沙柳SpsNAC042基因克隆及逆境表达分析
2021-10-14杨海峰薄高峰于兴旺李安玉
杨海峰,薄高峰,于兴旺,李安玉,张 鑫,郝 璞,刘 璐
(内蒙古农业大学 林学院,内蒙古 呼和浩特 010018)
NAC家族转录因子是一类植物所特有的转录因子,其有多种生物功能且广泛分布于陆生植物中。在NAC蛋白质N端的结构域是高度保守和特异的[1]。NAC转录因子家族成员众多,主要涉及参与植物的生长发育调控、植物的逆境胁迫应答、调控植物的抗病性、植物次生生长调控以及激素信号转导等过程。在植物不同发育时期和多种环境因素诱导下,激活特定目的蛋白,发挥着各种重要的生物学功能[2]。
NAC家族转录因子被认为是至今发现的植物基因组中最大的转录因子家族之一[3]。E.Soueretal[4]在矮牵牛(Petuniahybrida)中克隆出了NAC转录因子基因NAM(no apical meristem)。随后在矮牵牛和拟南芥(Arabidopsisthaliana)中发现了ATAF1/2和CUC2这2个转录因子。并以其首字母命名为NAC结构域[5]。NAC基因目前已在拟南芥、大豆(Glycinemax)、玉米(Zeamays)、棉花(Gossypiumspp.)、沙冬青(Ammopiptanthusmongolicus)、苹果(Malusdomestica)、小黑杨(Populus×xiaohei)、柽柳(Tamarixchinensis)等多种植物中获得,但木本植物中研究并不深入。其中以杨树为研究对象,对NAC家族进行了系统分析,共鉴定出163个NAC基因家族成员,进化分析显示这些基因被聚类成18个亚家族[6]。通过从小黑杨中克隆获得的NAC7基因并分析得出NAC7基因对高盐胁迫具有应答反应,基因主要在根中具有盐胁迫应答效应[7]。苏柳(Salix×jiangsuensis‘J2345’)中也成功克隆NAC基因,并推测柳树SlNAC1基因在非生物胁迫下表达较为稳定,受ABA和GA胁迫诱导[8]。
沙柳(Salixpsammophila)是中旱生落叶灌木,内蒙古西北地区主要防风固沙树种。具有根系发达、耐干旱、易成活、生长快等特点,适宜在沙区生长,有很好的固碳释氧和防风固沙作用[9]。目前已经从分枝角度[10]、生理胁迫[11]等多个方向展开研究。本研究由沙柳中克隆获得SpsNAC042基因,进行了生物信息学分析,对SpsNAC042基因开展了组织特异性表达及逆境下的表达分析。本研究为沙柳NAC基因家族的挖掘及逆境分子调控机制研究提供了基础,为相关调控机制研究提供重要理论依据,为进一步获得优异抗逆种质资源和种质创新奠定了理论基础。
