功能低聚糖对植物乳杆菌ZDY2013发酵乳发酵特性及冷藏效果的影响
2021-12-01张娜占英孟迎平李一娟陶雪莹魏华张志鸿
张娜,占英,孟迎平,李一娟,陶雪莹,魏华,2,张志鸿,2*
(1.南昌大学食品科学与技术国家重点实验室,江西南昌 330047)
(2.南昌大学中德联合研究院,江西南昌 330047)
益生菌是指摄入一定量后对人体产生有益健康作用的一类活的微生物。目前,研究和应用广泛的益生菌主要包括乳酸杆菌和双歧杆菌,其中益生菌开发的乳制品广受欢迎。乳品是益生菌递送的最常见食品基质,能有效保护菌株免受食品加工、储存和胃肠道环境损伤,并促进其抵达肠道发挥益生功效[1]。研究表明,每天摄入一定量含109~1011cfu活菌的发酵乳能有效预防肠道炎症、II型糖尿病和心血管疾病等[2-4]。
植物乳杆菌ZDY2013是一株分离于自然发酵酸豆角的具有自主知识产权的优势乳酸杆菌,其能够耐受胃肠道的酸和胆盐环境,顺利通过胃肠道消化系统,并能调节肠道菌群来改善宿主健康[5]。另外,前期研究发现,利用植物乳杆菌ZDY2013制作的发酵奶能预防食源性致病菌污染,并能防止产肠毒素蜡样芽孢杆菌对小鼠造成肠道菌群稳态失衡[6,7]。
发酵乳制品中添加益生元(如牛蒡多糖、菊粉和功能低聚糖等)被认为是提高产品质量、货架期和功能的有效策略,主要表现为提高乳酸菌在产品贮藏过程中的存活率、改善发酵乳风味[8]和增强其功能特性(抗肥胖、抗菌、抗糖尿病、缓解氧化应激等)[9-12]。例如,虞娇娇等[13]发现低聚果糖和低聚半乳糖可以显著增加发酵乳中乳酸菌活菌数,同时改善发酵乳的粘稠度;Madhu等[11]研究发现,发酵乳中添加低聚果糖可以促进植物乳杆菌CFR 2194和发酵乳杆菌CFR 2192的生长代谢,提高发酵乳的抗氧化活性。抗氧化活性一直被认为是乳制品的一种基本营养特性[1],其可延长发酵乳的保质期及保护机体免受氧化损伤[14]。
低聚木糖具有良好的抗氧化活性[15],且可改善Ⅱ型糖尿病[16]、抗肥胖和增强机体免疫力等益生功能;低聚异麦芽糖可显著促进体内双歧杆菌活性,提高肠道免疫和减少腹泻[17]等益生性。然而,添加低聚木糖和低聚异麦芽糖对发酵乳功能特性的影响报道甚少。为此,本文在发酵乳中添加低聚木糖、低聚异麦芽糖或者两种低聚糖的组合物来探究其对植物乳杆菌ZDY2013发酵特性及对发酵乳的影响,同时对冷藏期发酵乳抗氧化活性进行解析。本研究将为功能低聚糖乳制品的研发与工艺优化提供参考,并为植物乳杆菌ZDY2013功能应用提升研究奠定基础。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株
植物乳杆菌(Lactobacillus plantarum)ZDY2013为本团队分离于自然发酵酸豆角,保存于食品科学与技术国家重点实验室。
1.1.2 试剂与培养基
邻苯二甲醛、三氯乙酸、β-巯基乙醇、邻菲啰啉均为分析纯,上海阿拉丁生化科技股份有限公司;1,1-二苯基-2-三硝基苯肼(DPPH)、2,2’-联氮基-双-(3-乙基苯并噻唑啉-6-磺酸)二铵盐(ABTS)、低聚异麦芽糖(纯度≥90%),低聚木糖,上海源叶生物科技有限公司;过氧化氢、硫酸亚铁、过硫酸钾、四硼酸钠,十二烷基硫酸钠,甲醇,乙醇、四硼酸钠、十二烷基硫酸钠等均为国产分析纯;脱脂乳,金薄金生态科技有限公司;MRS肉汤培养基,北京索莱宝科技有限公司。
