不同生态环境条件沙生植物沙鞭的结实研究
2021-10-16刘峰马子兰刘涛吕婷梁瑞芳苏旭周勇辉刘玉萍
刘峰 马子兰 刘涛 吕婷 梁瑞芳 苏旭 周勇辉 刘玉萍


摘 要:為了阐明不同生态环境条件下沙生植物沙鞭的结实规律,该文对沙鞭137个种群结实情况进行了实地观察,发现沙鞭种子的结实情况可被划分为无种子、种子饱满和种子不饱满3种类型;在此基础上,该文采用聚类分析、Kruskal-Wallis检验和典范对应分析(CCA)等方法探究沙鞭种群结实情况与22个地理气候因子的相关性。结果表明:(1)沙鞭137个种群按照地理气候因子不同聚为3个组;(2)Kruskal-Wallis检验显示沙鞭3个组间种子结实情况差异不显著(P=0.269),即沙鞭种群间种子结实与其所处的地理气候因子无直接相关性;(3)典范对应分析(CCA)表明沙鞭种群间种子结实情况差异也不显著(P>0.05),但地理气候因子与种子饱满度以及无种子特征具有显著相关性,其中海拔和降雨因子(bio12-bio19)与种子饱满度呈正相关,而经纬度和温度因子(bio4,bio7)与种子饱满度呈负相关,无种子特征仅与最湿季平均温度(bio8)呈正相关。地理气候因子对沙鞭天然种群有性繁殖(有种子)重要性高于无性繁殖(无种子),表明制约沙生植物沙鞭有性繁殖的环境因子复杂,其无性繁殖可能是种群数量稳定的适应性表现。
关键词:禾本科,沙生植物,沙鞭,结实,非参数检验,典范对应分析
中图分类号:Q944.59
文献标识码:A
文章编号:1000-3142(2021)09-1457-08
Abstract:In order to clarify the fruiting regularities of Psammochloa villosa in different ecological environments,we observed the fruiting of 137 populations of P. villosa,and found that the fruiting could be divided into seedless,filled and unfilled seeds. Based on them,we analyzed the correlation between the populations of P. villosa and 22 geographical and climatic factors by applying cluster analysis,Kruskal-Wallis test and canonical correspondence analysis (CCA). The results were as follows:(1)A total of 137 populations of P. villosa were clustered into three groups according to different geographical and climatic factors. (2)Kruskal-Wallis test showed that there was no significant differences in fruiting among three groups of P. villosa (P=0.269). That is,the fruiting of populations was not direct related with geographical and climatic factors. (3)Meanwhile,the CCA analysis showed that there was no significant differences in fruiting among populations (P>0.05). However,there was a significant correlation between geographical and climatic factors and plumpness of seeds and seedless feature. Among them,altitude and bio12-bio19 were positively correlated with filled seed,while longitude,latitude,bio4 and bio7 were negatively correlated with filled seed,but the seedless feature was only positively related to the mean temperature of wettest quarter (bio8). In summary,geographical and climatic factors are more important to the sexual reproduction (with seeds)than the asexual reproduction (without seeds)in the natural population of P. villosa,which proves that the environmental factors that restrict the sexual reproduction of P. villosa are complex,and its asexual reproduction may be a stable adaptive performance of the population.
