药用植物连作障碍研究进展
2021-09-11胡双孙文静高林怡郭龙妹王莉莉黎万奎
胡双 孙文静 高林怡 郭龙妹 王莉莉 黎万奎


摘要:中医药作为中华民族的瑰宝,在防病治病方面发挥着巨大作用。为了满足国内外市场需求,药用植物的栽培面积不断扩大,随之而来的连作障碍问题也愈发突出,严重制约了中医药产业的持续健康发展。本文根据近年来药用植物连作障碍方面的研究成果,阐述了其危害,从土壤、植物、微生物3个方面分析其产生的原因,并总结了目前常用的防控措施,主要有合理施肥、改良耕作制度、品种选育和土壤灭菌,同时提出存在的一些问题,以期为今后的相关研究提供参考。
关键词:药用植物;连作障碍;化感物质;土壤酶;根际;防控措施;微生物
中图分类号: S567.04 文献标志码: A 文章编号:1002-1302(2021)16-0038-10
连作障碍(continuous cropping obstacles)指在正常的管理措施下,在同一块地连续多年种植相同作物而造成的作物产量降低、品质变劣、生长状况变差、病虫害加剧的现象[1]。欧美国家称之为“再植病害”或“再植问题”,日本称之为“忌地现象”或“连作障害”,在我国常称为“重茬问题”。连作障碍在农业生产中较为普遍,每年给全世界带来巨大损失。我国是中药消费大国,中药的进出口量逐年增加,种植面积不断扩大。目前,我国常用中药材有600多种,其中已实现人工种植的有300多种,种植面积达220多万hm2[2]。药用植物相对于其他作物而言有特殊的化学成分和现实应用,且在生产过程中更易发生连作障碍,因此克服药用植物连作障碍问题十分必要。本文根据近年来国内外药用植物连作障碍方面的文献,对其危害、产生原因和防控措施进行归纳总结。
1 药用植物连作障碍的危害
1.1 药材产量质量下降
大量研究发现,药用植物连作后,受各种因素影响,环境逐渐不适宜植物生长,导致其生长发育受阻,产量不断下降,严重的甚至出现绝收。例如,温郁金(Curcuma wenyujin Y. H. Chen et C. Ling)連作会造成地下块根和地上茎叶的生物产量远小于正茬,尤其是生长中期对地上部分的茎叶生物产量影响最大,蓬莪术(Curcuma phaeocaulis Val.)连作也会造成地上茎叶生物产量远低于正茬[3]。蒙古黄芪[Astragalus membranaceus var. mongholicus (Bunge) P. K. Hsiao]连作5年后植株生长缓慢,各项生物量指标较低,药材品质下降[4]。地黄[Rehmannia glutinosa (Gaertn.) DC.]连作后,其叶绿素含量、光合特性及根系活力在整个生育周期中均维持在较低水平,产量显著降低[5]。
此外,药用植物连作还会导致其次生代谢产物的合成紊乱,出现药材有效成分含量减少、质量下降、临床疗效变差的情况,这是区别于其他作物连作障碍最突出的特征。白花丹参(Salvia miltiorrhiza f. alba C. Y. Wu et H. W. Li)连作2年后,其地下部鲜质量和干质量大幅下降,且有效成分二氢丹参酮、隐丹参酮、丹参新酮、丹参酮ⅡA、丹酚酸B和迷迭香酸均不同程度下降[6]。白术(Atractylodes macrocephala Koidz.)连作后其根长、直径、体积、单株鲜质量、干质量及产量均降低,白术内酯Ⅰ、Ⅱ、Ⅲ降低,且连作1、2年白术的水分含量和连作3年白术的灰分含量均不符合2015年版《中华人民共和国药典》标准[7]。穿龙薯蓣(Dioscorea nipponica Makino)中皂苷类活性成分的含量随连作年限的增加而逐渐下降[8],广藿香[Pogostemon cablin (Blanco) Benth.]连作后,其挥发油、百秋李醇含量也明显下降[9]。
1.2 自然生态坏境破坏
由于药用植物连作障碍的发生损害了农民的经济效益,人们为了弥补损失,盲目地更换或扩张种植面积,破坏了原有的自然环境和植被类型,造成当地物种单一、生物多样性降低、生态环境脆弱、水土流失严重。此外,有些药农在连作障碍出现后,盲目增施化肥农药,不仅造成土壤质量下降、农药残留严重,而且往往收效甚微,增加了成本和浪费资源。在药用植物的异地迁移种植中,有的忽略了自然生产规律,实际效果不佳,同样造成了对自然资源的浪费和生态环境的破坏。
2 药用植物连作障碍的产生原因
药用植物连作障碍的产生过程十分复杂,是多种因素共同作用的结果,国内外学者从多角度提出了不同的解释,总的来说可以概括为土壤、植物和微生物3个方面。
2.1 土壤因素