Basal MRS(BMRS)培养基配方:10.0 g蛋白胨、5.0 g牛肉膏、5.0 g酵母膏、3.0 g氯化铵、4.0 g磷酸氢二钾、2.6 g磷酸二氢钾、0.102 g七水硫酸镁、0.05 g四水硫酸锰、0.5 g半胱氨酸和1.0 g吐温-80,加去离子水至1000 mL,加热溶解,调pH至6.2,121 ℃灭菌15 min,常温保存备用。
1.2 仪器与设备
厌氧培养箱,美国Gene Science公司;台式高速冷冻离心机,德国Eppendorf公司;PHS-3E pH计,上海伟业仪器厂;立式蒸汽灭菌锅,上海博讯仪器有限公司;电热恒温水浴锅,上海精宏实验设备有限公司;电子天平,德国Sartorius公司。
1.3 方法
1.3.1 发酵期实验方法
1.3.1.1 样品配制
配置含12 g/100 mL脱脂乳液体、BMRS培养基,10%、15%、20%、25%、30%(m/V)葡萄糖、低聚木糖、低聚异麦芽糖和低聚木糖与低聚异麦芽糖组合物(1:1)溶液,灭菌(115 ℃、20 min)后分装待用。
1.3.1.2 菌株活化
从-80 ℃冰箱中取出冻存菌株,用接种环进行划线至MRS平板中,37 ℃静置培养36 h,取活化后的单菌落接种于5 mL MRS肉汤培养基,37 ℃静置培养12 h。传代培养两次,接种量为1%。
1.3.1.3 培养基发酵液中菌株浓度和pH值测定
将上述活化后的植物乳杆菌ZDY2013接种至4 mL BMRS培养基中,加入1 mL 10%(m/V)不同低聚糖使得其终浓度为2.0%(m/V),阳性对照组加入同浓度的葡萄糖,接种量为1%,厌氧培养24 h,每隔3 h测定发酵液OD600值和pH值,比较不同种类低聚糖对菌株生长的影响。
1.3.1.4 发酵乳的pH值及植物乳杆菌生长量的测定
在12%(m/V)脱脂乳液体中分别加入灭菌后15%、20%、25%、35%(m/V)的葡萄糖、低聚糖(低聚异麦芽糖、低聚木糖或两种低聚糖的组合物),使发酵乳中葡萄糖或低聚糖的终浓度为1.5%、2.0%、2.5%和3.0%(m/V),其中低聚糖组合物的混合比为1:1,葡萄糖组为阳性对照组。取活化后的菌株以体积分数2%接种量接种于上述脱脂乳中,37 ℃静置培养12 h后测定发酵乳活菌数和pH值。
1.3.1.5 发酵乳中菌株生长曲线
根据1.3.1.4实验结果,以最高活菌计数为指标,选取2种适宜浓度的低聚糖添加量作为菌株的生长条件,在BMRS培养基中分别加入菌种生长适宜浓度的低聚糖,静置培养0~24 h时,每隔3 h取出发酵乳,测定发酵乳活菌数和pH值,测定菌株生长曲线。
1.3.2 冷藏期发酵乳指标测定
根据1.3.1.5实验结果,选取菌株生长进入稳定期时间为发酵结束时间,并进行4 ℃贮藏,贮藏期的1 d、7 d、14 d、21 d测定发酵乳中活菌数、pH值、持水性、抗氧化活性及蛋白水解能力。
1.3.2.1 冷藏期发酵乳pH值及活菌数变化
对冷藏期的发酵乳样品(葡萄糖组为对照组)进行取样,采用梯度稀释法对植物乳杆菌ZDY2013进行活菌计数;使用电子pH计测定冷藏期各阶段样品的pH值。
1.3.2.2 持水性
持水性的测定参考文献[18]并稍作修改。称取空的10 mL离心管质量记为W0,加入5 mL成品发酵乳(葡萄糖组为对照组)质量记为W1。在3000 r/min,4 ℃的条件下离心10 min。弃去上清液,称取质量记为W2,平行测定3次。持水力计算公式如下:

1.3.2.3 抗氧化活性
(1)DPPH自由基清除能力
对发酵乳(葡萄糖组为对照组)进行8000 r/min离心10 min,再用0.44 μm滤膜过滤,获得发酵乳清;取1.5 mL DPPH溶液(0.2 mmol/L,95%甲醇作为溶剂)与0.5 mL的发酵乳清样品或95%甲醇(对照)混合均匀,在室温下反应30 min。4 ℃条件下8000 r/min离心10 min,取上清液,于517 nm处测定吸光值。DPPH自由基清除率计算公式[19]如下:

式中:
A0——95%甲醇(对照)存在时的吸光值;
A1——加入乳清样品的吸光值。
(2)ABTS+自由基清除能力
参照Perna等[20]的方法并作修改。先用95%乙醇溶液将ABTS溶液稀释到OD734=0.700±0.020(A0)。将150 μL乳清样品与3 mL ABTS溶液混合摇匀,避光反应30 min后,在734 nm处测吸光度Ai,通过添加150 μL乙醇溶液代替样品作为空白对照,测得吸光度为A0,每个样品平行测定3次。ABTS+自由基清除能力计算公式如下:
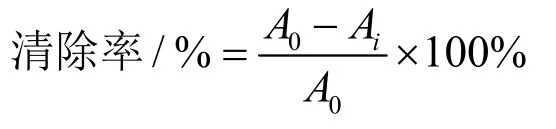
式中:
A0——未添加样品的吸光度;
Ai——添加不同样品反应后的吸光度。
(3)羟自由基清除能力
参照文献[21]所述方法测定。首先将2.5 mmol/L邻-菲罗琳溶液中,0.02 mol/L PBS(pH 7.4)和蒸馏水等体积混合,混合均匀后,加入等体积2.5 mmol/L FeSO4溶液和20 mmol/L H2O2溶液,充分混匀后,37 ℃恒温水浴1 h。根据536 nm处吸光度变化判断受试物清除羟自由基的能力。羟自由基清除率按下式计算。