Key words:Poaceae,psammophytes,Psammochloa villosa,fruiting,non-parametric test,canonical correspondence analysis (CCA)
沙鞭(Psammochloa villosa)是禾本科(Poaceae)、针茅族(Stipeae)、沙鞭属 (Psammochloa)的一种多年生草本植物,主要分布于我国内蒙古、甘肃、新疆、青海、陕西北部等地,通常生长在海拔为910~2 900 m的沙丘上(郭本兆,1987)。沙鞭是一种克隆植物,能够通过根茎快速繁殖,适应流动沙丘的生存环境(马毓泉,1994)。作为沙地植物群落的优势种,沙鞭具有较强的耐寒、耐旱及抗病和抗风沙能力;花序相对粗长、穗多粒大,是优良的牧草资源(马毓泉,1994;吕婷等,2018a)。因此,沙鞭不仅具有重要的生态价值,而且具有重要的经济价值。
迄今为止,国内外诸多学者对沙鞭的外部形态特征(马毓泉,1994;吕婷,2019)、叶表皮微形态特征(吕婷等,2018b)、地理分布及种质资源(刘瑛心,1985;马毓泉,1994;吕婷等,2018a)、种子萌发特性(黄振英,2003;黄振英等,2005)、克隆特性(董鸣等,1999)及遗传多样性(王可青等,1999; Li et al.,2001)等进行了较为全面的研究。例如,吕婷(2019)通过对沙鞭20个种群的12个表型性状(如旗叶长度和长宽比、花序长度、颖脉数等)作了统计分析,发现沙鞭外部形态具有丰富的表型多样性,并且绝大多数表型性状均存在不同程度的变异,尤其种群间变异明显大于种群内变异。吕婷等(2018b)在群体水平上对沙鞭的叶表皮微形态解剖特征进行了探讨,发现沙鞭的叶表皮结构为典型的狐茅型,脉间长细胞形态和壁波曲程度、短细胞形态和分布式样、气孔器体积大小与分布及副卫细胞性状、刺毛形态等在种群间有明显差异,具有重要的分类价值;同时,还指出沙鞭不同种群的叶表皮气孔密度存在差异,这是对荒漠环境的一种微观适应。吕婷等(2018a)通过对沙鞭野外自然地理分布区的全面调查,采集到沙鞭60个种群、450个个体的植物标本、DNA材料和成熟种子。就种子萌发特性而言,黄振英等(2005)发现0.5~2.0 cm浅层沙埋时,沙鞭种子萌发和幼苗出土率最高;当沙埋深度超过2 cm时,种子萌发和幼苗出土率随沙埋深度增加而降低。在克隆特性方面,董鸣等(1999)研究表明,沙鞭的克隆整合和生长特性对其生存能力有极其重要作用。王可青等(1999)采用等位酶技术对沙鞭4个天然种群的遗传结构和克隆多样性进行了探讨,发现沙鞭在同类单子叶植物中具有相对较低的遗传多样性,且种群间遗传变异大于种群内遗传变异。Li et al.(2001)利用ISSR分子标记对沙鞭7个种群的遗传变异做了分析,发现沙鞭种群的遗传变异水平较高,并且种群间遗传变异大于种群内遗传变异,这再次证实王可青等(1999)研究结果的正确性和合理性。达尔文广义适合度中指出,适合度是衡量一个个体存活和繁殖成功机会的尺度,适合度(直接、间接适合度)越大的物种群体中个体繁殖成功的机会就越大(刘鹤玲等,2007),然而,关于不同生态环境条件下沙生植物沙鞭结实情况的研究目前尚属空白。据此本研究以内蒙古高原及其毗邻地区的沙鞭种群作为研究对象,通过全面的野外调查和室内观察研究不同生态环境条件下沙鞭的结实情况,分析不同生态环境条件对沙鞭种群适合度的影响及其原因,旨在为今后沙鞭播种育苗、优良种质选育、种质资源合理开发利用提供理论依据,并为沙漠化环境治理中优良禾草筛选提供基础资料。