2.1.1 土壤物理性状的改变 药用植物连作后常发生土壤物理性状的改变,长期连作会打破土壤的动态稳定,使土壤板结僵化、透气性差,CO2和有害气体则不断积累在根系周围,当达到一定浓度时,就会引起根系中毒[10]。当土壤中的O2、CO2浓度及孔隙率等无法满足根系正常生长的需求时,则会阻碍植物对养分的吸收利用,最终严重制约药用植物的生长发育。简在友等发现,随连作年限的增加,种植人参(Panax ginseng C. A. Mey)的土壤中的物理性黏粒(粒径<0.01 mm)逐渐增多,团粒结构被破坏,发生土壤板结,土壤的非活性孔隙比例相对降低,通气、透水性能变差,进而影响人参药材的品质[11]。祖勒胡玛尔·乌斯满江发现,甘草(Glycyrrhiza uralensis Fisch)连作地的土壤容重显著大于轮作地,土壤养分、总孔隙度显著小于轮作地[12]。
2.1.2 土壤化学性状的改变 土壤酸化是指氢离子在土壤内部产生并从土壤外部输入,导致土壤pH值降低和盐基饱和度降低的过程,它将导致土壤中重金属元素的溶解和释放,并减少氮、磷、钾、有效硼和有效硅等营养元素含量,不利于药用植物的生长发育。在自然条件下,土壤酸化过程相对较慢,平均每过229万年土壤的pH值下降1个单位,但受人为活动影响,土壤酸化加速,有研究发现,我国农田土壤pH值在1980—2000年平均下降了0.5个单位[13]。目前,已经在多种药用植物如人参、孩儿参[Pseudostellaria heterophylla (Miq.)Pax ex Pax et Hoffm.]、西洋参(Panax quinquefolius L.)、丹参(Salvia miltiorrhiza Bunge)、白术、三七[Panax notoginseng (Burk.) F. H. Chen]、地黄、白花蛇舌草(Hedyotis diffusa Willd.)、大蒜(Allium sativum L.)、乌头(Aconitum carmichaelii Debx.)等中出现该问题,主要是长期连作导致土壤中酚酸类物质积累造成的。
土壤盐渍化是一个日益严峻的全球性问题,对植物生长发育的影响显著,主要是由于化肥的滥用造成土壤盐分积累过多。盐对植物的毒害作用一方面是由于土壤中钠的积累导致水的有效利用减少,另一方面是由于钠、氯离子对植物的毒性作用[14]。此外,盐胁迫还会造成药用植物光合作用下降[15],如当归[Angelica sinensis (Oliv.) Diels]连作叶片的光合特性、光合色素含量及药材挥发油含量显著降低,造成当归的产量、质量下降[16]。丙二醛(MDA)作为植物在盐胁迫下膜脂过氧化的主要产物,其含量可代表细胞膜的损伤程度,反映了植物的盐胁迫和耐盐性[14]。脯氨酸(Pro)是一种重要的渗透调节物质,其含量可作为植物抗逆性的生理指标[17]。兰州百合[Lilium davidii var. willmottiae (E. H. Wilson) Raffill]連作后,其叶片光合色素含量显著降低,光合荧光参数劣化,叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性下降,渗透调节物质MDA、Pro含量增加,可溶性蛋白含量降低[18];半夏[Pinellia ternata (Thunb.) Breit.]连作后也出现了类似的情况,但可溶性蛋白、可溶性糖含量升高[19]。以上研究结果均表明,连作后出现的土壤盐渍化是造成连作障碍的重要因素之一。
2.1.3 土壤养分失衡 由于药用植物在生长过程中对土壤中营养元素的需求有一定规律,并且在同类或近缘植物根系的分布情况相似,其根系大致停留在同一土层深度,长期连作必然造成土壤中某些元素的亏损或富集,从而造成土壤养分的偏耗,使土壤营养元素失衡,进而影响下茬植株的正常生长[20]。此外,由于土壤中的氮、磷、钾等大量元素更易受到人们关注,能够及时通过施肥给予补充,因此药用植物在生长过程中对微量元素的竞争十分激烈,而且可能受到外界环境如温度、湿度等的诱导,使其在特定时期对某种元素的吸收增多,长期恶性循环后形成“木桶效应”,对药用植物的生长十分不利。高桐梅等发现,随着芝麻(Sesamum indicum L.)连作年限的增加,土壤速效氮、磷、钾含量和交换性钙、镁含量及有效锰、硼、铁、铜、锌含量均下降,土壤供肥能力变差[21]。吴凤云发现,三七连作后土壤pH值、硝态氮、铵态氮、速效磷和全钾含量下降,而土壤Ca2+、Mg2+、HCO-3和全磷含量升高[22]。刘姣姣等发现,随着花椒(Zanthoxylum bungeanum Maxim)连作年限的增加,土壤有机质、全氮、全磷、全钾含量和速效氮、速效磷、速效钾等含量均有所降低,土壤pH值为8.09~8.48,呈弱碱性[23]。