式中:
Aa——发酵乳清代替蒸馏水的吸光度;
A1——混合液的吸光度;
Ab——蒸馏水代替H2O2溶液的吸光度。
1.3.2.4 蛋白水解力
邻苯二甲醛(OPA)溶液的配制参考Church等[22]的方法。OPA溶液的配制:混合100 mmol/L四硼酸钠25 mL、20%十二烷基硫酸钠2.5 mL、40 mg OPA溶解于1 mL甲醇、β-巯基乙醇100 μL,最后用去离子水稀释到体积为50 mL。OPA试剂现配现用。发酵乳样品(葡萄糖组为对照组)2.5 mL与0.75%三氯乙酸5 mL,混合均匀并静置10 min,4000 r/min离心10 min(4 ℃),收集上清液。取上清液150 μL,加入OPA试剂3 mL,混匀,室温下反应10 min后于340 nm测定吸光值。发酵乳样品A340的值与蛋白水解力呈正比例关系,值越大,说明产生的游离氨基越多,从而反映蛋白的水解程度越高。
1.4 数据处理
数据处理采用Graphpad prism 8.0软件进行分析,实验结果均为平行测定三次的值,用均数±标准差(mean±SD)表示。采用双因素方差分析(Two-Way ANOVA)的Tukey多重检验比较数据平均值的差异显著性。
2 结果与分析
2.1 不同碳源对植物乳杆菌ZDY2013体外代谢的影响
植物乳杆菌ZDY2013全基因组测序结果显示其具有多种碳水化合物代谢相关酶[23],说明其可以代谢多种碳水化合物。据研究发现,功能低聚糖代替培养基中的葡萄糖可以显著促进乳酸杆菌的生长[24,25]。选取低聚木糖、低聚异麦芽糖及其组合物改良的培养基对植物乳杆菌进行体外发酵,发现植物乳杆菌能以低聚木糖、低聚异麦芽糖及其组合物为碳源进行生长代谢,且低聚异麦芽糖优于低聚木糖(图1a)。然而,菌株进入生长稳定期的生物量均不及以葡萄糖为碳源时,这可能是由于葡萄糖是小分子碳源,易于被代谢。植物乳杆菌ZDY2013利用不同碳源代谢前3 h比较缓慢,而进入稳定期的时间均为9 h。与此结论相一致的是,发酵上清pH值在3 h内无明显变化,在9 h之后均进入稳定期(图1b)。另外本研究发现,葡萄糖为碳源时,进入稳定期的pH值最低,而低聚木糖为碳源时最高,这与代谢最终生物量高低相反,说明植物乳杆菌ZDY2013代谢能力越强,pH值越低,这可能是由于其代谢产有机酸的原因[6]。综上所述,不同碳源对植物乳杆菌ZDY2013的代谢有影响,而低聚异麦芽糖相比低聚木糖具有更强的促进代谢能力。

图1 不同碳源对植物乳杆菌ZDY2013生长代谢的影响Fig.1 The effect of carbon sources on metabolism ofL. plantarum ZDY2013
2.2 不同碳源浓度对植物乳杆菌ZDY2013发酵乳的影响
2.2.1 发酵乳中活菌数的差异

图2 不同碳源浓度对发酵乳中活菌数的影响Fig.2 The effect of concentrations of different carbon sources on the viable count ofL. plantarum ZDY2013 in fermented milk
在传统发酵乳制品中添加益生元(如功能低聚糖)来开发功能性乳制品是未来乳品行业发展新趋势。本研究发现,脱脂牛奶中添加同质量浓度的低聚木糖、低聚异麦芽糖及其组合物有利于维持植物乳杆菌ZDY2013在发酵乳中的活菌浓度(图2)。相比葡萄糖,添加1.5%和3.0%两种低聚糖的组合糖提高植物乳杆菌ZDY2013在发酵乳中的浓度最显著,分别达到9.33×108cfu/g(p<0.001)和1.05×109cfu/g(p<0.0001)。随着低聚糖添加量的增加,发酵乳中活菌数并非一直递增,尤其是单独添加低聚木糖、低聚异麦芽糖时,植物乳杆菌的浓度均在108cfu/g左右,说明低聚糖添加量与发酵乳活菌数并非呈现线性增长关系。综上结果表明,低聚木糖和低聚异麦芽糖的混合糖最适宜维持植物乳杆菌ZDY2013在发酵乳中的浓度。
2.2.2 发酵乳pH的差异

图3 不同碳源浓度对发酵乳pH的影响Fig.3 The effect of concentrations of different carbon sources on pH in fermented milk
发酵乳最终pH值可以判定发酵乳的环境是否适合乳酸菌生存。由图3可知,当糖浓度为2.0%或者2.5%时,发酵乳的pH无显著性差异,均在pH 5.20左右;当糖浓度为1.5%或者3.0%时,低聚木糖和低聚异麦芽糖的组合组pH值均低于其他3个组,其中浓度为3.0%时有显著差异(p<0.0001);随着低聚糖添加量的增加,发酵乳的最终pH值并未发生明显变化,除低聚木糖和低聚异麦芽糖的组合物浓度为3.0%,时pH为4.87,其余组pH值在5.07~5.19之间。说明低聚糖的添加量不容易影响发酵乳的最终酸碱环境,结合前期研究证实的植物乳杆菌ZDY2013具有耐受酸能力的结论[6],表明该菌株在发酵乳环境中将保持良好活力。
2.2.3 发酵乳中菌株的生长
根据图2结果,我们确定质量浓度为1.5%和3.0%的低聚糖组合物为发酵乳的适宜添加量,在此进一步对植物乳杆菌在该条件下的代谢特性进行测定,结果如图4。植物乳杆菌ZDY2013在两种条件下的生长动力学曲线相似,浓度均在12 h达到稳定,分别为7.94×108cfu/g和1.05×109cfu/g,相比体外发酵约推迟3 h(图1),说明菌株在添加低聚糖的发酵乳中生长代谢缓一点。特别的是,发酵乳的pH值均呈持续下降的趋势,且变化基本一致,说明菌株在发酵乳中持续产有机酸。