1 材料与方法
1.1 材料和数据获取
自2016年以来,我们连续4年对内蒙古高原及其毗邻地区自然分布的沙鞭种群的结实情况进行实地调查,调查点涵盖其主要天然分布区,同时详细记录调查点经度、纬度及海拔高度等地理因子,共获取137个不同生态环境条件下沙鞭种群的结实情况,每个种群观察10个个体,发现沙鞭种子结实可被划分为无种子(种群内全部无种子)、种子饱满(种群内种子全部饱满)和种子不饱满(种群内种子全部不饱满)3种类型(图1)。同时,本研究从全球WorldClim数据库1970—2000年年均气候数据库(https://www.worldclim.org/)提取沙鞭种群调查点年均温(bio1)、温度日偏差(bio2)、等温性(bio3)、温度季节性偏差(bio4)、最暖月最高温度(bio5)、最冷月最低温度(bio6)、年均温偏差(bio7)、最湿季平均温(bio8)、最干季平均温(bio9)、 最暖季平均温(bio10)、最冷季平均温(bio11)、年均降雨量(bio12)、最湿月降雨量(bio13)、最干月降水量(bio14)、降雨量变异系数(bio15)、最湿季降雨量(bio16)、最干季降雨量(bio17)、最暖季降雨量(bio18)和最冷季降雨量(bio19)等19种气候因子变量,并在地理空间云数据网站(http://www.gscloud.cn/)下载底图,最后运用ArcMap 10.5软件绘制沙鞭种群分布图。
1.2 研究方法
首先,将沙鞭137个种群按照地理气候数据进行标准化处理,通过PCA聚类分析形成群体间环境变量分组;然后,根据分组结果对沙鞭种群种子的结实情况作Kruskal-Wallis检验,探讨沙鞭种群间结实情况(0表示无种子、1表示种子饱满、2表示种子不饱满)是否存在差异,以及结实情况是否与地理气候因子存在相关性;最后,对沙鞭种群种子的结实情况与其地理气候因子进行典范对应分析(CCA),阐明沙鞭种群种子的结实规律及驱动因素。此外,本文应用R语言3.3.4软件“Vegan”程序包和SPSS 24.0软件分别进行PCA聚类分析、典范对应分析(CCA)和Kruskal-Wallis检验,并利用R语言3.3.4软件“ggplot2”程序包制图。
2 结果与分析
2.1 沙鞭不同种群地理气候因子聚类
基于沙鞭天然种群经度、纬度、海拔和bio 1-bio 19等22個地理气候因子数据提取,通过22个地理气候因子主成分分析,采用特征根大于1作为主成分轴显著性对沙鞭137个种群进行欧式距离PCA聚类,前4个主成分占总变异的92.44%(表1),PC1主要包括经度等因子,PC2主要包括bio19等因子,PC3主要包括bio8等因子,PC4主要包括海拔等因子,结果表明沙鞭137个种群可被聚为3个组(表2)。即组1包括45个种群,其中种群1等代表种群主要分布在内蒙古高原西部;组2包括44个种群,其中种群4等代表种群主要分布在内蒙古高原中部;组3包括48个种群,其中种群30等代表种群主要分布在内蒙古高原东部(图2)。由此说明沙鞭不同种群间环境条件具有较大差异。
2.2 沙鞭不同种群组间结实情况差异
通过对沙鞭种群3个组内结实情况与各组频次(表2)统计加权,得到各组秩平均值和卡方值(表3),然后对3个组的结实情况进行Kruskal-Wallis检验。结果表明,沙鞭3个组间种子结实情况差异不显著(P=0.269),即沙鞭种群种子结实与其所处的地理气候因子没有直接相关性。
2.3 沙鞭种群种子结实情况与地理气候因子关系