2.1.4 土壤酶活性降低 土壤酶作为土壤系统的重要组成部分,是土壤中动植物残体分解、植物根系分泌和土壤微生物代谢的产物,是一类具有催化活性的特殊物质,参与了土壤中有机质的分解和腐殖质的形成等生化过程,是土壤质量和土壤生物活性的综合表现[24]。土壤酶活性受土壤性质和环境条件的影响,可以代表土壤中物质代谢的旺盛程度,并在一定程度上反映植物对养分的吸收利用情况[25-26]。如CAT能够酶促分解对植株、土壤产生毒害作用的过氧化氢;蔗糖酶(SUC)可将土壤中的蔗糖转化为可被植物利用的葡萄糖和果糖;脲酶(URE)是土壤中对尿素转化起重要作用的酶类,可表征土壤的供氮能力;磷酸酶(PHO)能酶促分解各种有机磷,为植物生长提供有效磷素;蛋白酶(PRO)参与土壤中蛋白质、氨基酸及其他含蛋白质氮的有机化合物的转化;多酚氧化酶(PPO)可以驱动土壤中芳香族化合物的分解与转化,从而消除土壤中芳香族污染物对土壤质量的不良影响[27-29]。表1为近年来关于药用植物连作对土壤酶活性影响的报道,可以看出,连作后多数土壤酶活性降低,不利于土壤的健康稳定和药用植物的生长发育,而PPO活性上升可能受到土壤干湿状况、溶解氧含量、pH值和温度等因素影响[38],会阻碍有机质矿化时产生的酚类中间产物合成腐殖质,造成酚类化合物的积累[39],从而引起植物中毒。
2.2 植物因素
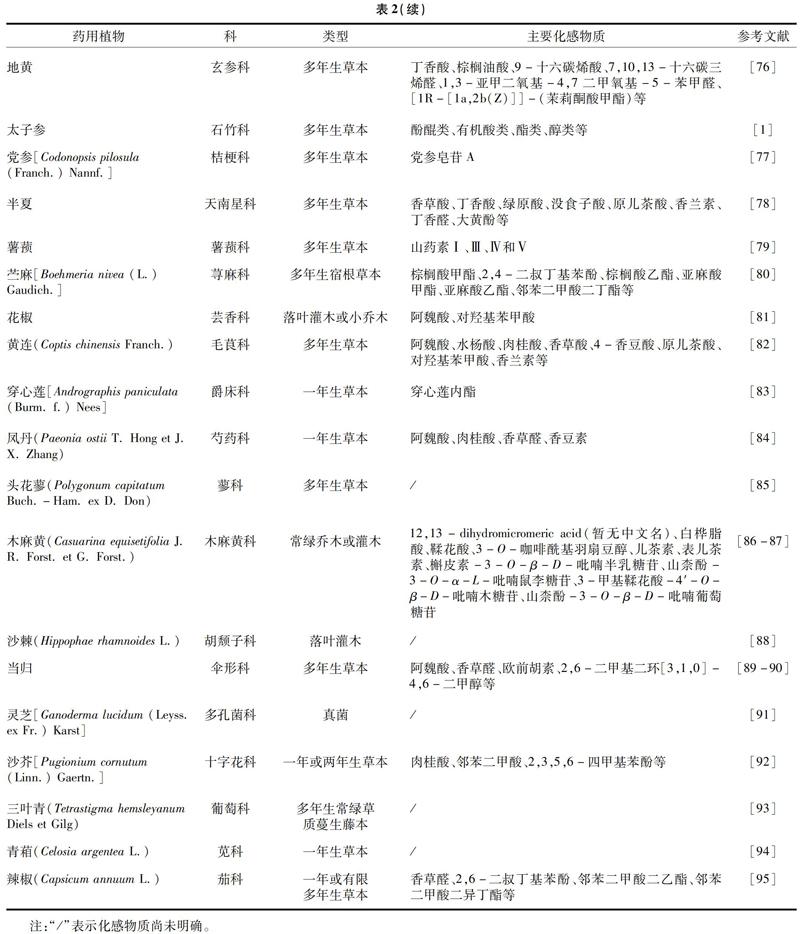
药用植物连作后,常常会出现化感自毒现象。早在1938年,Mahboobi等便首次提出化感作用(selfalleloppathy)这一概念[40-42],即植物通过淋溶、挥发、腐解及其根系分泌物等特殊途径释放一些物质对植物、微生物的生长产生促进或抑制的现象。化感作用普遍存在于自然界中,主要包括化感偏害作用、自毒作用、自促作用和互惠作用[43]。自毒作用即一种发生在种内的生长抑制作用,植物连作后,前茬植物残体与病原微生物的代谢产物可对植物产生毒害,连同植物分泌的自毒物质共同干扰植株生长代谢,最终形成了植物的化感自毒现象[44]。化感物质种类多样,存在于药用植物的各个组织中,引起药用植物自毒的机制主要是破坏细胞结构、影响光合作用、影响活性氧的代谢和干扰激素代谢平衡等[45]。表2为近年来关于药用植物化感自毒方面的报道,相对来说,它们较其他药用植物不耐连作,更易受到化感物质的侵害。从科级上看,药用植物主要集中在菊科、百合科、唇形科、豆科和五加科;从植物类型上看,这些植物大多为草本植物,且以多年生草本为主;从化感物质上看,主要是酚酸类物质,还包括药用植物的一些特有成分。从文献报道的频率上来看,主要集中在一些市场需求大和栽培面积广的药用植物,如白术、大蒜、丹参、人参、西洋参、三七、地黄和太子参等,它们的连作障碍问题也更加严重。但不表明未被报道的药用植物就没有化感自毒的情况,可能是因为其野生资源已满足市场需求而不需要大规模的人工栽培或是因为问题不突出而并没给人们带来实质性的损失,所以未得到关注。
2.3 微生物因素
2.3.1 微生物种类和数量降低 土壤微生物是陆地生态系统的重要组成部分,主要包含细菌、放线菌和真菌三大类群,其数量不仅可以反映能量代谢的过程,还可反映土壤肥力状况,在土壤养分循环和土壤矿物分解中起着重要作用,土壤微生物可以产生和利用各种大气成分(如N2、O2、H2、NO、N2O、CO2、CH4、H2S等),其种类和数量直接影响土壤的生化活性和养分转化,并调节土壤养分动态[96-99]。一般情况下,土壤中细菌最多,放线菌和真菌较少[10],而连作会显著改变药用植物根际微生物群落结构,造成微生物种类和数量降低,且根际土壤中真菌数量增多、细菌数量减少,土壤微生物类型由细菌型向真菌型过渡。众多学者一致认为,真菌型土壤是地力衰退的标志[100]。肖春萍等研究发现,人参栽培年限增加会显著抑制人参根际土壤微生物活性及微生物量的积累[101]。张周等发现,玉竹连作后,根际土壤中细菌、放线菌数量分别较对照土壤降低了12.4%、13.2%,真菌数量则增加了239%,好氧自生固氮菌、氨化细菌、无机磷细菌数量分别降低了36.3%、48.6%、71.1%,而有机磷细菌数量增加了12.1%[102]。以上研究结果均表明,连作不利于土壤微生物的稳态,从而导致药用植物产生连作障碍。