1 材料与方法
1.1 试验材料
供试材料来自内蒙古达拉特旗国家沙柳无性系种质资源保存库,选取长势良好的1年生单一无性系沙柳枝条(无性系编号11~30)扦插至内蒙古农业大学苗圃地。待成活后截取长势良好、长约50 cm的幼苗,放入植物培养箱中水培,待长出10 cm嫩枝后提取RNA用于基因克隆。
选取沙柳(无性系11~30)生长良好的茎尖、茎、叶、花、幼叶,置于液氮中保存待用。将枝条水培至生根约5 cm,取根置于液氮中保存待用。取沙柳嫩枝放置在2 L三角瓶中。以20%聚乙二醇(PEG)6000干旱胁迫处理;250 mmol NaCl盐胁迫处理;培养箱中进行4℃水培冷处理;培养箱进行40℃水培热处理;培养箱进行24℃水培对照组。溶液保持在1/2,光照14 h黑暗10 h,并在1、2、4、8、12 h和24 h处理后收集叶片,3个生物学重复,每个重复3株,置于液氮中保存待用。
1.2 试验方法
1.2.1 RNA提取及cDNA反转录 使用天根RNAprep Pure多糖多酚植物总RNA 提取试剂盒(DP441)提取总RNA,以1%琼脂糖凝胶电泳检测RNA的完整性。取4 μg总RNA,以Trans Script All in one First strand cDNA Synthesis Supermix for qPCR试剂盒进行反转录合成cDNA,用于克隆基因。
1.2.2 基因克隆 以全株的cDNA为模板,NAC1-F、NAC2-R为引物,利用A8聚合酶(北京艾德莱2×A8 Fast HiFi PCR MasterMix)进行PCR反应,反应条件为95℃预变性5 min;98℃ 10 s,60℃ 30 s,72℃ 1 min,35个循环;72℃ 5 min,4℃保存。以1%琼脂糖凝胶电泳检测PCR产物,切胶回收后连接克隆载体pEASY-Blunt zero CloningVector(全式金,北京)。选取阳性克隆菌斑,送至北京Thermo Fisher公司进行测序。