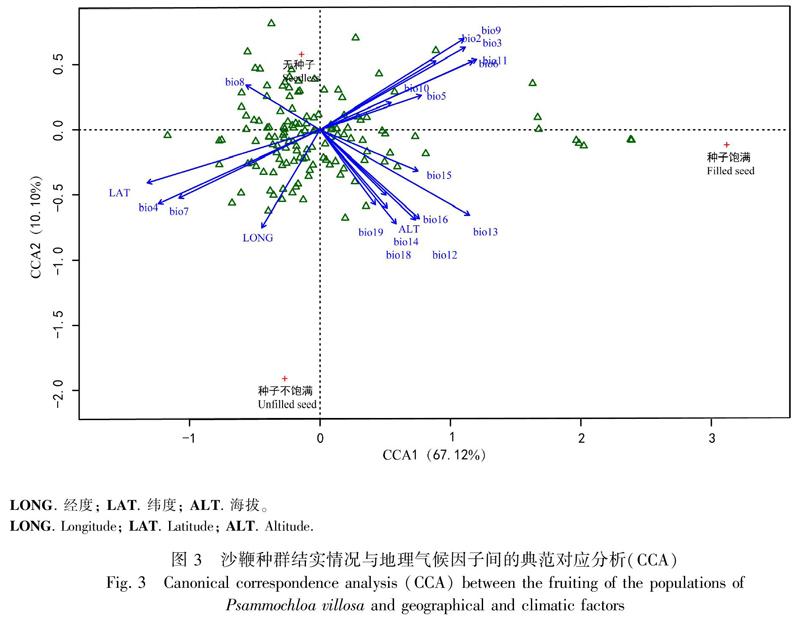
通过对沙鞭种群种子结实情况与地理气候因子间的典范对应分析(CCA),发现沙鞭种群种子结实情况与地理气候变量可得降维最小值2的维度,第一维度CCA1解释列联表的67.12%,第二维度CCA2解释列联表的10.10%,说明这两个维度上背景数据足以说明沙鞭种群种子结实情况的77.22%,CCA排序分析较为理想。沙鞭137个种群主要集中分布于典范轴中心,表明沙鞭不同种群间结实情况差异不明显(P>0.05);同时,与沙鞭种群无种子相比,种子饱满和种子不饱满距离中心相对较远,说明地理气候因子对沙鞭种群种子饱满度的影响远大于对无种子影响(图3)。此外,地理气候因子与种子饱满度具有显著相关性,其中,海拔、bio12-bio19与种子饱满度呈正相关,说明海拔、降雨等条件的改善有利于种子饱满度增加;经纬度、bio4和bio7与种子不饱满度夹角余弦值为正(图3),说明经纬度、 温度季节性变化和温度年变化是种子不饱满的主要驱动因素,而其他温度因子的改善则有利于种子饱满。
3 讨论与结论
本研究发现沙鞭种群间的结实情况差异与地理气候因子之间没有显著相关性(P > 0.05),且种群间的结实情况差异也不显著(P > 0.05),但地理气候因子是影响沙鞭种子结实的重要因素。前人研究表明,气候因子主要通过作用于植物生长过程中的物候期来影响植物结实(杨允菲等,1989;贺晓等,2005)。例如,杨允菲等(1989)研究发现干旱、高温、大风等逆境条件严重影响羊草受粉,致使其结实率急剧下降;贺晓等(2005)通过探讨诺丹冰草花序分化过程,发现低温会延长其幼穗分化期,从而使其分化出更多小穗,所以认为幼穗分化期是决定种子产量的关键时期。沙鞭与羊草和冰草均隶属于禾本科早熟禾亚科植物,且都以无性繁殖为主要繁殖方式,据此,我们推测气候因子可能同样通过影响沙鞭物候期影响其结实状况。沈奇等(2018)在研究环境因子与紫苏籽粒产量之间的关系时发现,较经度而言,纬度和海拔对植物生长影响更为显著,籽粒产量随着纬度的增高而增加,而随着海拔增加呈现先升高后降低的趋势。王仁忠(2000a,b)通过对羊草种群能量生殖分配规律的研究,认为茎和叶鞘的能量分配比例大于穗和叶的能量分配比例是羊草种群种子低产的重要原因,同时发现随着放牧强度增加,羊草的平均每穗结实数显著下降。因此,我们认为沙鞭种子结实不仅受到地理气候等非生物因子的影响,还受到种群能量生殖分配、放牧等生物因子的影响,是多种生态因子综合作用的结果。
同时,本研究发现海拔以及降雨条件的改善有利于增加沙鞭种子的饱满度,由此说明海拔与水分因子可能是制约沙生植物沙鞭有性繁殖的主导因素。沈奇等(2018)在对贵州省6个生态区域中紫苏籽粒产量与环境因子之间关系的研究中指出,海拔因子主要通过影响环境温度和降雨量来影响植物生长。随着海拔升高,温度降低,且降雨频度和总量较低海拔地区大(陈重潘,2010);同时,随着海拔升高,昼夜温差增大,植物光合色素含量和比例增高,有利于植物进行光合作用(祁建等,2007)。因而,海拔和降雨量与种子饱满度呈正相关关系,这与范丽颖等(2008)、刘莹等(2019)对花楸种子、砂生槐种子的研究结果相同。另外,种子饱满度与其发芽率具有显著正相关关系(陈惠哲等,2004;林惠玲,2018),我们推测海拔和水分因子是沙鞭有性繁殖成功的主导因素,而经纬度、温度季节性和年变化因子是沙鞭有性繁殖种群结实不饱满的促进因子。从繁殖成功角度分析,环境温度的波动不利于采取有性繁殖对策的种群,而可能会促进无性繁殖。