2.3.2 致病菌增多和益生菌减少 大量研究结果表明,土壤微生物群落在土壤生态系统可持续性中发挥着重要作用[103-104]。连作后土壤环境发生改变,逐渐不适宜多数微生物的生存,土壤微生物的种类、数量都下降,此时环境有利于有害微生物的入侵和生存,使得土壤中的有益微生物量随着土壤连作年限的增加而逐渐减少,有害微生物量逐渐增加。通过长期积累,植物根际被有害微生物占据,有益微生物的繁殖被抑制,造成严重的土传病害如根腐病、黑腐病、锈腐病、全蚀病、枯萎病等的发生,最终形成连作障碍。地黄野生状态土壤菌群和头茬土壤菌群相类似,其中变形菌门、厚壁菌门占据优势地位,根际富含芽孢杆菌、假单胞菌等有益生防菌,但重茬地黄根际土壤滋生大量病原菌,益生菌群減少,患病地黄根际土壤中总细菌与总真菌的比值由 7.63×109降至 8.70×106,而且不同连作年限地黄根际土壤细菌群落结构也存在一定差异[105-106]。太子参连作后,根际土壤中真菌群落结构与功能多样性都发生明显改变,其多样性指数和均匀度指数随连作年限增加而下降,益生菌含量显著降低,而病原菌种类及多样性增加,尤其是尖孢镰刀菌(Fusarium oxysporum)含量升高幅度较大[107]。
3 药用植物连作障碍的防控措施
3.1 合理施肥
3.1.1 施用有机肥 无机肥料使全球氮磷循环远远超出了安全运行空间,对生物多样性、人类健康和大气产生了负面影响[108-109]。有机肥是我国传统农业的主要肥料,每年因养殖业产生大量的禽畜粪便,将这些废料资源作为主要原料加工成有机肥施入土壤,不仅解决了粪便污染问题,还可以提高土壤生态系统的抗逆性和恢复力,有利于维持农业生态平衡[110]。有机肥质地细腻,几乎适用于各种作物和大部分土壤,富含多种营养元素及生理活性物质,能起到培肥保墒的作用,长期施用可有效改善地力,增加土壤中氮、磷、钾等元素的含量,促进有机质更新,可使作物明显增产;此外,其分解会抑制病原菌的繁殖与传播,增加细菌、放线菌数量,调节土壤pH值、生理缺素等症状,显著减轻病虫害的发生;同时能够降低土壤有机、无机复合度和土壤容重,有效减轻土壤板结严重等问题[111]。陈希等发现,在育苗与大田移栽时都施用生物有机肥,不仅能有效减小滁菊根际土壤尖孢镰刀菌数量、提升土壤酶活性,而且能显著促进滁菊生长,提高其产量及质量[112]。王博林等发现,施用蚕沙发酵肥显著提高了白术有效成分含量,且产量也有所增加,从而有效缓解了白术连作障碍[113]。石雷磊等在对杭白菊的研究中也发现了同样的结果[114]。
3.1.2 施用微生物菌肥 微生物菌肥是指以致病因子为靶标,筛选高效促生防病抗逆有益微生物菌株,将获得的菌株制成菌剂,添加到有机物料中进行发酵,并将其用于改善土壤环境的一种有机肥料[2]。微生物菌肥中包含大量的微生物活体,除具有一般肥料的功能之外,可以明显增加土壤的微生物量,构建稳定健康的土壤微环境。微生物菌肥施用后,有益微生物提前定殖于植物根际,不仅促进了植物的生长发育,而且对植物病害具有生物防治作用。多种微生物可以诱导植物产生PPO、POD、脂氧合酶(LOX)和苯甲氨酸解氨酶(PAL)等参与植物防御反应,有利于防病抗病[115]。有些微生物还可以分泌抗菌素类物质,形成优势种群来抑制其他种类微生物的生长,阻止了病原菌的入侵和扩散,有效缓解了药用植物的连作障碍。吴莉雅等发现,施用菌肥处理对太子参的连作障碍有良好的缓解效果,能够提高重茬种植太子参的品质[116]。沈宝宇等发现,施用微生物菌肥能不同程度地使豌豆(Pisum sativum L.)株高、节荚高度、结荚节数、单荚粒数、有效荚数、单株产量和百粒质量增加,能够有效增产[117]。
3.2 改良耕作制度
3.2.1 轮作 一种药用植物可以和其他科的药用植物轮作,也可以和非药用植物轮作,轮作是缓解药用植物连作障碍问题最简单且方便的方法。轮作后,可有效避免药用植物的化感自毒现象,一种药用植物产生的化感物质对同种或相似种植物有一定的迫害,但是对其他植物不一定存在伤害,并且轮作植物不同,效果也不同,需要通过不断探索来寻求最合适的轮作植物才能将损失最小化,利益最大化。沈玉聪研究阿魏酸等酚酸类物质及三七总皂苷等皂苷类物质对玉米(Zea mays L.)、小麦(Triticum aestivum L.)、白菜(Brassica rapa var. glabra Regel)的化感抑制强度发现,小麦更适合作为三七的轮作作物[118]。轮作后由于植物根系分泌的物质不同,改善了土壤环境,有利于提高微生物的种类和数量,抑制病原菌繁殖,重建土壤微生态的平衡。黄芪轮作后,优化了根际土壤细菌群落α多样性指数,同时还保留了较多的特异OTU数,表明轮作处理可以提高根际细菌多样性及丰富度[4],并且提高黄芪根际土壤酸性磷酸酶(ACP)和URE活性,降低根腐病发病率,提高黄芪根冠比及毛蕊异黄酮葡萄糖苷、芒柄花苷含量[119]。
