山黧豆160年研究历程及进展
2021-11-09徐全乐蒋景龙焦成瑾张大伟NeilTurnerShivKumar熊友才
徐全乐,蒋景龙,焦成瑾,张大伟,Neil C. Turner,Shiv Kumar,熊友才*
(1 西北农林科技大学 生命科学学院,陕西杨陵 712100;2 陕西理工大学 生物科学与工程学院,陕西汉中723000;3 天水师范学院 生物工程与技术学院,甘肃天水 741000;4 兰州大学 生命科学学院/草地农业生态系统国家重点实验室,兰州 730000;5 Centre for Plant Genetics and Breeding, the University of Western Australia, M080, Perth, WA6009, Australia; 6 International Centre for Agricultural Research in the Dry Areas [ICARDA], Rabat P.O. Box 6299, Morocco)
山黧豆(Lathyrussativus)是世界上最古老的作物之一[1],广泛分布在欧亚大陆、北美洲、南美温带地区和非洲东部等地[2-4]。由于富含蛋白、具有优良而广谱的抗逆性,山黧豆是干旱、半干旱地区和贫困地区的重要农作物选择[1],也是对抗亚太地区等存在的“隐性饥饿”、丰富农作物系统等的优良种质资源[5]。在Kew’s千年种子库及全球作物多样性信托基金主导的“气候变化的农业适应”项目中,山黧豆是29个优先级作物之一,对于物种多样性保护、针对气候变化条件下的遗传育种以及未来的粮食安全等具有重要的意义[1, 6]。然而,长期以山黧豆为主要食物来源造成的神经山黧豆中毒事件,以及传统观点对于其内源活性物质β-N-草酰-L-α,β-二氨基丙酸(β-N-oxalyl-L-α,β-diaminopropionic acid,β-ODAP)毒理特征的过度强调等限制了山黧豆的进一步开发利用。但过去一百多年中,围绕山黧豆中毒机理、β-ODAP的功能、优良品种的选育等核心问题的研究却始终没有止步。该文简要地总结了山黧豆研究160年经历的不同阶段、取得的主要成绩以及研究进展,并提出了目前存在的问题和未来研究方向,以期为今后山黧豆的种质资源开发和利用研究提供借鉴和参考。
1 山黧豆研究历程概述
在SCI引文数据库中可检索到山黧豆研究的文献记载始见于1861年。该报道记载了印度北部城市阿拉哈巴德发现的机体麻痹症状和食用山黧豆的相关性[7]。实际上,从17世纪一直到20世纪90年代,在印度、希腊、德国、俄国、孟加拉、法国、西班牙、叙利亚、意大利、阿比西尼亚、阿尔及利亚、阿富汗、埃塞俄比亚、伊朗、中国等国家发生过多次较大规模的山黧豆中毒事件[8](图1)。这些分布在不同区域和不同历史时期记载,带来了广泛的关注,使山黧豆中毒一度成为重大公共卫生问题。相关基础研究也一直以山黧豆中毒机制的研究和“低毒、无毒”品种的选育为主要核心。通过梳理,山黧豆漫长的研究历史可以粗略地划分为山黧豆中毒因素的探索、神经山黧豆中毒机理的解析、神经山黧豆中毒及β-ODAP生物学功能的再认识等3个阶段。
1.1 山黧豆中毒因素的探索
山黧豆中毒事件可以追溯到希波克拉底(Hippocrates,BCE460-370)的叙述。但直到1844年,印度科学家Sleeman[9]才对此进行了较为准确的描述。1873年,Cantani将其正式命名为Lathyrism[10],即山黧豆中毒;也可称之为neurolathyrism[11],即神经山黧豆中毒。之后,印度、埃塞俄比亚等国家发生了多起较大规模的山黧豆中毒事件(图1),也有一些文献进行了详细的报道和分析,并且发现山黧豆中毒造成的神经损伤是不可逆转的[12]。

图1 部分山黧豆中毒事件的时间轴图Fig.1 Timeline of lathyrism in partial
山黧豆不同品种引起的中毒主要表现为两种症状:一种是β-氨基丙腈(β-aminopropionitrile,BAPN)造成的骨中毒(osteolathyrism)[11, 13],一种是神经山黧豆中毒[11]。骨中毒仅占神经山黧豆中毒的12%左右[14-16]。而且,在动物饲喂实验和实际放牧中均未发现骨中毒的报道[17]。因此,科学家们主要对神经山黧豆中毒的原因进行了不懈的探索。β-氨基丙腈[18]、L-α,γ-二氨基丁酸[19]、β-腈基丙氨酸[20]等均曾被认为是山黧豆的致毒因子,但随后因为导致的症状差异及在山黧豆品种之间分布的差异性被否认。Roy等[21]利用山黧豆的乙醇提取物注射鸡后产生了严重的神经退行性症状,这与用纯化的β-ODAP[22-23]注射鸡产生的症状完全相同;说明β-ODAP是山黧豆导致的鸡神经退行性症状的主要因素。对人工合成和天然纯化的β-ODAP进行的红外光谱和核磁共振结构分析进一步佐证了β-ODAP是山黧豆中毒的因素[24]。Spencer等[25]通过人和灵长动物食蟹猴的研究证实了β-ODAP和神经山黧豆中毒的直接关联。β-ODAP的分离及其与神经山黧豆中毒的关联研究也成为山黧豆研究史上的标志性事件。
1.2 山黧豆中毒机理解析
β-ODAP的天然提取物中含有7%~10%的α构象[26],这说明β-ODAP可以发生分子重排形成α-ODAP并达到平衡[27-29]。但是,β-ODAP引发的神经山黧豆中毒具有结构的专一性,α-ODAP和D-构型β-ODAP均不导致神经山黧豆中毒[30-31]。