1.2.3SpsNAC042基因生物信息学分析 通过 NCBI 的 ORF(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)在线软件分析NAC基因的开放阅读框;利用Phytozome 11数据库搜索同源核苷酸及氨基酸序列;ExPASy(http://web.expasy.org/protscale/)在线预测亲水性和疏水性;ExPASy-ProtParam tool(http://web.expasy.org/protparam/)预测等电点、分子量;在线软件NPS@(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_gor4.html)预测二级结构。Singal P4.1预测信号肽,http://www.cbs.dtu.dk/services/TMHMM/预测跨膜结构;T-COFFEE (http://tcoffee.crg.cat/apps/tcoffee/index.html)进行氨基酸多序列比对;MEME Suite(http://meme-suite.org/)进行氨基酸结构比对;利用软件MEGA6构建系统进化树[12]。
1.2.4 沙柳SpsNAC042基因RT-qPCR表达分析 利用Primer3根据克隆的CDS序列设计定量引物qN1-F(GCAACTGGACTAAGAACGAATC G),qN1-R(ACGACCCAGTACTGGCCCTT),保证其特异性。利用实时定量PCR(RT-qPCR)的方法,以UBQ为内参基因,分析NAC042基因在沙柳不同组织中的表达量。用Thermo RNA试剂盒分别提取正常条件下生长沙柳的根、茎、叶、花、茎尖,幼叶等组织的RNA;提取不同时间点经过干旱胁迫、盐胁迫、低温胁迫、高温胁迫成熟叶片的RNA。将提取的RNA进行cDNA第一链合成。采用 2-ΔΔCt法分析RT-qPCR试验数据。
2 结果与分析
2.1 SpsNAC042基因克隆
提取沙柳全株的总RNA,经琼脂糖凝胶电泳与紫外分光光度计分析,提取的RNA质量较好,可用于cDNA转录。以NAC1-F、NAC1-R为引物,进行PCR扩增,扩增片段与目标条带900 bp一致(图1)。经Thermo Fishe公司测序,确定克隆获得900 bp的目标基因CDS序列(图2)。

图2 SpsNAC042基因CDS及蛋白序列Fig.2 SpsNAC042 gene CDS and protein sequence
2.2 目标序列的生物信息学分析