3.2.2 间作套种 间作套种是对土地资源和种植空间进行充分利用的农业生产模式,又称立体农业[120]。由于不同植物根系分泌物不同且需要的养分也有所差异,因此该方法可以避免土壤养分偏耗问题,亦不会引起土壤微生物失衡,可以维持土壤微生物多样性。并且植物交替种植就像隔离带、防火墙一样,通过阻隔病害的发生、发展和流行,构成了天然的物理屏障。王鹏等研究发现,桔梗与大葱间作后,土壤微生物总量、细菌数量增加,真菌数量减少,并且土壤细菌数量与真菌数量的比值升高[121]。此外,间作栽培在空间上实现了立体化搭配,增加了间作植物叶面光合有效辐射,有利于提高作物的群体光能利用率和群体光合效率[122]。曾文丹等研究发现,木薯(Manihot esculenta Crantz)间作套种栽培技术,不仅可提高复种指数、延长光合作用时间,同时能够改善田间的气候环境,减少病虫害的发生和除草剂的使用,降低种植成本,增加单位土地面积的经济效益[123]。
3.3 品种的选育
不同药用植物对连作的耐受能力不同,即使是同种药用植物,其耐受能力在不同环境下也有变化,有的植株抵抗不良坏境影响的能力要强,并且这种抗性具有遗传性。吕丰娟等通过对芝麻萌发期和田间耐性进行筛选研究,成功获得耐连作能力较为稳定的金黄麻、武宁黑芝麻[124]。如果能够掌握药用植物耐连作的分子机制,就能通过选种、选配、品系繁育和定向培育等技术措施,将抗旱、抗酸、抗盐、抗病虫害、抗连作等优良性状保留下来,并不断强化,从而有效解决连作障碍问题。此外,品种选育对药用植物的进化有巨大作用,对现实生产也有实际意义,是最有效的方法,但也是较困难的方法。
3.4 土壤灭菌
药用植物连作后,由于改变了土壤坏境,使大量病原微生物滋生,受植物分泌的物质诱导,生活于土壤中的植物病原性真菌、细菌、病毒和线虫侵染植物根、茎部,导致土传病害十分严重。而土壤灭菌就是为了使土壤坏境不利于病原微生物的进一步繁殖或是直接将其杀灭,达到控制病原微生物、缓解连作障碍的目的,其方法可分为物理方法和化学方法。物理方法主要有高温闷棚、蒸汽消毒、热水消毒、火焰消毒、太阳能消毒和深翻土壤等[2]。任永权研究发现,对15、8年连作土壤进行热处理后,太子参产量最高分别增长了545.16%、139.29%[125]。连作土壤于60 ℃处理6 h,或在较高温度(90 ℃或120 ℃)下处理3 h即能达到显著的增产效果。化学方法主要是施用化学消毒剂,如氯化苦、棉隆、1,3-二氯丙烯、黄腐酸钾、甲基碘、异硫氰酸甲酯、异硫氰酸烯丙酯、环氧丙烷、威百亩、二氧化硫、叠氮化钠、硫酰氟、石灰氮等[2]。王峰等发现,用氯化苦消毒土壤后,三七主要致病菌假单胞菌属、鞘脂单胞菌属细菌占比均大幅下降,而对三七生长有益的伯克氏菌属、节杆菌属细菌占比则相对增高,并且三七种植后轮作作物烤烟的生长情况也得到极大改善,有助于三七种植后土地的自然修复,从而提高三七种植后土地的利用率[126]。但土壤灭菌存在杀死益生菌的风险,需要慎重考虑和选择。
4 存在的问题和展望
尽管目前药用植物连作障碍问题已经受到众多研究者的关注,并取得了诸多成果,但相较于其他作物的研究仍滞后,且研究不够深入和系统,未能揭示其中的内在机制,因此无法从根本上解决该问题,仅能做到缓解。在缓解措施上,大多数研究仅对单项技术进行分析,鲜有通过多种手段同时施用来进行处理的研究,存在较大局限性。药用植物相较于其他作物而言更加注重其内在药效成分,对于栽培技术的要求更加严格,因此对于连作障碍发生后的环境修复也更加必要。但更重要的应该是药用植物栽培过程中的动态监控,应尽量避免连作障碍的发生,而目前该方面的研究十分匮乏,需要科研人员与时俱进,不断融入新的方法与技术手段才能实现。
总的来说,连作障碍是植物、土壤、微生物3个因素共同作用的结果,而根际恰是三者的结合之处。根际被认为是植物的第2个基因組,是根-土-微生物相互作用的热点[127]。根际错综复杂的关系表明,三者均可被操纵或设计成有利于植物的方向,以实现可持续的农业收益[128]。由此可见,根际将是未来研究的热点和难点,也是解决药用植物连作障碍问题的关键。此外,药用植物不同,可能导致其连作障碍的主要原因不同,且和地域特征有一定的关联,因此需要有针对性地对不同因素主导的连作障碍问题采取相应的措施,同时也可以采用多种手段,达到有效和快速缓解连作障碍的目的。药用植物连作障碍是一个非常复杂的问题,需要多学科交叉融合才有望将其解决。药用植物连作障碍是在大规模栽培的情况下才产生的,野生药用植物根本不会出现连作障碍或者其连作障碍没有被人们认识到,是中医药大发展背景下出现的问题,需要我们不断去探索和发现其中的奥秘,才能更好地推动中医药的持续健康发展。
参考文献:
[1]林茂兹. 太子参连作障碍及其分子生态学机制研究[D]. 福州:福建农林大学,2010.
[2]吴红淼,林文雄. 药用植物连作障碍研究评述和发展透视[J]. 中国生态农业学报(中英文),2020,28(6):775-793.
[3]李 茜. 温郁金和蓬莪术的化感作用研究[D]. 福州:福建农林大学,2010.
[4]李冰圳. 连作及轮作对蒙古黄芪根际微生物多样性及其品质的影响[D]. 呼和浩特:内蒙古大学,2019.
