶活性[16]、群落结构及物种多样性[17]、植物多样性与土壤养分等[18]的关系进行了研究,而对于植物多样性是否会引起土壤酶活性变化的研究较少。因此在前人研究工作的基础上,进一步探讨茂兰喀斯特森林土壤酶活性与植物多样性的关系,可以为茂兰国家级自然保护区的长期管理、土壤改良、物种多样性保护等方面提供参考。
1 研究区概况
贵州茂兰国家级喀斯特森林自然保护区(107°52′10″~108°5′40″E,25°9′20″~25°20′50″N)位于中国西南地区贵州省南部黔、桂交界处。此地属亚热带季风湿润气候,夏季高温多雨,冬季温和湿润,四季分明。林内有丰富的生物资源,植被类型主要为常绿落叶阔叶混交林。区域内地势起伏不大,相对高差约648.6 m。年平均温15.3 ℃,气温年较差为18.3 ℃。雨季集中分布于4—10月,年降水量约1 752.5 mm,年平均相对湿度为83%。特殊的喀斯特地貌,导致区内水文地质条件复杂,林内岩石露头多,岩石裂隙发育,岩石透水性极强,地表水强烈渗漏。区域内土壤分布不均,如漏斗地形有成片土壤分布,水分条件好(约52%),但部分地段有积水;槽谷地形谷底平坦,水分条件适中(约44%);坡地地形土层浅薄不连续,土壤多分布于石缝隙且水分条件差(约34%)。研究区域3种地形部位的基本概况如表1所示[19]。
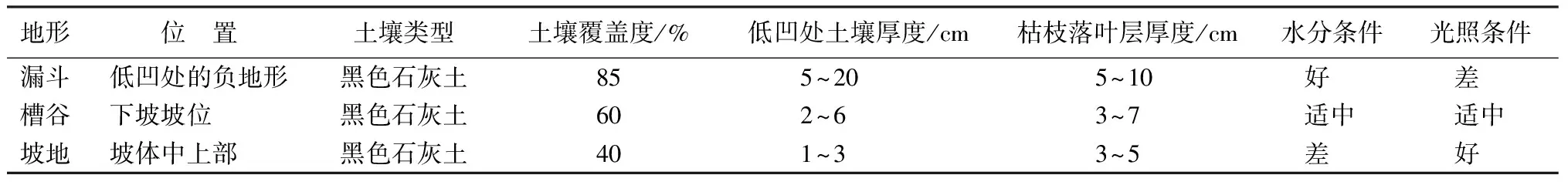
表1 3种地形的基本特征
2 材料与方法
2.1 样地选择与植被调查
2019年6—7月在茂兰自然保护区选取漏斗、槽谷、坡地3种地形,分别在每种地形中选取典型地段设置面积为20 m×20 m的连续样地10个,共30个样地。采用GPS仪器进行定位,并记录每个样地的水分条件、光照条件、土壤类型、海拔、坡位、坡向等环境因子。对样地内的乔木、灌木进行每木调查,记录乔、灌木的种名、株数、树高、胸径、枝下高、冠幅等。
2.2 土壤样品采集与处理
在每种地形随机选择3个20 m×20 m的大样方,以梅花五点法设置近2 m2采样区,每个采样区中设置3个采样点。采样时先将采样点表层凋落物等杂物去除,因林下土壤层较浅薄,故采集0~10 cm土壤层,将同一采样区内的3个采样点土壤等比例混合均匀。土样带回实验室后,挑出可见的凋落物、石头、残根等,自然风干,过1 mm筛,用于土壤酶活性测定。
2.3 土壤酶活性的测定
采用高锰酸钾滴定法测定过氧化氢酶活性;采用磷酸苯二钠比色法测定碱性磷酸酶活性;采用苯酚钠-次氯酸钠比色法测定脲酶活性;采用3,5-二硝基水杨酸比色法测定蔗糖酶活性[20]。
2.4 植物群落特征计算
采用生物群落多样性的测度方法计算乔、灌木层的重要值。计算公式如下[21]:
重要值(IV)=相对密度+相对频度+相对显著度
植物多样性指数选取Margalef丰富度指数(Rl)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)、Simpson优势度指数(λ)来表示。计算公式如下[17]:
Margalef丰富度指数:Rl=(S-1)/log2N;
Shannon-Wiener多样性指数:H′=-∑(Pilog2Pi);
Pielou均匀度指数:J=H′/log2S;
Simpson优势度指数:λ=∑(ni(ni-1)/N(N-1))。
式中:S为物种数目,N为群落中所有物种个体数总和,Pi为第i个物种的个体数(ni)与群落总个体数(N)的比值,即Pi=ni/N。
2.5 数据处理
采用Excel软件进行数据的整理及预处理,使用SPSS 21.0对不同地形土壤酶活性及植物多样性进行方差分析及多重比较(LSD法),使用Excel软件进行绘图,使用Canoco 4.5对土壤酶活性、植物群落特征进行冗余分析(RDA)、建立二维排序图。
3 结果与分析
3.1 不同地形土壤酶活性变化
4种土壤酶在不同地形内所表现的酶活性由表2所示。3种地形的过氧化氢酶活性由大到小依次为槽谷、漏斗、坡地,且坡地地形的过氧化氢酶活性与漏斗、槽谷地形的有显著差异。3种地形的碱性磷酸酶活性由大到小依次为槽谷、坡地、漏斗,且槽谷地形的碱性磷酸酶活性与漏斗地形的差异性显著,坡地地形的碱性磷酸酶活性与漏斗、槽谷地形的均无差异性。3种地形的脲酶活性由大到小依次为漏斗、坡地、槽谷,且漏斗地形的脲酶活性与槽谷、坡地地形的有显著性差异。3种地形的蔗糖酶活性由大到小依次为槽谷、漏斗、坡地,其中槽谷与坡地地形的蔗糖酶活性间差异性显著,漏斗地形的蔗糖酶活性与槽谷、坡地地形的无显著性差异。综上所述,4种土壤酶在不同地形中,其酶活性有显著差异性。

表2 不同地形土壤酶活性
3.2 不同地形木本植物组成
对茂兰喀斯特森林3种地形的木本植物进行群落调查及统计,其中漏斗地形中木本植物共计35科61属84种,槽谷的木本植物共计38科64属89种,坡地的木本植物共计35科65属78种。计算乔、灌层植物的重要值,划分3种地形植物群落乔、灌层的优势物种,重要值排前10的优势种统计结果见表3。
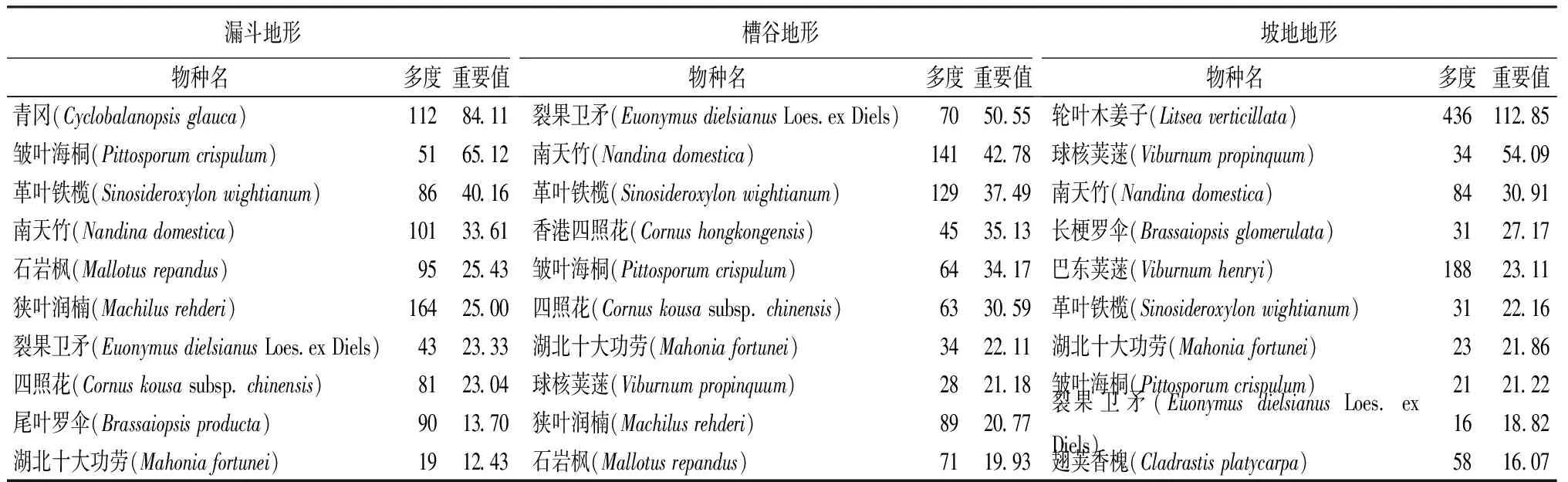
表3 不同地形中重要值前10的优势种
由表3可知,对各地形的优势种按照重要值大小进行统计,结果显示,虽然不同地形植被类型均为常绿落叶阔叶混交林,但各地形的物种多样性差异较大。3类样地中重要值排名前10的优势种各不相同,侧面证明了茂兰喀斯特森林区域物种多样性较为丰富。漏斗地形的显著优势种为青冈(Cyclobalanopsisglauca),重要值达到84.11,多度为112;槽谷地形的最大优势种为裂果卫矛(EuonymusdielsianusLoes. ex Diels),重要值达到50.55,多度为70。坡地地形的最大优势种为轮叶木姜子(Litseaverticillata),以多度维持优势,其多度为436,重要值为112.85。重要值前10的物种多度占样地总个体数的比例由大到小依次为坡地(52.46%)、漏斗(51.47%)、槽谷(43.72%)。