流行病学调查发现,神经山黧豆中毒显著影响下肢,属于上运动神经元疾病[32];发病概率受到营养水平、饮食、性别、年龄和血型等多重影响[33-34],主要在青年男子中发病[35]。
山黧豆中毒的病理学研究主要来自于鸡、成年鼠、猴等动物的神经中毒模型[36-42],偶见在人体中的报道[32]。研究表明,β-ODAP注射幼鼠脑部的葡萄糖、糖原、ATP、磷酸肌醇和乙酰胆碱等含量降低,无机磷、乳酸和乙酰胆碱酯酶水平增高[43]。而且,β-ODAP 可穿过幼年猴子等动物模型的血脑屏障,在中枢神经系统积累[37-38, 44]。
β-ODAP是谷氨酸的结构类似物,属于兴奋性氨基酸(excitatory amino acids,EAA)[45]。谷氨酸是神经系统中较普遍的兴奋性神经递质,其受体表现为N-甲基-D-门冬氨酸(N-methyl-D-asparate,NMDA)受体、海人草酸(Kainate,KA)和使君子氨酸(quisqualate,QA)受体(又称为α-氨基-3羟基-5甲基-4异噁唑丙酸受体,α-amino-3- hydroxy-5-methyl-4-isoxazole propionate receptor,AMPA受体)等3种亚型。研究表明,β-ODAP与NMDA受体的甘氨酸位点具有较弱的拮抗作用[46-47],这种作用在一定程度上表现为神经保护作用[47]。然而,绝大部分研究表明β-ODAP可通过QA和/或KA受体系统作用于神经元细胞[48-51],尤其是对于AMPA受体具有高度的亲和能力[52]。β-ODAP可以通过AMPA受体作用于大脑皮层神经系统[53-54],引起神经细胞内钙离子浓度增加[55],表现为谷氨酸类似的兴奋型神经毒性机制并导致神经退行性症状[52-53, 56]。在β-ODAP激活AMPA受体作用时伴随着谷氨酸释放到突触间隙和NO的产生,进而产生活性氧形成氧化胁迫[57]。此外,β-ODAP通过激活蛋白激酶C(Protein kinase C,PKC)调控AMPA受体磷酸化及其功能[58]或导致磷脂酰乙醇胺结合蛋白(phosphatidylehthanolamine binding protein 1,PEBP1)磷酸化水平上调[59]。PEBP1水平下调及磷酸化修饰共同激活丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号和G蛋白偶联受体信号,导致兴奋性中毒[59]。
β-ODAP还可以通过增加星形胶质细胞的谷氨酰胺合成酶活性调节中枢神经系统的谷氨酸含量[60],并通过抑制神经元附近的星形细胞的谷氨酸/胱氨酸转运体,导致神经元突触的谷氨酸含量持续增加,引起兴奋性中毒[61]。在抑制谷氨酸/胱氨酸转运体的同时,β-ODAP可以抑制胱硫醚裂解酶(cystathionine-c-lyase),导致Cys供应受到抑制,谷胱甘肽(glutathione,GSH)合成受限[62-63],引起抗氧化胁迫能力的降低。
然而,β-ODAP和突触膜只能以较低的特异性结合,不能确保β-ODAP与谷氨酸受体作用并产生效应[64]。而且,AMPA受体的广泛性、β-ODAP对不同物种作用存在的差异性[23, 45]、β-ODAP和兴奋性氨基酸(谷氨酸和天冬氨酸)之间核磁结构的有限相似性研究[65]等,均表明β-ODAP导致的山黧豆中毒除了兴奋性神经中毒机制之外可能还存在其他的作用机制。例如,代谢型谷氨酸受体I(group I metabotropic glutamatergic receptors,mGluRs)以非直接作用方式参与神经性山黧豆中毒[66]。β-ODAP还可以导致线粒体复合物Ⅰ功能紊乱[67]、线粒体内活性氧[68]和Ca2+浓度过载[69]或者天冬氨酸-α酮戊二酸转氨酶及天冬氨酸-丙酮酸转氨酶活性升高,导致慢性氨中毒[70-71]。值得关注的是,β-ODAP是酪氨酸氨基转移酶(tyrosine aminotransferase,TAT)的抑制剂,可以导致脑部多巴和多巴胺的合成增加,表现为非兴奋性作用方式[72]。
1.3 神经山黧豆中毒及β-ODAP生物学功能的再认识
1.3.1 膳食平衡与神经山黧豆中毒在灾后以山黧豆为主要食物来源的人群中,神经性山黧豆中毒的比例小于5%[73]。这种个体化的差异可能受到年龄、烹调方法、生理状态等多种因素影响[34]。而且,对于人神经性山黧豆中毒机制的理解主要来自于动物实验的推论[34]。但是,大部分β-ODAP(84%)在包括灵长类动物在内的山黧豆中毒动物模型中未进行进一步代谢[44, 74],在人体内则可被氧化为无毒的2,3-二氨基丙酸(2,3-Diaminopropionic acid,DAP)[34]。此外,通过浸泡、煮沸、煎烤等烹调手段及发酵处理等均可大幅度(90%以上)去除β-ODAP以消除山黧豆毒性[22, 75-76]。在印度监狱中日常混用4~6盎司的山黧豆数年也表现出安全性[77]。因此,对于β-ODAP及神经山黧豆中毒的理解和研究仍然需要进一步加强。