[5]古 力,牛苗苗,郑红艳,等. 连作地黄的植株形态生理效应研究[J]. 中药材,2013,36(5):691-695.
[6]刘 伟,张 琳,章云云,等. 不同连作年限对白花丹参生长及其活性成分含量的影响[J]. 中国中药杂志,2013,38(24):4252-4256.
[7]熊鹏飞. 不同连作年限白术的化感作用及其对生长与产质量的影响[D]. 贵阳:贵州大学,2016.
[8]孙 鹏,尹海波,姚 佳. 基于连作障碍条件下穿山龙皂苷类活性成分的含量变化[J].实用药物与临床,2016,19(3):339-342.
[9]何丽平,吴友根,张军锋,等. 连作广藿香挥发油及百秋李醇含量的变化[J]. 热带生物学报,2017,8(2):169-173.
[10]檀国印,杨志玲,袁志林,等. 药用植物连作障碍及其防治途径研究进展[J]. 西北农林科技大学学报(自然科学版),2012,40(4):197-204.
[11]简在友,王文全,孟 丽,等. 人参属药用植物连作障碍研究进展[J]. 中国现代中药,2008,10(6):3-5.
[12]祖勒胡玛尔·乌斯满江. 甘草连作、轮作不同种植模式对土壤理化性质与微生物数量的影响[D]. 乌鲁木齐:新疆农业大学,2016.
[13]Guo J H,Liu X J,Zhang Y,et al. Significant acidification in major Chinese croplands[J]. Science,2010,327(5968):1008-1010.
[14]Zelm E V,Zhang Y X,Testerink C. Salt tolerance mechanisms of plants[J]. Annual Review of Plant Biology,2020,71(1):403-433.
[15]Rana M,Richard A. J,André L. Approaches to increasing the salt tolerance of wheat and other cereals[J]. Journal of Experimental Botany,2006,57(5):1025-1043.
[16]王惠珍,张新慧,李应东,等. 轮作与连作当归光合特性和挥发油的比较[J]. 草业学报,2011,20(1):69-74.
[17]Toyooka K,Goto Y,Asatsuma S,et al. A mobile secretory vesicle cluster involved in mass transport from the Golgi to the plant cell exterior[J]. Plant Cell,2009,21(4):1212-1229.
[18]师桂英,孙鸿强,于彦琳,等. 连作栽培对兰州百合(Lilium davidii var. unicolor)叶片PSⅡ光化学效率和抗氧化作用的影响[J]. 中国沙漠,2020,40(2):206-213.
[19]安 艳. 半夏连作障碍影响因子研究[D]. 兰州:甘肃农业大学,2018.
[20]张 菊,李艳萍,赵 远,等. 药用植物连作障碍及其缓解措施研究进展[J]. 云南中医中药杂志,2019,40(11):86-88.
[21]高桐梅,吴 寅,李春明,等. 芝麻连作对农艺性状及土壤生化特性的影响[J]. 土壤通报,2016,47(4):897-902.
[22]吴凤云. 三七连作对土壤酶活及土壤化学性质的影响研究[D]. 昆明:云南师范大学,2017.
[23]刘姣姣,何 静,陈 伟,等. 花椒连作对土壤化学性质及酶活性的影响[J]. 分子植物育种,2019,17(22):7545-7550.
[24]Floch C,Capowiez Y,Criquet S. Enzyme activities in apple orchard agroecosystems:how are they affected by management strategy and soil properties[J]. Soil Biology and Biochemistry,2009,41(1):61-68.
[25]Wei X D,Zou H L,Chu L M,et al. Field released transgenic papaya effect on soil microbial communities and enzyme activities[J]. Journal of Environmental Sciences,2006,18(4):734-740.
[26]Aon M A,Colaneri A C. Temporal and spatial evolution of enzymatic activities and physico-chemical properties in an agricultural soil[J]. Applied Soil Ecology,2001,18(3):255-270.
[27]樊芳芳. 連作对高粱生长及土壤环境的影响[D]. 太原:山西大学,2016.
[28]Jian S,Li J,Ji C,et al. Soil extracellular enzyme activities,soil carbon and nitrogen storage under nitrogen fertilization:a meta-analysis[J]. Soil Biology and Biochemistry,2016,101:32-43.
[29]马艳丽,王 鹏. 连作对土壤酶活性影响的研究进展[J]. 防护林科技,2013(4):34-35,61.
[30]李奉国,马龙传,孔 勇,等. 连作对大蒜土壤养分、微生物结构和酶活的影响[J].中国农业科技导报,2019,21(1):141-147.
[31]许永利. 生姜连作土壤生态系统调控研究[D]. 北京:中国农业大学,2014.
[32]李 婧. 连作障碍对栝楼生理活性的影响及其根际土壤环境的动态探究[D]. 金华:浙江师范大学,2018.
[33]孙鸿强. 连作对兰州百合生理特性及土壤环境效应的影响[D]. 兰州:甘肃农业大学,2017.
[34]孙凯宁,杨 宁,王克安,等. 山药连作对土壤微生物群落及土壤酶活性的影响[J]. 水土保持研究,2015,22(6):95-98.
[35]陈 慧,杨志玲,袁志林,等. 白术连作根际土壤的理化性质及微生物区系变化[J].植物资源与环境学报,2014,23(1):24-29.
[36]李振方,杨燕秋,谢冬凤,等. 连作条件下地黄药用品质及土壤微生态特性分析[J].中国生态农业学报,2012,20(2):217-224.
[37]张新慧,张恩和,郎多勇,等. 不同茬口对当归根际土壤酶活性及其产量和品质的影响[J]. 中草药,2011,42(11):2322-2325.
[38]郝建朝,吴沿友,连 宾,等. 土壤多酚氧化酶性质研究及意义[J]. 土壤通报,2006,37(3):470-474.