3.3 不同地形木本植物多样性
各地形物种多样性通过Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(Rl)、Pielou均匀度指数(J)及Simpson优势度指数(λ)进行表征(表4)。不同地形的物种数不同,其中槽谷物种数最多,坡地物种数最少。Shannon-Wiener多样性指数是物种数和相对密度综合形成的指数。漏斗、槽谷地形的Shannon-Wiener多样性指数大于坡地地形的,且漏斗地形的多样性指数与槽谷地形的无显著差异。不同地形间的Margalef丰富度指数无显著差异。Pielou均匀度指数表示物种个体数之间的差异。漏斗、槽谷地形的Pielou均匀度指数高于坡地地形的;且漏斗地形的均匀度指数与槽谷地形间无显著差异。Simpson优势度指数由于受优势种影响较大,表现为坡地地形的Simpson优势度指数高于漏斗、槽谷地形的,且漏斗地形的优势度指数与槽谷地形间无显著差异。由此可知,漏斗、槽谷地形的植物多样性指数高于坡地地形的,但坡地地形的优势度指数高于槽谷、漏斗地形的。进一步说明,漏斗及槽谷地形的木本植物数量分布较均匀,而坡地地形物种数量分布不均匀,优势种的地位突出。
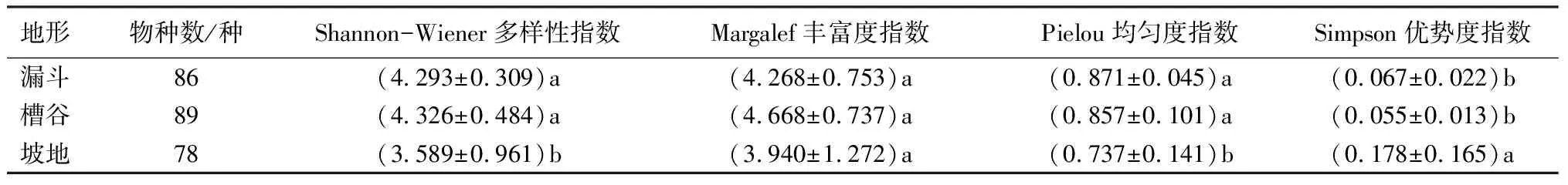
表4 不同地形植物多样性指数
3.4 土壤酶活性与植物多样性的关系
采用冗余分析(RDA)对土壤酶活性与植物多样性的关系进行探究,结果如表5、表6及图1所示。表5为植物多样性指数对4种土壤酶活性差异性的解释量,前2轴4种土壤酶活性解释量分别为47.9%、4.3%,即第Ⅰ轴、第Ⅱ轴植物多样性可累计解释土壤酶活性特征的53.2%,且对土壤酶活性与植物多样性关系的累计解释量达99.1%。这表明前2轴能反映土壤酶活性与木本植物多样性关系的绝大部分信息,且主要由第Ⅰ轴决定。

表5 土壤酶活性RDA排序的特征值及累积解释量
图1为土壤酶活性与植物多样性指数的二维排序图,图中空心箭头为土壤酶活性,实心箭头为植物多样性指数。从二维排序图可知,物种数、Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数、Simpson优势度指数箭头连线均较长,均能较好地解释土壤酶活性的差异。但碱性磷酸酶、脲酶活性箭头连线较短,说明这2个酶活性与植物多样性之间的关系较弱。Shannon-Wiener多样性指数、Pielou均匀度指数、Margalef丰富度指数、物种数与蔗糖酶活性夹角较小,呈显著正相关;Simpson优势度指数与蔗糖酶活性夹角较小,呈显著负相关。Pielou均匀度指数、Shannon-Wiener多样性指数、Margalef丰富度指数与过氧化氢酶活性的夹角较小,呈正相关;物种数、Simpson优势度指数与过氧化氢酶活性的夹角较小,呈负相关。