在营养均衡条件下建立的灵长类动物的山黧豆中毒模型是可逆的,说明营养不良等因素可能是不可逆神经山黧豆中毒的必要因素[78-79]。在对埃塞俄比亚228例患者的流行病调查中,首次注意到了膳食平衡和含硫氨基酸缺乏在山黧豆中毒中的作用[80]。研究表明,山黧豆与硫营养比较丰富的谷类作物混合食用可以防止中毒的发生[81-82]。山黧豆饲料中添加甲硫氨酸可防止鸡的神经中毒[83]。而且,洋葱、姜、蒜等调味品具有较好的抗氧化活性[84],可以降低氧化胁迫促进的山黧豆中毒风险[82,85]。在培养基上添加Met和/或Cys可以使神经元细胞避免β-ODAP引起的毒性[86-87]。这些结果均表明山黧豆中毒是神经毒性和营养不良的交互作用[88]。因此,膳食营养的均衡对于神经山黧豆中毒的预防至关重要[77]。这一观点直到现在仍然是适用的,是山黧豆食用和饲用的重要原则。
由于优良的综合抗逆性,山黧豆在原始农业生产体系中占据竞争优势并被贫困人群过量食用(占主粮的1/3—1/2),造成中毒。尽管它远不是有毒植物或有毒豆类的代表,但在营养科学的研究史上,从来没有一种豆科作物像山黧豆一样被钉上有毒的标签。实际上,山黧豆中毒主要发生在灾后的边远农村[89],种植户为文盲、极度贫困且长期以山黧豆为唯一食物来源[1]。当和其他豆类一样处于常规混合食用时,山黧豆毫无疑问是安全的[90]。因此,战胜山黧豆中毒主要是消除贫穷和无知[8]。
1.3.2 β-ODAP的生物学功能除了山黧豆之外,猪屎豆[91]、金合欢[92]、苏铁[93],及传统中药材人参、三七等中均含有较高水平β-ODAP(被命名为三七素,dencichine)[94-96],说明β-ODAP的存在具有广泛性,这也提示β-ODAP在植物中可能具有重要的生物学功能。例如,β-ODAP可以参与Zn2+转运[97],清除羟基自由基[98],在强光下保护植物光呼吸代谢关键酶乙醇酸氧化酶活[99]等。此外,β-ODAP水平与山黧豆的抗逆性也密切关联[100-103]。例如,干旱引起的ABA水平增加可以进一步促进β-ODAP的合成[104]。蒋景龙等[105]和宋瑶瑶等[106]分别综述了山黧豆抗逆性研究以及β-ODAP代谢与抗逆性的关系。但是,β-ODAP与山黧豆抗逆性的关联性仍需要进一步研究证实[57]。
值得关注的是,β-ODAP的神经作用具有剂量效应,在几十个微摩尔水平对于AMPA受体的过度刺激可导致神经性山黧豆中毒[86, 107];在严格控制 β-ODAP剂量(pmol)的情况下,可以激活以小脑cGMP变化为标志的神经传递而不引起神经元和动物模型的行为改变,表明该模型在神经退行性疾病研究中可能具有重要的价值[108]。
2 山黧豆基础研究的文献概况
在SCI科学引文索引数据库中以Lathyrus为主题搜索到的文章接近3 000篇,其中以Lathyrussativus或者grass pea(栽培山黧豆)为主题的文章接近2 700篇,约占总数的90%(图2,A)。这说明栽培山黧豆是该领域的主要研究对象。以20世纪60年代为拐点,β-ODAP的鉴定极大地促进了山黧豆的相关研究,发文量呈现明显增加;直至20世纪90年代,β-ODAP代谢规律的研究促使发文量呈现新一轮的激增(图2,A)。其中,以美国、印度、巴西、澳大利亚、加拿大、英国、意大利、德国、土耳其和中国等国科学家的发文量居多(图2,B)。对近30年的文献分析表明,山黧豆研究主要集中于临床神经学、神经科学、药理学和药剂学、生化与分子生物学、毒理学等领域[109],尤其在植物修复、健康促进、离体再生与遗传育种、β-ODAP与逆境响应、基因组及多组学研究等方面取得了一定的进展[110]。
在国际上,比利时根特大学Lambein教授以接近60篇的发文量位居全球山黧豆研究科学家之首(图2,C),其工作极大地推动了β-ODAP生物合成、毒理机制、流行病学、营养学等众多领域的研究。印度知名山黧豆学者Rao在山黧豆中毒机制及功能营养学研究、Spencer在山黧豆流行病学及毒理学研究、Ikegami和Kuo等在β-ODAP的生物合成及代谢研究、Siddique等在栽培生理研究、Rubiales和Patto在山黧豆遗传多样性和植物病理研究、Mehta在山黧豆中毒机制及生理生化研究等均做出了重要贡献。极大地推动了人们对于山黧豆的全方位认识,发文量均名列前茅。在国内,兰州大学植物化学家李志孝教授的发文量位居全球科学家第三,在β-ODAP的检测和生物学功能研究等方面做出了重要贡献。中国山黧豆研究先驱、兰州大学王亚馥教授等多位中国学者均位列Top50(图2,C),在接近50年的研究过程中做出了系列重要贡献(表1),使中国的山黧豆研究始终处于国际主流地位。
CNKI中收录的山黧豆研究中文文章始见于1966年(图3,A),共有40余家科研机构发表了学术论文191篇,学位论文33篇(图3,B)。其中,兰州大学的发文量遥遥领先于国内其他单位(图3,B),在一定程度上反映了兰州大学在该领域的传统优势地位。早在20世纪70年代,兰州大学陈耀祖院士就建立了国内首个山黧豆课题组,通过不断的开拓和传承,在β-ODAP的检测分析和生理生态学功能等研究方面形成了鲜明的特色。此外,中国农业大学围绕林生山黧豆的引种栽培、丁二氨酸的代谢等展开研究,西北农林科技大学聚焦于β-ODAP的生物合成途径研究和种质资源利用基础研究,其发文量均位于国内三甲(图3,B)。南充市农业科学院聚焦于山黧豆作为绿肥的推广应用研究,中国农科院在山黧豆遗传多样性的研究等方面也形成了一定的传统和优势。
3 目前的研究进展