[39]张新慧,张恩和. 不同茬口对当归根际微生物数量和产量的影响[J]. 中草药,2008,39(2):267-269.
[40]Mahboobi N,Heidarian A R. Allelopathic effects of medicinal plants on germination and seedling growth of some weeds[J]. Journal of Fundamental and Applied Sciences,2016,8(2):323-336.
[41]Mahmoodzadeh H,Ghasemi M,Zanganeh H. Allelopathic effect of medicinal plant Cannabis sativa L. on Lactuca sativa L. seed germination[J]. Acta Agriculturae Slovenica,2015,105(2):233-239.
[42]Molisch H,der Einflusse P A D A. Allelopathie[J]. Nature,1938,3568:493.
[43]王建花,陈 婷,林文雄. 植物化感作用类型及其在农业中的应用[J]. 中国生态农业学报,2013,21(10):1173-1183.
[44]陈冬梅. 作物多样性栽培对烟草连作障碍的生态调控机制[D]. 福州:福建农林大学,2010.
[45]张亚琴,陈 雨,雷飞益,等. 药用植物化感自毒作用研究进展[J]. 中草药,2018,49(8):1946-1956.
[46]郭兰萍,黄璐琦,蒋有绪,等. 苍术根茎及根际土水提物生物活性研究及化感物质的鉴定[J]. 生态学报,2006(2):528-535.
[47]薛兴华. 白术自毒作用的初步研究[D]. 贵阳:贵州师范大学,2016.
[48]刘晓珍,肖 逸,戴传超. 盐城药用菊花连作障碍形成原因初步研究[J]. 土壤,2012,44(6):1035-1040.
[49]柳皓月,刘景坤,谢 敏,等. 云木香根际土壤的主要化感物质及其对植物抗氧化酶系统的影响[C]//中国第九届植物化感作用学术研讨会论文摘要集. 杨凌:中国植物保护学会植物化感作用专业委员会,2019.
[50]黄建国. 黄花蒿的化感效应[J]. 山地农业生物学报,2015,34(4):1-8,17,101.
[51]黄钰芳,张恩和,张新慧,等. 兰州百合根及鳞茎水浸液自毒作用的研究[J]. 草业学报,2017,26(8):93-103.
[52]陈君良. 兰州百合根系分泌物自毒作用的研究及化感物质的GC-MS分析[D]. 兰州:甘肃农业大学,2016.
[53]肖 岚,刘应蛟,徐 贝,等. 玉竹根际土壤化感作用研究及化感物质的鉴定[J].中国农学通报,2015,31(34):163-167.
[54]刘素慧,刘世琦,张自坤,等. 大蒜根系分泌物对同属作物的抑制作用[J]. 中国农业科学,2011,44(12):2625-2632.
[55]郭晓光,杨 琴,陈泽华,等. 浙贝母根系分泌物对其幼苗化感作用的影响[J]. 山西农业科学,2013,41(11):1197-1201.
[56]朱 婧. 丹参化感物质及其化感作用研究[D]. 雅安:四川农业大学,2014.
[57]李贤超. 广藿香植株化感作用及其根际土壤自毒作用与矿质元素含量的研究[D]. 海口:海南大学,2014.
[58]薛 启,王康才,梁永富,等. 藿香不同部位浸提液对其种子萌发及幼苗生长的化感作用[J]. 南京农业大学学报,2017,40(4):611-617.
[59]Zhang S S,Jin Y,Zhu W J,et al. Baicalin released from Scutellaria baicalensis induces autotoxicity and promotes soilborn pathogens[J]. Journal of Chemical Ecology,2010,36(3):329-338.
[60]杨晓燕. 甘草根际土壤化感物质的研究[C]//第21届全国色谱学术报告会及仪器展览会会议论文集. 兰州:中国化学会,2017:812-813.
[61]Kai G,He X,Yan Z,et al. Allelochemicals from the rhizosphere soil of cultivated Astragalus hoantchy[J]. Journal of Agricultural and Food Chemistry,2016,64(17):3345-3352.
[62]黃兴学. 豇豆连作土壤中自毒物质鉴定及肉桂酸对豇豆光合作用的影响[D]. 武汉:华中农业大学,2010.
[63]张秋菊. 三萜人参皂苷对人参、西洋参等植物生长发育的效应研究[D]. 长春:吉林农业大学,2012.
[64]刘芳君. 无土栽培人参根系分泌物的化感作用研究[D]. 长春:吉林农业大学,2014.
[65]任 晶. 人参根际有机酸的化感活性及环境行为研究[D]. 长春:吉林农业大学,2016.
[66]蒋景龙,余 妙,李 丽,等. 西洋参根腐病与人参皂苷积累关系[J]. 中成药,2019,41(9):2164-2169.
[67]焦曉林,毕晓宝,张雪松,等. 不同基质条件下西洋参皂苷提取物的自毒作用[J].中国中药杂志,2015,40(8):1433-1438.
[68]杨家学. 西洋参酚酸类化合物的化感作用及影响因子研究[D]. 北京:中国协和医科大学,2009.
[69]Yang M,Zhang X D,Xu Y G,et al. Autotoxic ginsenosides in the rhizosphere contribute to the replant failure of Panax notoginseng[J]. PLoS One,2015,10(2):e0118555.
[70]吴立洁. 三七根际土壤中酚酸类物质化感作用及其干预措施研究[D]. 北京:北京中医药大学,2014.
[71]向 维. 三七根系分泌物的自毒作用及自毒物质研究[D]. 南宁:广西大学,2016.
[72]钏有聪,罗丽芬,袁 也,等. 皂苷对三七种子的自毒活性及其与结构的关系[C]//中国植物病理学会2016年学术年会论文集. 南京:中国植物病理学会,2016.