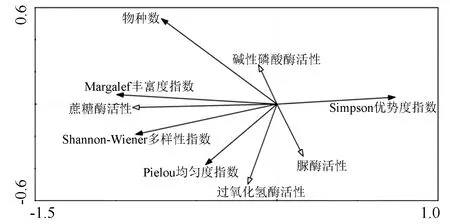
图1 土壤酶活性与植物多样性指数的冗余度
由二维排序图可知,不同植物多样性指数对土壤酶活性的影响差异较大。使用蒙特卡洛检验对影响土壤酶活性的植物多样性进行重要性排序,结果如表6所示。
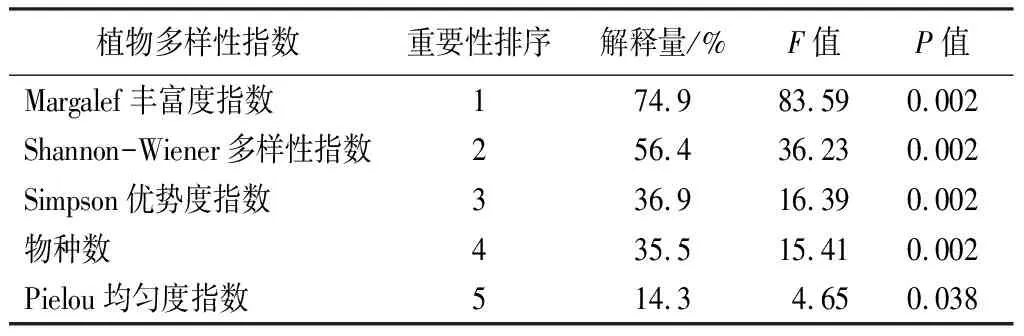
表6 植物多样性指数解释的重要性排序和显著性检验结果
其重要性由大到小依次为Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数、物种数、Pielou均匀度指数。除Pielou均匀度指数对土壤酶活性影响较小且未达显著水平外,其余植物多样性指数均与土壤酶活性呈极显著水平(P<0.01)。
4 讨论
4.1 地形对土壤酶活性的影响
本研究中,不同地形的土壤酶活性存在差异,其中槽谷的过氧化氢酶、碱性磷酸酶、蔗糖酶活性均大于漏斗、坡地的,且槽谷地形的这3类酶活性与漏斗、坡地地形间存在显著性差异。不同地形的脲酶活性由大到小依次为漏斗、坡地、槽谷,这与廖全兰等[22]的研究结果相似。推断是因为槽谷地形的光照强度、温度、含水量等均较适中,微生物活性较强,且物种多样性最丰富,使槽谷地形的土壤过氧化氢酶、碱性磷酸酶、蔗糖酶的活性均最大。脲酶活性反映土壤供氮水平。已有研究表明,漏斗地形全氮量最大,使得脲酶活性在漏斗地形中表现出最大值[22]。本研究表明,土壤过氧化氢酶、碱性磷酸酶、蔗糖酶活性在3种不同地形间的变化与木本植物多样性的变化趋势相似,匀表现为槽谷的土壤过氧化氢酶、碱性磷酸酶、蔗糖酶活性大于漏斗、坡地地形的。植物可直接或间接地影响土壤酶活性[4],地形变化会引起森林环境水热条件的重新分布[12],进而改变植物群落构成,使不同地形间土壤养分含量出现差异,导致土壤酶在不同地形间表现出不同的活性差异。综上所述,地形变化引起植物群落结构的变化,进一步引起林下环境的变化,导致不同地形土壤酶活性存在差异。
4.2 地形对植物多样性的影响
地形影响太阳辐射及降水的空间再分配,导致土壤温度、水分、养分等资源的空间异质性,进而影响植被分布[23-24]。在山地森林群落中,地形是影响木本植物分布、森林群落空间格局等的重要因素[25]。植物多样性指数能客观地反应植物群落中物种的丰富度、均匀度及群落的优势度。本研究中,植物多样性指数随着地形的变化,其大小也随之改变。其中槽谷的物种数、Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数均大于漏斗及坡地的,而Simpson优势度指数则在坡地出现最大值,且与漏斗、槽谷地形的优势度指数有显著差异。出现这种现象的原因是3种地形中槽谷地形地势平缓,光照条件、水分及土壤养分均较好,植物适宜在此环境下生长,故而槽谷地形植物丰富[17];坡地地形地势陡峭,土层浅薄,保水性能差,水土流失严重,生境条件比较干旱,优势种主要以灌木及耐旱性乔木为主,林下光照条件较好,有利于优势种的生长、分布[25]。综上所述,不同地形植物多样性的差异,主要是地形通过水热条件影响植被的类型及分布。不同地形重要值排名前10的优势种各不相同,旁证了茂兰喀斯特森林区域物种多样性较高。