与谷物相比,对于豆类在饮食和营养方面的学术研究要少很多,主要集中在豆类的抗营养因子和毒性研究[111]。对于毒理特征的过度强调,使得山黧豆的种质资源利用长期处于低谷。但在相关学者的长期坚持下,也取得了较为丰硕的成果。
3.1 山黧豆毒理学研究
流行病学调查和细胞实验表明,神经山黧豆中毒与含硫营养的缺失呈正相关[81, 87],但其细胞及分子机理尚不明确。由于β-ODAP与Glu是结构类似物且均为兴奋性氨基酸,β-ODAP可能和Glu 竞争性结合Glu 受体AMPA并影响Ca2+通透性、神经元的发育等[112]。兴奋性神经中毒机制也是目前接受程度最高的神经山黧豆中毒机制。
AMPA 受体阻滞剂1-萘基乙酰基-精胺(1-naphthyl acetyl-spermine,NAS)可以特异性地抑制β-ODAP 诱导的细胞内[Ca2+]升高[68]。这进一步说明β-ODAP可结合并异常激活神经细胞的AMPA受体,破坏神经细胞内钙稳态。并使得Integrin β1等表达量上调,黏着斑激酶磷酸化增加,黏着斑蛋白单元在细胞微丝上大量聚集影响细胞骨架聚合,最终引起神经细胞营养障碍[113-114]。利用蛋白组学分析也表明,参与细胞骨架结构与信号传递、细胞代谢、自由基清除、氧化胁迫、神经退行性疾病的多个蛋白在β-ODAP处理2 h后表达量上调[115]。
3.2 β-ODAP分析方法
山黧豆种子的β-ODAP含量介于0.1%~2.5%[116],其检测分析是山黧豆研究的核心方法之一。在化学合成β-ODAP标准品的基础上[117-118],建立了多种β-ODAP的检测方法,为β-ODAP含量分析及毒理机制研究奠定了坚实的基础。多篇文献曾详细综述了β-ODAP检测方法[119-120],包括比色法[121],基于PITC、FDNB、6-aminoquinolyl-N-hydroxysuccinimidyl carbamate (AQC)等衍生的多种HPLC检测方法[122-127],氨基酸分析仪法[128],毛细管电泳法[129-130],流动注射法[131],液相-质谱法[96]等。近来,Emmrich等[132]利用稳定同位素标记的β-ODAP为内标建立了质谱分析方法,成为了β-ODAP分析检测的金标准。此外,BIA[133]、草酸[134]等β-ODAP生物合成关键前体检测方法的建立,为β-ODAP代谢规律的研究奠定了基础。
3.3 山黧豆品质改良基础研究
3.3.1 β-ODAP生物合成研究β-ODAP在山黧豆不同组织和器官中均有分布,其中在幼苗和发育种子中含量最高[135-136]。研究表明,山黧豆种子β-ODAP的含量主要受到遗传背景的影响,但环境因素也有一定作用[15, 137-138]。Jiao等[139]全面综述了β-ODAP含量的影响因素。例如,β-ODAP随山黧豆生长阶段呈现规律性变化[136]、水分胁迫增加β-ODAP含量[140]、盐胁迫降低β-ODAP含量[141]等。此外,β-ODAP含量与N、P、金属离子等多种元素相关[106, 139, 142-143]。
β-ODAP的生物合成途径研究是山黧豆的核心研究内容之一,对于山黧豆的种质资源利用,三七、西洋参等含有β-ODAP物种的品质形成机理研究等具有重要意义。经过多位科学家的持续研究和验证[100, 144-151],提出了广为接受的β-ODAP生物合成途径。即环状分子异噁唑-5-酮(Isoxazolin-5-one)在半胱氨酸合成酶(cysteine synthase,CS)催化下与O-乙酰丝氨酸(O-acetylserine,OAS)反应生成重要中间产物β-异噁唑-5-酮-丙氨酸[β-(isoxazoline-5- on-2-yl)-L-alanine,BIA][100, 150-152];BIA开环后接受草酰辅酶A的草酰基而最终形成β-ODAP[100, 153]。然而,在该合成途径中,CS虽然能够催化合成BIA,但其效率较低,仅为CS催化Cys合成效率的十分之一不到[150];而且BIA开环后的产物L-α,β-二氨基丙酸(L-α,β-diaminopropionic acid,DAP)尚未检测到,催化DAP形成β-ODAP的关键酶尚不清楚。
山黧豆β-腈基丙氨酸合成酶(β-cyanoalanine synthase,β-CAS)由于蛋白序列及功能与 CS高度类似,也被划分到CS家族[154-155]。通过种子萌发期的CS、β-CAS活性检测、BIA含量和β-CAS基因的组织表达分析等发现,β-CAS在β-ODAP的生物合成中起关键作用[155-158]。该酶表现为β-CAS和CS两种酶活,是磷酸吡哆醛(pyridoxal phosphate,PLP)依赖的双功能酶[155, 158];可与丝氨酸乙酰基转移酶(serine acetyltransferase,SAT)形成半胱氨酸合成酶调控复合物CRC(cysteine regulatory complex)起到β-ODAP生物合成分子开关的作用[158]。在硫充足情况下,山黧豆β-CAS 和LsSAT2形成CRC调控复合物,增强了SAT活性;充足的硫供应保证了Cys的合成,进而在游离β-CAS作用下合成BIA及β-ODAP。在低硫情况下,OAS的积累促使CRC解聚,降低了SAT活性;低水平的Cys限制了以其为底物的BIA及β-ODAP合成。但积累的OAS可在CS催化下和异噁唑-5-酮以较低效率合成BIA[158]。BIA在体内可以自发开环形成DAP。利用烟草瞬时表达系统进行验证表明,DAP可在BAHD3酰基转移酶的作用下和草酰辅酶A反应生成β-ODAP[159]。本课题组利用原核表达系统获得BAHD3后,与豌豆粗提液进行反应也检测到了β-ODAP的生成(数据待发表)。这说明BAHD3确实是催化BIA合成β-ODAP的关键酶。