例如,李春艳等(2015)发现小麦在灌浆期遇到高温胁迫时,将加快种子成熟速度、缩短灌浆时间,籽粒表现为瘦瘪,粒重明显下降;Li et al.(2014)发现夏季夜间增温可以促进羊草的无性繁殖,本研究结果与之一致。经纬度作为重要的地理因子,對土壤养分含量具有重要影响,王苗利等(2019)发现狗牙根土壤养分含量随经纬度改变呈现明显差异,经度与狗牙根土壤交换性钙含量及土壤pH呈显著负相关关系,而纬度与狗牙根土壤交换性钙和交换性镁含量呈显著正相关关系,据此我们推测沙鞭土壤养分含量与经纬度之间也可能存在一定的相关性。因此,研究沙生植物沙鞭种子的饱满度与环境因子之间的关系,对于提高沙鞭种子质量,筛选优良种质资源和遗传育种具有重要意义。
本研究证明沙鞭种群无种子特征仅与最湿季平均温度(bio8)存在正相关关系,不受诸多环境因子制约,据此我们推测最湿季平均温度是驱动沙鞭无性繁殖种群生长的主要因素,说明与有性繁殖相比,无性繁殖受环境因子制约少,适合度更高,同时解释了无性繁殖对策是沙生植物沙鞭种群环境适应性的重要保障,进一步验证了沙鞭种群主要依靠无性繁殖策略拓展种群数量特征的合理性。无性繁殖有利于提高个体的生存和竞争力,而有性繁殖对于提高和维持种群的进化适应能力十分必要(陈尚等,1999)。因此,研究不同生态环境条件下沙鞭种群繁殖对策有着重要意义,并将为探索其他同时具有克隆生长与有性繁殖的荒漠植物采取不同的繁殖对策机制提供理论参考。
参考文献:
CHEN CP,2010. Relationship between crop production and geographical latitude and altitude[J]. Gansu Agric,(3):27-28.[陈重潘,2010. 浅议地理纬度和海拔与作物生产的关系[J]. 甘肃农业,(3):27-28.]
CHEN HZ,ZHU DF,LIN XQ,et al.,2004. Effects of seed plumpness on germinating rate,seedling rate and growth of hybrid rice[J]. Fujian J Agric Sci,19(2):65-67.[陈惠哲,朱德峰,林贤青,等,2004. 杂交稻种子饱满度对发芽率、成苗率及秧苗生长的影响[J]. 福建农业学报,19(2): 65-67.]
CHEN S,MA Y,LI ZZ,et al.,1999. Analysis and measurement of the fitness of seed reproduction and vegetative reproduction of clonal plant[J]. Acta Ecol Sin,19(2):145-148.[陈尚,马艳,李自珍,等,1999. 克隆植物种子繁殖和营养繁殖的适合度分析和度量[J]. 生态学报,19(2):145-148.]
DONG M,A LA TB,XING XR,et al.,1999. Genet features and ramet population features in the rhizomatous grass species Psammochloa villosa[J]. Acta Phytoecol Sin,23(4):302-310.[董鸣,阿拉腾宝,邢雪荣,等,1999. 根茎禾草沙鞭的克隆基株及分株种群特征[J]. 植物生态学报,23(4):302-310.]