4.3 土壤酶活性与植物多样性的关系
土壤酶活性的变化规律不仅与植被的演替有关,还与植被树种组成有关[26]。不同的植物种类及组成,其凋落物的种类、养分含量、根系分泌物不同,导致土壤酶活性的差异[7]。本研究中,Shannon-Wiener多样性指数、Pielou均匀度指数、Margalef丰富度指数与蔗糖酶、过氧化氢酶活性呈正相关,Simpson优势度指数与蔗糖酶、过氧化氢酶活性呈负相关,这与杨万勤等[7]研究结果一致。植物种类、数量的提高促进土壤酶活性,这是因为植物种类、数量的提高可为土壤表层积累较多的枯枝落叶。腐殖质及土壤中养分含量高、土壤疏松、通气与水热条件良好的生境有利于土壤微生物的繁殖及生长,使得土壤酶活性增强[27-28]。本研究还发现,碱性磷酸酶、脲酶活性与植物多样性之间关系较弱,是因为茂兰喀斯特森林环境资源稀缺,生境条件严酷,土壤承载能力较差,而植物对土壤养分的需求过大,从而造成茂兰不同地形森林土壤酶活性与植物多样性的变化存在差异,且植物的生长不会立即对土壤生物产生影响。植物多样性的差异与土壤生物学的变化不同步,植物凋落物进入土壤后才会引起土壤微生物及养分的变化[29],因此部分土壤酶活性与植物多样性相关性较弱。综上所述,植物可通过凋落物及植物根系影响土壤酶活性,使土壤酶活性与植物多样性存在相关性,但其相关性因酶的种类不同而呈现差异性。
5 结论
茂兰喀斯特森林土壤酶活性在不同地形间存在显著性差异,其中槽谷地形的过氧化氢酶、碱性磷酸酶、蔗糖酶活性均大于漏斗及坡地地形的,过氧化氢酶、蔗糖酶活性在槽谷与坡地地形间存在显著性差异;脲酶活性在漏斗地形中出现最大值,且漏斗地形的脲酶活性与槽谷、坡地地形的存在显著性差异。不同地形间植物多样性存在显著差异,其中槽谷地形的物种数、Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数均大于漏斗、坡地的;而Simpson优势度指数的最大值出现在坡地地形,且坡地地形的优势度指数与漏斗、槽谷地形的有显著差异。不同地形间重要值排名前10的优势种各不相同,旁证了茂兰喀斯特森林区域物种多样性较高。冗余分析结果表明,Shannon-Wiener多样性指数、Pielou均匀度指数、Margalef丰富度指数与蔗糖酶、过氧化氢酶活性呈正相关,Simpson优势度指数与蔗糖酶、过氧化氢酶活性呈负相关,碱性磷酸酶、脲酶活性与植物多样之间关系较弱。植物可通过凋落物及植物根系影响土壤酶活性,土壤酶活性与植物多样性存在相关性,但其相关性因酶的种类不同而呈现差异性。