2.2.1 蛋白理化特性分析 扩增得到完整的CDS序列,长900 bp,编码299个氨基酸。利用ExPASy-ProtPaRamtool软件在线分析SpsNAC042氨基酸序列的理化性质。测定蛋白亲水性,SpsNAC042理论等电点(PI)为6.45,分子量为34 190.99 ku,SpsNAC042基因蛋白质分子式为C1521H2344N406O452S20,蛋白质的半衰期为30 h,不稳定指数51.39,为不稳定蛋白。
2.2.2 SpsNAC042蛋白亲水性和跨膜区分析 利用ExPASy-ProtPaRamtool分析得到SpsNAC042蛋白在第42个氨基酸处,疏水性最强,最大值1.578,在第200个氨基酸处,亲水性最强,最小值-2.444,总平均疏水指数(GRAVY)-0.460,蛋白质亲水区域多于疏水区域,是亲水性蛋白(图3)。带负电荷的残基总数(Asp+Glu):36,带正电荷的残基总数(Arg+Lys):35。利用Pfam在线软件分析,SpsNAC042只有1个蛋白结构域NAM,氨基酸序列12~134为NAM结构域。
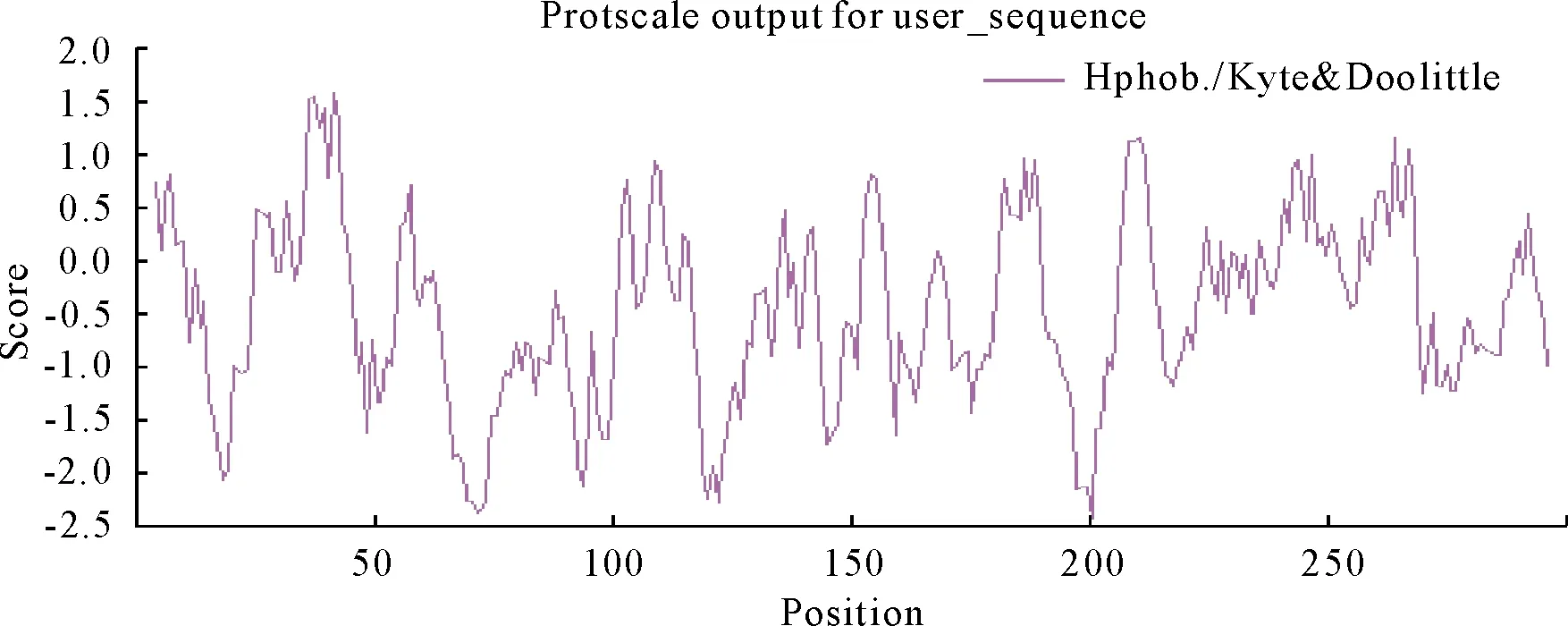
图3 蛋白亲疏水性分析Fig.3 Distributing curve of hydropathy of the protein
2.2.3 SpsNAC042蛋白质结构预测 蛋白质结构显示,SpsNAC042主要包含结构形式为α螺旋(alphahelix)占9.70%,延伸链(extendedstrand)占24.41%,无规则卷曲(randomcoil)占65.89%。3种结构所占比列差异比较大,以无规则卷曲为主,α螺旋最小(图4)。SpsNAC042基因的N端和C端以延伸链和无规则卷曲为主。三级结构中全局模型质量评估(GMQE)为0.36,覆盖率48.72%(图5)。通过SWISS-MODEL对SpsNAC042蛋白质的三级结构预测,得到的三级结构(图5)。
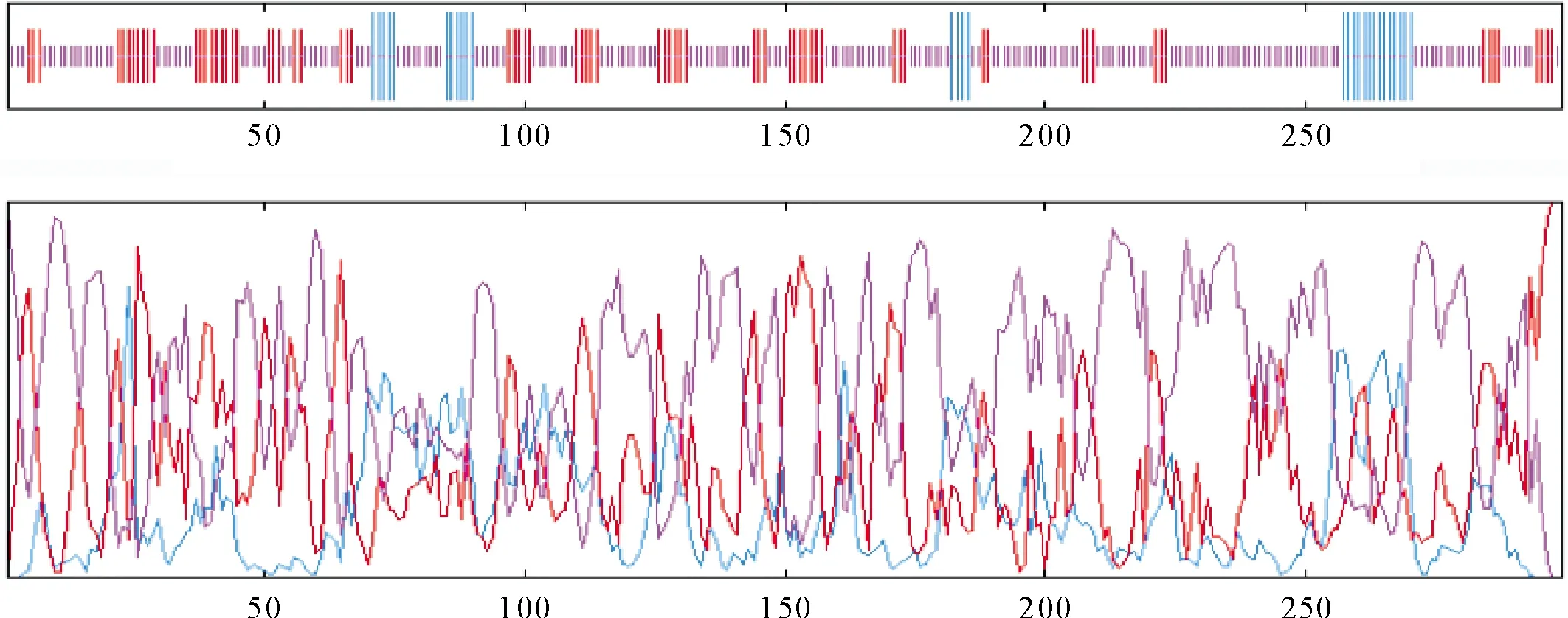
图4 SpsNAC042蛋白二级结构预测Fig.4 Prediction of secondary structure of SpsNAC042 protein
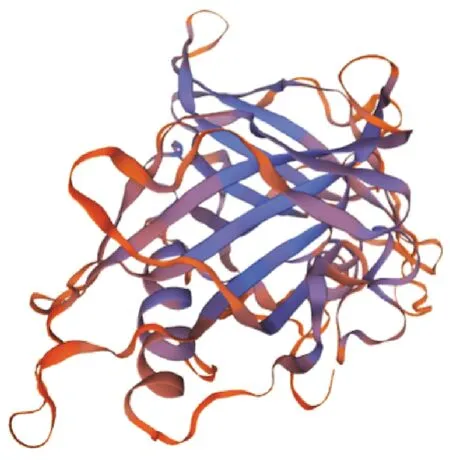
图5 SpsNAC042蛋白的三级结构预测Fig.5 Putative of SpsNAC042 protein tertiary structure