图4 植物乳杆菌ZDY2013在低聚糖发酵乳中的生长特性Fig.4 The growth characterization ofL. plantarumZDY2013 in oligosaccharides fermented milk
2.3 冷藏期低聚糖发酵乳活菌数和pH变化
上述结果表明添加功能低聚糖组合物有利于提高发酵乳中植物乳杆菌的浓度以及降低发酵乳pH值。为进一步探究这种特征可以维持多久,测定了冷藏期发酵乳中的活菌数和pH值。结果如图5a所示,无论添加的是不同浓度低聚糖组合物还是对照的葡萄糖,活菌数在冷藏期均出现下降,下降趋势类似,且在第21 d出现显著差异(p<0.0001),但是下降均不到一个数量级,这说明发酵乳在储存期有效维持了植物乳杆菌较高活菌浓度(2.34×108cfu/g以上),这一结果与虞娇娇等[13]研究结果类似,其发现添加低聚果糖的发酵乳在冷藏期对瑞氏乳杆菌MB2-1具有一定的保护作用。发酵乳pH值在冷藏期也均发生相同下降趋势,不同的是pH值在第7 d即发生显著下降(p<0.0001),随后pH值处于维持状态(图5b)。然而,添加低聚糖组合物的发酵乳冷藏期的pH值高于对照的葡萄糖(pH 4.80左右),尤其是添加1.5%组合物的组,pH值停留着5.20左右。这可能是由于在低温条件下植物乳杆菌代谢葡萄糖的能力更强,产生的有机酸更多,而添加益生元低聚糖能防止发酵乳在冷藏期的后酸化,改善其风味和质量[26]。

图5 冷藏期发酵乳中的活菌数和pH变化Fig.5 Viable bacteria counts and pH value of fermented milk during refrigerated storage
2.4 冷藏期低聚糖发酵乳持水性变化
持水性是评价发酵乳质量的一个重要指标,其与蛋白、固形物的的含量相关,良好品质的发酵乳持水性高,乳清析出少,质地均一粘稠。本研究发现,添加低聚糖的发酵乳持水性相比对照组更高,且随着冷藏时间的延长,发酵乳的持水性显著提高(图6;p<0.0001)。当低聚糖组合物添加浓度为1.5%时,发酵乳的持水性从第7 d开始一直保持在90%以上,而浓度为3.0%时,持水性略低,从第14 d开始达到约90%,这可能是渗透压差异导致的。整体而言,添加低聚糖更能改善发酵乳品质,且不一定要高浓度,这对生产实践具有一定的指导意义,添加适当浓度的低聚糖才有利于增强发酵乳持水性。发酵乳良好的持水性主要是冷藏过程中,pH值变低,小分子肽和酪蛋白增加,后者在酸性的环境下相互作用形成三维空间胶体结构,包容更多的水分子,从而提高了发酵乳的持水力[27,28]。