FAN LY,REN J,LIN YM,et al.,2008. Provenance variation of seed quality of Sorbus pohuashanensis in Northeast China[J]. J NE Norm Univ (Nat Sci Ed),40(3):131-135.[范丽颖,任军,林玉梅,等,2008. 东北地区花楸种子质量的种源变异研究[J]. 东北师大学报(自然科学版),40(3): 131-135.]
HE X,LI QF,LU HP,2005. Study on inflorescence differentiation of Agropyron desertorum cv. Nordan[J]. J Inn Mongol Norm Univ (Nat Sci Ed),34(1):106-109.[贺晓,李青丰,陆海平,2005. 诺丹冰草花序的分化过程研究[J]. 内蒙古师范大学学报(自然科学版),34(1):106-109.]
HUANG ZY,2003. Adaptation strategies of seed dormancy and germination of Psammochloa villosa,a sand dune grass inhabiting Ordos Plateau,China[J]. Acta Bot Boreal-Occident Sin,23(7):1128-1133.[黄振英,2003. 鄂尔多斯高原固沙禾草沙鞭种子休眠和萌发与环境的关系[J]. 西北植物学报,23(7):1128-1133.]
HUANG ZY,DONG M,ZHANG SM,2005. Strategies of seed germination on sand dune and seedling desiccation tolerance,of Psammochloa villosa (Poaceae)[J]. Acta Ecol Sin,25(2):298-303.[黄振英,董鸣,张淑敏,2005. 沙鞭(禾本科)种子在沙丘上的萌发策略及幼苗的耐干燥特性[J]. 生态学报,25(2):298-303.]
KUO PC,1987. Flora Reipublicae Popularis Sinicae[M]. Beijing:Science Press,9:307-309.[郭本兆,1987. 中国植物志[M]. 北京:科学出版社,9:307-309.]
LI A,GE S,2001. Genetic variation and clonal diversity of Psammochloa villosa (Poaceae)detected by ISSR markers[J]. Ann Bot,87:585-590.
LI CY,FU KY,ZHANG RQ,et al.,2015. Effect of high temperature post anthesis on the development of starch granules in winter wheat[J]. J Tritic Crops,35(10):1395-1402.[李春艷,付凯勇,张润琪,等,2015. 灌浆期高温对冬小麦淀粉粒发育的影响[J]. 麦类作物学报,35(10): 1395-1402.]
LIN HL,2018. Effects of seed plumpness and water content on rice germinating rate[J]. Seed Sci Technol,36(3):119-120.[林惠玲,2018. 种子饱满度及含水量对水稻发芽率的影响研究[J]. 种子科技,36(3):119-120.]
LIU HL,JIANG XY,LIU Q,2007. Theory of inclusive fitness and kin selection:kin altruistic behavior and its evolutionary mechanism[J]. Sci Technol Dialect,24(5):26-29.[刘鹤玲,蒋湘岳,刘奇,2007. 广义适合度与亲缘选择学说:亲缘利他行为及其进化机制[J]. 科学技术与辩证法,24(5): 26-29.]
L T,2019. Studies on genetic diversity of Psammochloa villosa,a peculiar sandy plant[D]. Xining:Qinghai Normal University:14-16.[吕婷,2019. 沙生牧草沙鞭的遗传多样性研究[D]. 西宁:青海师范大学:14-16.]
L T,LIU YP,ZHOU YH,et al.,2018a. Germplasm collection and preliminary studies on genealogical differentiation of a desert species Psammochloa villosa[J]. Acta Agr Sin,26(3):733-740.[吕婷,刘玉萍,周勇辉,等,2018a. 荒漠植物沙鞭(Psammochloa villosa)种质资源收集及谱系遗传分化初探[J]. 草地学报,26(3):733-740.]