2.2.4 SpsNAC042蛋白同源序列分析及同源结构域分析 将克隆获得的SpsNAC042序列与欧洲红皮柳SapurV1A.0284s0040.1、毛果杨Potri.007G065400.1、苹果MDP0000298182、桃子Prupe.4G143600.1、拟南芥AT1G56010.2、水稻LOC_Os06g46270.1、玉米GRMZM2G063522_T01所编码的氨基酸序列进行多序列比对分析(图6)。SpsNAC005与其他的氨基酸序列都具有1个典型的保守结构域NAM,且均有A、B、C、D、E共5个氨基酸同源序列区域。同时N 端序列相对保守,而 C 端是高度多变的转录激活功能域,符合NAC基因家族的典型特征。上述结构表明克隆所获得的基因属于NAC基因家族成员。
利用MEME Suite在线软件,对SpsNAC042和同源基因进行同源结构域对比(图6),结构域参数设定为12。结果表明SpsNAC042和对比植物均有保守结构NAM,均具有A、B、C、D、E共5个保守亚结构域,符合NAC基因家族的特征。其中柳科与杨树科亲缘关系相近,所分析的结构域完全一致。这组对比基因的同源结构相似度很高,推测SpsNAC042在进化过程中,结构较稳定。
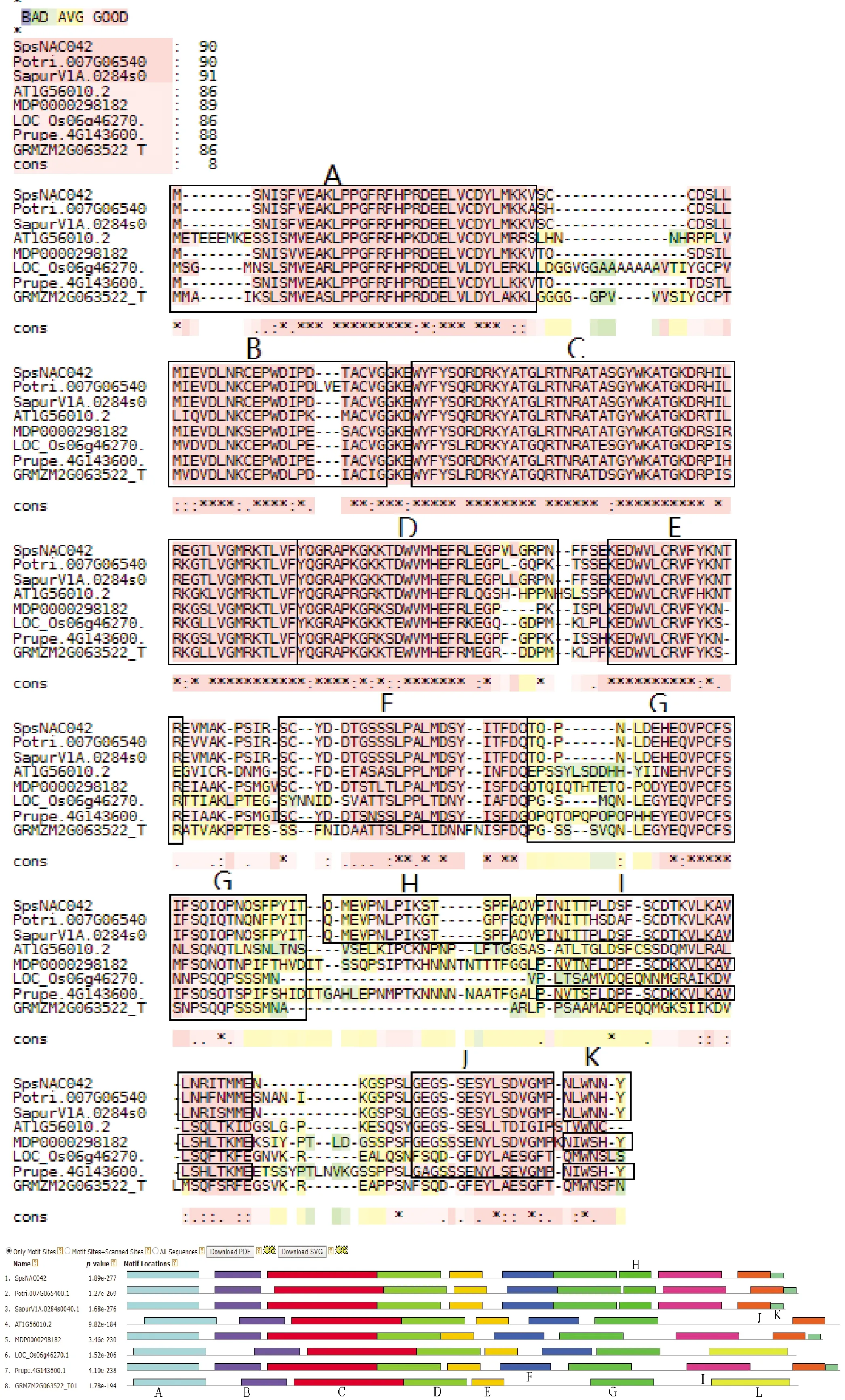
图6 SpsNAC042基因家族氨基酸序列多重比对和同源结构域对比Fig.6 Multiple alignment of amino acid sequences and comparison of homologous domains of SpsNAC042 gene family
2.2.5SpsNAC042系统发育树构建 将本研究克隆所得的SpsNAC042基因编码的氨基酸序列与拟南芥、欧洲红皮柳、毛果杨、717杨、苹果、桃子、拟南芥、水稻、玉米同源基因,利用NJ算法进行进化树分析(图7)。结果表明,沙柳NAC042与欧洲红皮柳、毛果杨及毛白杨科属亲缘关系相近的为同一进化分支,木本植物与草本植物在不同进化分支,预测NAC基因有可能在木本植物和草本植物中存在分化,与上述同源序列和同源结构域分析结果一致。
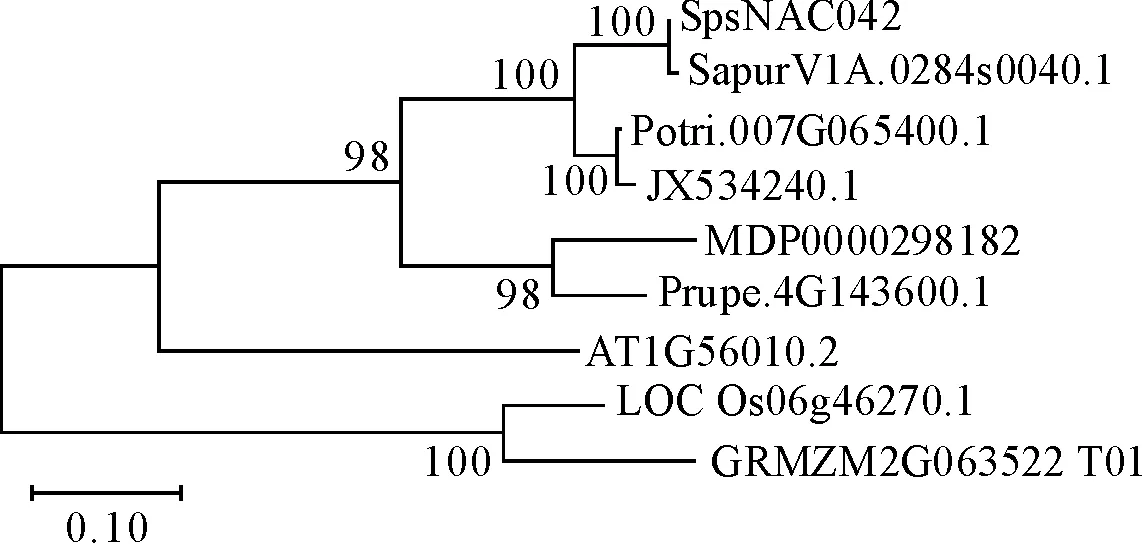