长期以来,三七等植物中的β-ODAP研究均参照、引用山黧豆中的相关研究结果[160]。最近,由于三七染色体级别基因组的组装,发现三七中可能存在有别于山黧豆的β-ODAP合成路径[161]。其主要依据之一是基于代谢组分析在三七中未检测到BIA及其前体物质异噁唑-5-酮[161]。然而,这一结果尚需进一步验证。一是在山黧豆基因组已公布的情况下,该潜在通路中发掘的β-ODAP生物合成关键酶,如3-phosphoserine aminotransferase (PSAT)、Class II PLP-dependent enzymes(PALP)、ornithine cyclodeaminase(OCD)等应与山黧豆基因组[162]、转录组[163]等数据进行比较,以明确山黧豆中是否也存在相同代谢途径。二是应进一步通过质谱等方法确认BIA及异噁唑-5-酮在山黧豆、三七、人参、西洋参等物种中的含量。因此,山黧豆仍然是β-ODAP生物合成研究的模式生物。
3.3.2 蛋白酶抑制剂等抗营养因子研究含硫氨基酸缺乏及存在过敏原蛋白、蛋白酶抑制剂等抗营养因子是山黧豆品质改良研究的主要方面[106]。山黧豆过敏可导致支气管哮喘、过敏性鼻炎、面部浮肿和荨麻疹等症状[164-167]。但山黧豆过敏原蛋白的研究相对较少,在过敏原蛋白数据库(allergome)中仅收录了4种山黧豆过敏原蛋白,其中Lat S 来自于栽培山黧豆。Qureshi等[167]鉴定了一个山黧豆过敏原蛋白2S albumin,与豌豆2S albumin序列相似性达到了85%。Xu等[168]利用花生过敏患者血清确认了花生与山黧豆的交叉过敏反应,并纯化和鉴定了山黧豆47 kD过敏原蛋白β-lathyrin。山黧豆β-lathyrin与豌豆vicilin 、扁豆Len c 1 相似性较高,其抗原表位T8在花生、大豆、扁豆、绿豆等多种豆科作物中广泛存在。进一步证实了山黧豆与花生、扁豆等存在的交叉过敏关系[168-169]。
胰蛋白酶抑制剂(trypsin inhibitor,TI)是大豆等豆科植物中重要的抗营养因子,可分为KTI(Kunitztrypsin inhibit)和BBI(Bowman-Birkproteaseinhibitor)两大类[170]。在山黧豆中也分别检测到了KTI和BBI活性[171-174]。研究表明,山黧豆TI易溶于异丙醇[175],其活性介于大豆和蚕豆等其他食用豆之间[17]。Xu等[176]对不同地域来源的51个山黧豆样本进行了抗营养因子KTI和BBI的生化特征分析,筛选到了1个巴西山黧豆(PI337007)和2个塞浦路斯山黧豆品种(PI206375,PI283567)具有较低的KTI和BBI活性,为进一步的品种选育奠定了基础。实际上,在将山黧豆作为猪等动物饲料时,抗营养因子KTI和BBI的影响远远大于β-ODAP[177]。山黧豆β-ODAP的生物合成与硫代谢密切相关,而过敏原蛋白β-lathyrin可能受到硫代谢关键化合物OAS的调节,抗营养因子KTI和BBI也富含二硫键[106]。因而,研究硫代谢对于山黧豆含硫氨基酸、β-ODAP及抗营养因子KTI和BBI合成的调控,进而筛选低抗营养因子种质将是山黧豆品质改良研究的重要内容。
3.3.3 山黧豆种质资源筛选及品种改良伴随着神经山黧豆中毒机制的研究,低β-ODAP品系的选育一直都是山黧豆育种的重要方向。其中,种质资源评价和筛选、传统育种、突变体育种、体细胞突变、转基因育种和农田综合管理等多种手段均可起到重要作用[178]。熊友才等[179]曾进行了细致的综述。
栽培山黧豆具有极其丰富的农艺品质变异,通过品质资源筛选有望获得优良品系[180]。例如,Roy等[181]通过筛选获得了低β-ODAP品系P-24,并广泛栽培。陈耀祖等[182]从65个品种中筛选到5个低毒高产、抗旱耐寒的栽培山黧豆品种,并用于动物饲料。此外,‘西选1号’、‘南选山黧豆’等也是国内筛选的优良品种(表1)。通常,传统杂交育种在作物育种中占据主体地位。但是,栽培山黧豆和除L.Amphicarpos、L.cicera之外的山黧豆种内杂交不亲和[183],与豌豆等物种的种间杂交不亲和[184],这极大地限制了传统杂交技术的应用。山黧豆和豌豆原生质体融合是一条可能的途径[184]。中国科学家曾用60Coγ射线[185]、C6+重离子[186]、60Coγ射线与EMS复合处理[187]等对山黧豆种子进行了诱变剂量和诱变效果的探索。近年来,Emmrich[159]利用EMS诱变获得了3个低β-ODAP品系,并在非洲进行推广。
山黧豆离体再生体系的建立[188-194]是基因工程育种的前提条件。研究表明,再生植株[195]和体细胞突变株[196-197]的β-ODAP含量均显著降低。此外,针对β-ODAP生物合成途径相关基因,利用山黧豆遗传转化体系[198-199],有望获得低β-ODAP品系。例如,在山黧豆中过表达草酸脱羧酶基因,在降低草酸含量(75%)的同时降低了β-ODAP含量(73%)[200]。此外,DNA分子标记被广泛用于山黧豆遗传育种和多样性研究等[201]。