图6 发酵乳在冷藏期间的持水性Fig.6 Water holding capacity of fermented milk during refrigerated storage
2.5 冷藏期低聚糖发酵乳抗氧化活性变化
发酵乳具有多种益生特性,抗氧化活性是发酵乳的一项重要功能,决定其货架期和品质。图7为添加了功能低聚糖的植物乳杆菌发酵乳在冷藏期的抗氧化活性变化。添加不同浓度低聚糖组合物及对照组的发酵乳在冷藏期DPPH自由基清除率均高于60%,尤其是第7 d均达到最高90%左右(图7a)。类似的ABTS+自由基清除率也从第7 d开始,由70%~80%下降到40%~50%(图7b;p<0.0001)。这些结果与Habibi Najafi等[29]观察到含有菊粉和小麦纤维的发酵乳在冷藏期自由基清除率下降,以及李思宁等[30]在发酵乳冷藏期发现的DPPH自由基清除率的变化趋势相似。这可能是由于冷藏期具有清除这些自由基的生物活性肽发生水解和裂解导致的[29]。另外,同等碳源浓度条件下,添加低聚糖的发酵乳自由基清除能力强于添加葡萄糖的发酵乳(p<0.05),这可能与低聚糖能改善植物乳杆菌在发酵乳中的浓度和代谢产物的活性有关[10,31,32],也可能与低聚木糖本身的抗氧化活性相关[15]。不同于DPPH自由基和ABTS+自由基清除能力的变化,冷藏期发酵乳清除羟自由基的能力持续下降(图7c;p<0.0001)。相同质量浓度的碳源条件下,添加低聚糖组合物的羟自由基的清除能力整体好于葡萄糖,尤其是浓度为3.0%时的第1 d和第7 d;而浓度为1.5%时,添加低聚糖组合物的发酵乳冷藏两周后的羟自由基清除能力依然能维持70%左右,说明发酵乳中添加适量的功能低聚糖,有利于增强其抗氧化活性[30,33]。

图7 发酵乳在冷藏期间的抗氧化活性Fig.7 Antioxidant activity of fermented milk during refrigerated storage
2.6 冷藏期低聚糖发酵乳蛋白水解活性变化
乳酸菌在发酵牛乳期间,分泌的胞外蛋白水解酶会水解牛乳蛋白,释放游离氨基酸和小分子的肽,而结构稳定的抗氧化肽能发挥抗氧化活性[34]。测定添加功能低聚糖的植物乳杆菌发酵乳在冷藏期的蛋白水解活性可间接反映其抗氧化活性。实验结果如图8所示,随着冷藏时间延长,不同处理组的发酵乳蛋白水解活性持续增强,其在第14和21 d与第1 d有显著性差异(p<0.0001),推测其释放了更多的游离氨基和肽。冷藏期间,添加1.5%或3.0%低聚糖组合物的发酵乳的蛋白水解活性均高于含相同浓度葡萄糖的发酵乳,且在相同时间点,添加1.5%低聚糖组合物的发酵乳的水解活性最高,这些结果与图7抗氧化能力更强结论一致。

图8 发酵乳在冷藏期间的蛋白水解活性Fig.8 Proteolytic activity of fermented milk during refrigerated storage
3 结论
植物乳杆菌ZDY2013发酵乳具有预防乳品被杂菌污染和恢复由产肠毒素蜡样芽孢杆菌引起肠道稳态紊乱的功能。本文选用该菌能利用的低聚木糖、低聚异麦芽糖作为益生元添加在乳品中进行植物乳杆菌ZDY2013发酵,分析其对发酵乳活菌数和pH值的影响,并评价发酵乳在4 ℃冷藏期活菌数、pH值、持水性、以及抗氧化活性的变化特点。结果表明,添加低聚木糖和低聚异麦芽糖组合物的发酵乳能显著改善植物乳杆菌的活菌数,维持菌株生存的较好环境,提高其品质,增强其货架期内抗氧化活性。本研究为具有益生元特性的功能低聚糖在发酵乳中的应用,及植物乳杆菌ZDY2013的功能深度开发奠定基础。