注:SapurV1A.0284s0040.1欧洲红皮柳、Potri.007G065400.1毛果杨、JX534240.1 717杨、MDP0000298182苹果、Prupe.4G143600.1桃子、AT1G56010.2拟南芥、LOC_Os06g46270.1水稻、GRMZM2G063522_T01玉米图7 SpsNAC042基因家族系统进化树Fig.7 The neighbor-joinning tree of SpsNAC042 gene family
2.2.6 沙柳SpsNAC042基因的组织特异性定量分析 以沙柳7个不同组织部位为模板,通过中RT-qPCR检测分析各组织部位的相对表达量(图8)。以UBQ基因为参照,其中花中的基因相对表达量最高,幼叶、茎9~15节间根、叶的表达量依次降低,茎尖的表达量最低。
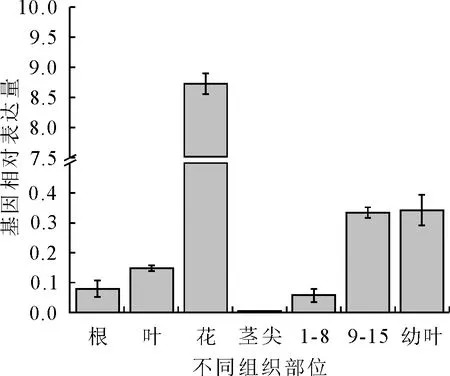
图8 RT-qPCR分析SpsNAC042基因的组织特异性表达Fig.8 Tissue specific expression analysis of SpsNAC042 with RT-qPCR
2.2.7 沙柳SpsNAC042基因在逆境处理下定量表达分析 将沙柳在干旱胁迫、盐胁迫、低温胁迫、高温胁迫进行处理,利用RT-qPCR分析SpsNAC042基因的相对表达量。结果表明,在干旱处理下SpsNAC042基因与对照组相比整体呈增长趋势,并在8 h处达到最高;在低温处理下,基因表达量迅速积累,在2 h处达到最高,然后维持在较低水平表达;(图9A、9C)。在盐和高温处理下SpsNAC042基因缓慢积累,后期迅速增高,最终在24小时处达到最高(图9B、9D)。
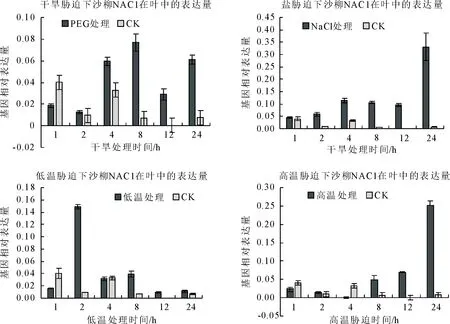
图9 RT-qPCR分析SpsNAC042基因在不同逆境下的特异性表达Fig.9 Specific expression of SpsNAC042 gene under different stresses of RT-qPCR analysis
3 结论与讨论
多序列及结构域比对分析表明,SpsNAC042蛋白含有 1 个高度保守的典型NAM结构域。系统发育树构建表明SpsNAC042基因与欧洲红皮柳和毛果杨同源基因的同源性较高。SpsNAC042基因在盐胁迫和高温胁迫下应答较为明显。在低温和干旱胁迫下有应答。该基因归属于SNAC(Stress-responsiveNAC)大类。