长期以来,山黧豆品种选育都是以低β-ODAP含量为首要目标。随着对神经山黧豆中毒机制和β-ODAP生物学功能研究的深入理解,山黧豆育种工作也向低β-ODAP含量兼具早花、大籽粒、高产、高收获指数等优良农艺性状发展[137]。此外,山黧豆具有极强的抗旱、抗病性,可以作为豌豆等其他作物抗逆遗传育种的基因库。
3.4 山黧豆种质资源利用研究
少数主流作物的盛行以及依赖单一作物作为主食的饮食习惯造成了很多高品质作物的资源浪费,也影响到人们的食品安全和人口健康。在联合国粮农组织提出的零饥饿计划(Zero Hunger in Asia and the Pacific)中,未来智慧食物(Future Smart Food,FSF)代表了农业生产及消费的新方向[5]。FSF又被称为“被忽视和未充分利用的作物”(neglected and underutilized species,NUS)、“孤儿作物”(orphan crops)、“有希望的作物”(promising crops)等,可以提供丰富的营养元素、具有一定气候适应性、低成本以及易栽培等特征。山黧豆因为较高的蛋白含量、广谱的抗逆性以及对环境友好等特征而名列其中[5]。因此,进一步筛选山黧豆种质资源,并在干旱、半干旱地区以及待发达地区的推广应用具有广阔前景。
作为绿肥和饲料也是目前山黧豆种质资源的主要利用方向。山黧豆作为绿肥常见于澳大利亚、加拿大以及中国四川、贵州、湖北、河北等地。在加拿大进行的研究表明,山黧豆代替休耕方式和小麦轮作,可以显著提升土壤有机氮等理化性质[202-203]。虽然山黧豆和豌豆、扁豆等豆科绿肥在土壤水分、有效氮及有效磷等的改善中差异不大[204],但更适合半干旱土壤和贫瘠土壤[205]。韩文斌等[206]比较了山黧豆不同播期和播量对橘园间作下山黧豆产量及农艺性状的影响,对橘园山黧豆间作高产栽培措施进行了分析和评价[207]。而且,在橘园中播种山黧豆等作为豆科绿肥以起到保水、降温、改土肥田作用已明确写入中国农业农村部发布的《果菜茶有机肥替代化肥技术指导意见》中,并和苕子、箭筈豌豆等一起作为长江上游柑橘带、赣南-湘南-桂北柑橘带、浙-闽-粤柑橘带“绿肥+自然生草”模式的主推品种。
在饲料应用方面,Hanbury等[17]对山黧豆饲用的安全性和可能性做了详细的综述;可用于鱼、鸟、猪、牛、羊等[17, 208],也可和大麦等混合作为青贮饲料[209]。陈耀祖院士等曾筛选到低β-ODAP山黧豆(0.075%~0.093%)。在甘肃武威的种植和动物实验表明,β-ODAP含量稳定,在饲喂驴、猪、羊(日饲喂量50%~80%)超过半年未发现急性或慢性山黧豆中毒[180,182]。李凤民等[210]曾将山黧豆与苜蓿等制备为混合饲料用于兔等啮齿类动物。山黧豆饲料尤其适合于反刍动物。在反刍动物中没有山黧豆中毒的报道,可能是反刍动物的肠、瘤胃中具有降解β-ODAP的微生物埃氏巨球型菌(Megasphaeraelsdenii)、双酶杆菌(Clostridiumbifermentans)等[211-212]。这也表明可采用复合菌剂处理提高山黧豆饲草安全性。
3.5 学术会议和学术组织
自1985年首次召开山黧豆国际会议后,在法国、英国、埃塞俄比亚、孟加拉、比利时等国家举办了至少10次国际性专题研讨会[179]。其中,2004年在叙利亚召开的国际会议(International Workshop on Grasspea as a Food/Feed Crop)上探讨了山黧豆食用和饲用的安全性。2009年在比利时召开的国际会议(International Workshop on the Toxico-Nutritional Neurodegenerations Konzo and Lathyrism)上对于神经山黧豆中毒与营养均衡之间的关系等进行了深入探讨,为山黧豆的安全利用奠定基调[179]。
为了加强信息交流,促进国际合作研究,山黧豆研究相关人员发起了国际山黧豆提升和山黧豆中毒消除合作组织(International Network for the Improvement ofLathyrussativusand the Eradication of Lathyrism,INILSEL)、木薯氰化物中毒和神经山黧豆中毒合作组织(Cassava Cyanide Diseases &Neurolathyrism Network,CCDNN)等,建设了Lathyrus Lathyrism Newsletter和CCDNN News等山黧豆专业通讯期刊,极大地促进了国际山黧豆研究专家的交流与合作。近期,由英国詹姆斯·赫顿研究所(the James Hutton Institute)、全球作物多样性信托基金(the Crop Trust)和国际干旱地区农业研究中心(ICARDA)联合发布的山黧豆网站(www.grasspea.net),将致力于整合现有山黧豆信息和资源,促进山黧豆合作研究。
中国的山黧豆研究长期呈现发散态势。自2018年以来,中国山黧豆科研人员的交流逐渐增加,以兰州大学为牵头单位成立了甘肃省山黧豆产业技术创新战略联盟,并于2021年初在陕西杨陵召开了全国性的山黧豆生物学学术研讨会,产生了热烈的思想碰撞,极大地促进了山黧豆研究学者之间的交流,有力地推动了中国山黧豆高水平基础研究和推广应用研究(表1)。