L T,LIU YP,ZHOU YH,et al.,2018b. Micromorphological characteristics of leaf epidermis and ecological adaptation of Psammochloa villosa,a desert plant from the Inner Mongolian Plateau[J]. Bull Bot Res,38(3):330-337.[吕婷,刘玉萍,周勇辉,等,2018b. 沙生植物沙鞭叶表皮微形态特征及其生态适应性研究[J]. 植物研究,38(3): 330-337.]
LIU YX,1985. Flora in Desertis Reipublicae Populorum Sinarum[M]. Beijing:Science Press,1:34.[刘瑛心,1985. 中国沙漠植物志[M]. 北京:科学出版社,1:34.]
LIU Y,MA WJ,YANG GJ,et al.,2019. Geographical variation of the seed and germination characteristics of natural populations of Sophora moorcroftiana[J]. J NW For Univ,34(6):22-29.[刘莹,麻文俊,杨桂娟,等,2019. 砂生槐天然种群种实形态和萌发特征地理变异[J]. 西北林学院学报,34(6):22-29.]
LI ZL,LIN JX,ZHANG TY,et al.,2014. Effects of summer nocturnal warming on biomass production of Leymus chinensis in the songnen grassland of China:From bud bank and photosynthetic compensation[J]. J Agron Crop Sci,200:66-76.
MA YQ,1994. Flora Innermongolica[M]. Hohhot:Inner Mongolia People Press,5:115-152.[马毓泉,1994. 内蒙古植物志[M]. 呼和浩特:内蒙古人民出版社,5:115-152.]
QI J,MA KM,ZHANG YX,2007. The altitudinal variation of leaf traits of Quercus liaotungensis and associated environmental explanations[J]. Acta Ecol Sin,27(3):930-937.[祁建,马克明,张育新,2007. 辽东栎(Quercus liaotungensis)叶特性沿海拔梯度的变化及其环境解释[J]. 生态学报,27(3):930-937.]
SHEN Q,ZHAO JX,QIU XB,et al.,2018. Research on influence of environment factors to yield and quality traits of Perilla frutescent[J]. Chin J Chin Mat Med,43(20):4033-4043.[沈奇,赵继献,邱雪柏,等,2018. 环境因子对紫苏籽粒产量及品质性状影响研究[J]. 中国中药杂志,43(20): 4033-4043.]
WANG KQ,GE S,DONG M,1999. Allozyme variance and clonal diversity in the rhizomatous grass Psammochloa villosa (Gramineae)[J]. Acta Bot Sin,41(5):537-540.[王可青,葛颂,董鸣,1999. 根茎禾草沙鞭的等位酶变异及克隆多样性[J]. 植物学报,41(5):537-540.]
WANG ML,ZHANG JX,GUO ZP,et al.,2019. Effects of different longitudes and latitudes on soil nutrition in wild bermudagrass distribution areas in China[J]. Grassl Turf,39(5):53-61.[王苗利,张靖雪,郭志鹏,等,2019. 经纬度对中国野生狗牙根土壤养分的影响[J]. 草原与草坪,39(5):53-61.]
WANG RZ,2000a. Energy allocation to growth and reproduction in Leymus chinensis population[J]. Chin J Appl Ecol,11(4):591-594.[王仁忠,2000a. 羊草种群能量生殖分配的研究[J]. 应用生态学报,11(4):591-594.]
WANG RZ,2000b. Effect of grazing on reproduction in Leymus chinensis population[J]. Chin J Appl Ecol,11(3):399-402.[王仁忠,2000b. 放牧影响下羊草种群生殖生态学的研究[J]. 应用生态学報,11(3):399-402.]
YANG YF,ZHU YC,1989. A preliminary study on seed production of Aneurolepidium chinensis population[J]. Acta Phytoecol et Geobot Sin,13(1):73-78.[杨允菲,祝廷成,1989. 羊草种群种子生产的初步研究[J]. 植物生态学与地植物学学报,13(1):73-78.]
(责任编辑 周翠鸣)