本研究以木本植物沙柳为材料,选取SpsNAC042转录因子基因进行抗逆功能分析。M.Nuruzzamanetal[13]研究表明,20%~25%的NAC基因都至少与一种胁迫相关[13]。而干旱、高盐、低温、高温是影响植物生长发育的主要逆境因子[14]。故本研究应用荧光定量PCR法分析了SpsNAC042基因在干旱、高盐、冷和高温胁迫下的表达模式,发现NAC基因与多种非生物胁迫响应相关。在有关香蕉(Musanana)的研究中表明,MusaNAC042在提高转基因香蕉植株的耐盐性和耐旱性方面的作用明显,MusaNAC042的表达与盐胁迫和干旱胁迫呈正相关[15]。在小黑杨的研究中表明NAC7转录因子基因对盐胁迫具有应答反应[7]。上述研究与本研究结果相似,在本研究中干旱、低温、高温、盐胁迫均能诱导SpsNAC042基因的表达,但在应答快慢上有不同程度的差异。该结果表明沙柳NAC042基因对胁迫有所应答,很有可能参与沙柳的抗逆调控。
根据同源序列分析,本研究由沙柳中克隆获得SpsNAC042基因,进化树分析表明与草本植物拟南芥NAC1(AT1G56010)基因为同源基因[16]。在拟南芥中研究发现NAC1转录因子可能参与外植体特异性伤口信号传导途径。在器官形成、再生、侧根、不定根形成中发挥不同的作用[16]。有学者研究表明,与拟南芥NAC1为同源基因的葡萄VvNAC1基因在非生物胁迫下有积极作用,VvNAC1基因在发育中的花表达增强[17]。在本研究基因组织特异性表达与之相吻合。此外,研究人员还进行了一系列的生物胁迫试验,并表示VvNAC1基因可能会触发不同信号防御途径的调节[17]。因为NAC基因结构域的C端是高度多变的转录激活功能域,可能导致不同调控功能。后续研究会补充该试验内容。
根据基于拟南芥、水稻、石松和苔藓中大部分NAC基因的系统进化树分析研究,将功能相近的NAC基因各自归类,其中胁迫响应的NAC基因都归属于SNAC(Sress-responsiveNAC)大类[18]。NAC转录因子中很多成员对非生物胁迫都具有应答反应,如柑橘在低温、干旱、高盐和ABA胁迫条件下NAC83基因呈现出差异性的表达[19]。MdNAC1转基因苹果植株在干旱胁迫条件下,抗旱性明显增强[20]。玉米ZmNAC62转基因株系参与调控干旱,低温胁迫[21]。AgNAC1对高温,低温,干旱和盐胁迫均有响应[22]。根据上述研究及本研究中SpsNAC042基因在逆境下的相应,分析预测SpsNAC042基因应该在沙柳遭遇生物或非生物胁迫时响应表达,为正调节因子,隶属SNAC大类。