表1 中国山黧豆研究进程中的重要事件
4 存在的问题和未来研究方向
近年来,山黧豆相关基础研究和推广应用在经过短暂的高潮后,目前处于深入发展的瓶颈期,但前景仍然十分广阔。

A. 不同年份的发表情况;B.不同国家的发表情况;C. 发表文章数量前50名的科学家图2 山黧豆研究发表的SCI 论文情况A. Papers published in different years; B. Papers published in different countries; C. Top 50 scientistsFig.2 Distribution of Lathyrus-related SCI papers
4.1 山黧豆分子生物学等基础研究亟待强化
基因组学及代谢组学等是解析豆科作物天然产物生物合成途径的关键技术。在分子生物学、基因工程等研究手段层出不穷、飞速发展的今天,山黧豆的分子生物学研究处于相对滞后的状况。Emmrich等[162]虽然获得了山黧豆的基因组草图,并利用BUSCO(Benchmarking Universal Single-Copy Orthologs)评估显示基因组组装完整度超过80%;但山黧豆较大的基因组使得染色体级别的组装仍然面临巨大挑战。近年来,转录组学[163]、代谢组学[213]等纷纷用于β-ODAP的合成通路研究,蛋白组学[214]、转录组学[215]、代谢组学[216]等用于山黧豆抗脱水胁迫的分子机制研究,而转录组学在山黧豆遗传多样性研究中也广泛采用[217]。但是,β-ODAP生物合成关键基因的功能研究和山黧豆抗逆分子机制研究等仍然相对匮乏,严重影响了对于山黧豆优良生物学性状的理解和进一步开发利用。

续表1 Continued Table 1
此外,蓬勃发展的CRISPR基因编辑技术在水稻、小麦等作物基因功能研究及遗传改良中的应用已经非常广泛。Zhang 等[218]综述了CRISPR在作物定点改造及精准育种中的重要应用,并对利用CRISPR加速作物的遗传改良解决粮食短缺问题进行了展望。但是,山黧豆高效离体再生和转化体系的不成熟限制了CRISPR技术的应用。VIGS(Virus induced gene silencing)技术是豆科作物基因功能研究的重要工具[219],在栽培山黧豆[220]中的成功应用将有利于基因功能的深入研究。
4.2 山黧豆种质资源利用急需系统规划
种质资源是农业科技原始创新、现代种业发展的物质基础。法国国家植物保护机构(CBNPMP)和国际干旱地区农业研究中心(ICARDA)分别保存了4 477份和3 239份山黧豆种质资源。印度和孟加拉等传统山黧豆食用国家也分别保存了2 000份左右。其中,栽培山黧豆品种占据一半以上[221]。中国的山黧豆种质资源收集呈散发状态,兰州大学、西北农林科技大学、中国农科院、甘肃农科院、榆林农科院、南充市农科院、江苏农科院等均有收集保存。此外,国家种质库共收集引进山黧豆属种质资源476份,包含中国内种植的栽培种有36份。
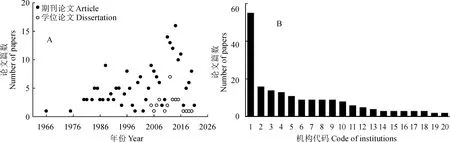
A. 不同年份的发表情况;B. 发表文章数量前20名的机构:1.兰州大学;2.中国农业大学;3. 西北农林科技大学;4.南充市农业科学研究所(现南充市农业科学院);5.东北师范大学;6.中国农业科学院中兽医研究所(现兰州兽医研究所); 7.中国农业科学院农业资源与农业区划研究所;8.西北民族学院(现西北民族大学);9.天水师范学院;10.陕西理工学院(现陕西理工大学);11.华中农业大学;12.甘肃省农业科学院土壤肥料研究所(现甘肃省农业科学院土壤肥料与节水农业研究所);13.贵州大学;14.四川大学;15.贵州省农业科学院土壤肥料研究所;16.中国科学院近代物理研究所;17.黑龙江八一农垦大学;18.哈尔滨师范大学;19.甘肃农业大学;20.中国农业科学院作物科学研究所图3 山黧豆研究发表的中文论文情况A. Papers published in different years; B. Top 20 institutions: 1. Lanzhou University; 2. China Agricultural University; 3. Northwest A&F University; 4. Nanchong Academy of Agricultural Sciences; 5. Northeast Normal University; 6. Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (CAAS); 7. Institute of Agricultural Resources and Regional Planning, CAAS; 8. Northwest Minzu University; 9. Tianshui Normal University; 10. Shaanxi University of Technology; 11. Huazhong Agricultural University; 12. Institute of Soil, Fertilizer and Water-saving Agriculture, Gansu Academy of Agricultural Sciences; 13. Guizhou University; 14 Sichuan University; 15. Soil and Fertilizer Institute, Guizhou Academy of Agricultural Sciences; 16. Institute of Modern Physics, Chinese Academy of Sciences; 17. Heilongjiang Bayi Agricultural University; 18. Harbin Normal University; 19. Gansu Agricultural University; 20. Institute of Crop Sciences, CAAS.Fig.3 Lathyrus-related papers published in Chinese
山黧豆种质资源仍然处于粗放式利用阶段,缺乏系统规划和深度挖掘。目前,定西等地主产栽培山黧豆,用于土地倒茬轮作或被收购用于鱼料、鸟料配伍;宁夏局部仍较大规模采用栽培山黧豆作为牛羊饲料;四川等地主产扁荚山黧豆,广泛用于绿肥等。实际上,山黧豆蛋白含量高、综合抗逆性好,可在海拔1 800 ~ 2 800 m的高寒地带生长[222],针对青海等高海拔地区豆科牧草缺乏现状,对山黧豆品种展开筛选工作非常必要。此外,山黧豆根系发达、耐贫瘠,对于镉、铅等重金属污染土壤具有较好的修复作用[223-226],可以应用于荒漠土壤改良、矿山土壤修复等。而且,根瘤菌可以显著提升山黧豆的铅耐受能力[227]。
在最近召开的全国山黧豆生物学学术研讨会上,西北农林科技大学山黧豆课题组提出了山黧豆的综合利用方案,认为山黧豆在食用、饲用、观赏、药用、土壤改良等多方面均具有进一步深入挖掘的潜力(图4)。目前,该课题组联合甘肃农科院西北种质资源保存与创新利用中心,通过对山黧豆种质资源进行精准鉴定和评价筛选,进一步推动山黧豆种质资源保存分库(圃)建设,为西部农业生态的安全保障提供有价值的种质资源。

图4 山黧豆种质资源利用Fig.4 Utilization of Lathyrus germplasm
4.3 山黧豆作为功能性食品的探索前景广阔
过量食用山黧豆导致神经中毒的发生,促使印度等国家政府在20世纪50、60年代颁布的防止食品参杂条例(Rule 44-A of the Prevention of Food Adulteration Rules)中限制了山黧豆的大面积销售和使用。然而,包括印度、孟加拉、葡萄牙、中国等多个国家的干旱半干旱地区仍然零散种植并食用山黧豆。印度流行病调查显示,在营养均衡条件下食用山黧豆长达34年并未引起山黧豆神经中毒发生[228-229]。孟加拉的调查也发现,山黧豆在食物中的占比小于1/3的情况下食用是安全的[230]。近30年以来,未见到山黧豆中毒事件的报道,说明减少食用范围和多元化饮食完全可以避免山黧豆中毒发生。实际上,波兰在1997年就重新开放了山黧豆作为农作物的品种登记(1960年曾移除)[231]。印度医学研究理事会(indian council of medical research,ICMR)也于2016年解除了对于山黧豆的禁止条例[232],并在发布的2020年食品安全和标准修订草案中将禁止和限制销售山黧豆修订为山黧豆在食物中的添加量不超过2%。这为山黧豆种质资源的重新利用和加工带来了曙光。
在解析神经山黧豆中毒因素的同时,科学家们也在关注山黧豆自身的营养价值[233-234]。例如,山黧豆种子含有41%淀粉和17%膳食纤维(2%可溶,15%不可溶),适合添加到食物中以增加饱腹感和延迟血糖反应[235];较高的膳食纤维有利于心血管疾病治疗[236]。实际上,山黧豆研究国际知名专家Rao[237]和Lambein[1]等均认为山黧豆种子含有多种潜在功能成分。对这些功能成分及其药理活性研究表明,山黧豆作为新型功能食品具有重要的开发价值[238]。例如,山黧豆油脂含量约2%,其中亚麻酸等ω3脂肪酸及其他多不饱和脂肪酸可达60%[239-240],从而使山黧豆具有潜在的降血脂、抗血栓形成、抗炎等诸多药用功效功能[241]。山黧豆种子含有的肌醇磷酸聚糖(inositol phosphoglycan,IPG)具有类胰岛素活性,是山黧豆用于减缓糖尿病症状的主要原因[242]。富含的赖氨酸[243]可以作为膳食营养平衡的重要来源。而且,山黧豆是已知作物中高精氨酸(HA)的唯一膳食来源[1],这进一步提升了山黧豆的营养利用价值[244-246],因为HA可以作为酶底物持续产生和维持内源性NO水平,引发血管扩张等。甚至,低血浆HA水平可作为心脏猝死的独立预测因子[247]。即使对于山黧豆中备受关注的β-ODAP而言,低剂量对于神经细胞也具有一定的保护作用,并可用于制备治疗神经变性病药物[248]。而且,人成神经母细胞瘤的体外研究表明,β-ODAP对阿尔兹海默症的治疗可能具有一定的作用[90]。
致谢:感谢兰州大学李志孝教授在成稿过程中的仔细阅读并提出宝贵意见!感谢王亚馥教授、李志孝教授、王崇英教授等山黧豆研究先驱的不懈努力和长期坚守,使山黧豆研究的良好传统得以传承!感谢全国山黧豆研究同仁和甘肃省山黧豆产业技术创新战略联盟的长期支持!