[73]王立光,陈 军,李静雯,等. 胡麻残茬水提液化感自毒作用研究[J]. 中国油料作物学报,2019,41(3):445-454.
[74]吕丰娟,肖运萍,汪瑞清,等. 连作对不同抗性芝麻根系分泌物中酚酸类物质含量的影响及其化感作用[J]. 江西农业学报,2020,32(4):1-8.
[75]韩春梅,李春龙,叶少平,等. 生姜水浸液对生姜幼苗根际土壤酶活性、微生物群落结构及土壤养分的影响[J]. 生态学报,2012,32(2):489-498.
[76]张 宝. 地黄化感自毒物质时空分布及根区微生物群落结构研究[D]. 福州:福建农林大学,2015.
[77]谢 敏,燕志强,李秀壮,等. 党参自毒作用研究[C]//中国第八届植物化感作用学术研讨会论文摘要集. 南京:中国植物保护学会植物化感作用专业委员会,2017.
[78]何志贵. 半夏连作障碍发生机制与轮作修复研究[D]. 咸阳:西北农林科技大学,2019.
[79]东莹莹. 怀山药化感作用的初步研究[D]. 开封:河南大学,2014.
[80]白玉超. 苎麻潜在的化感物质对苎麻根际微生物多样性及生理生化的影响[D]. 长沙:湖南农业大学,2017.
[81]母 容,王进闯,王彦杰,等. 花椒自毒作用研究[C]//中国第六届植物化感作用学术研讨会论文摘要集. 成都:中国植物保护学会植物化感作用专业委员会,2013.
[82]张 丹. 基于黄连化感物质与微生物群落结构变化的连作障碍机制研究[D]. 成都:成都中医药大学,2015.
[83]曾令杰,刘 意,禇晨亮,等. 穿心莲化感作用与GAP栽培规范的研究[J]. 现代中药研究与实践,2011,25(3):5-7.
[84]覃逸明,聂刘旺,黄雨清,等. 凤丹(Paeonia ostii T.)自毒物质的检测及其作用机制[J]. 生态学报,2009,29(3):1153-1161.
[85]刘 勇,刘 燕,王星星,等. 头花蓼整株及根际土壤水浸提液的化感自毒作用[J].江苏农业科学,2019,47(17):266-270.
[86]李 键. 木麻黄化感物质对其幼苗生理特征和蛋白质组差异表达的影响研究[D]. 福州:福建农林大学,2013.
[87]刘 奕. 自毒胁迫对木麻黄幼苗根系活性氧代谢及其清除系统的影响研究[D]. 福州:福建农林大学,2014.
[88]袁思安,郭 峰,唐翠平,等. 中国沙棘浸提液对其种子萌发的化感效应[J]. 干旱区资源与环境,2015,29(5):116-121.
[89]惠继瑞. 当归自毒作用研究[D]. 兰州:西北师范大学,2008.
[90]辛爱一,李秀壮,金 辉,等. 当归化感物质及作用机理研究[C]//中国第九届植物化感作用学术研讨会论文摘要集. 杨凌:中国植物保护学会植物化感作用专业委员会,2019.
[91]马红梅,赵培芳. 灵芝连作障碍下的自毒作用[J]. 北方园艺,2016(6):133-136.
[92]鲍红春. 沙芥化感作用的研究[D]. 呼和浩特:内蒙古农业大学,2015.
[93]周 武,汪小玉,李淑红. 药用植物三叶青的化感自毒作用[J]. 丽水学院学报,2015,37(5):26-30.
[94]梁笑婷,林熠斌,宋圆圆,等. 青葙的自毒作用及对其他植物的化感作用[J]. 华南农业大学学报,2018,39(5):32-38.
[95]王馨悦. 自毒物质对辣椒幼苗生长的影响和MT缓解DEP胁迫的生理机制研究[D].南昌:江西农业大学,2019.
[96]Offre P,Spang A,Schleper C. Archaea in biogeochemical cycles[J]. Annual Review of Microbiology,2013,67(1):437-457.
[97]Glaring M A,Vester J K,Lylloff J E,et al. Microbial diversity in a permanently cold and alkaline environment in greenland[J]. PloS One,2015,10(4):e0124863.
[98]Noah F. Embracing the unknown:disentangling the complexities of the soil microbiome[J]. Nature Reviews Microbiology,2017,15(10):579-590.
[99]张 琳,王 晓,史国玉,等. 中药连作障碍形成及其时空效应理论探讨[J]. 现代中药研究与实践,2014,28(5):78-83.
[100]杨建忠,官会林,刘大会,等. 三七连作障碍发生机理及消减技术研究[J]. 北方园艺,2016(14):160-163.
[101]肖春萍,杨利民,马锋敏. 栽培年限对人参根际土壤微生物活性及微生物量的影响[J]. 中国中药杂志,2014,39(24):4740-4747.
[102]张 周,盛 浩,袁 红,等. 玉竹连作对土壤微生物区系的影响[J]. 蔬菜,2019(10):15-20.
[103]Waldrop M P,Balser T C,Firestone M K. Linking microbial community composition to function in a tropical soil[J]. Soil Biology and Biochemistry,2000,32(13):1837-1846.
[104]Nannipieri P,Ascher P,Ceccherini M T,et al. Microbial diversity and soil functions[J]. European Journal of Soil Science,2003,54(4):655-670.
[105]康春晓. 怀地黄连作障碍的微生物学机制及益生菌剂的研发[D]. 新乡:河南师范大学,2017.
[106]吴林坤,黄伟民,王娟英,等. 不同连作年限野生地黄根际土壤微生物群落多样性分析[J]. 作物学报,2015,41(2):308-317.
[107]陈 军. 连作条件下太子参根际关键微生物差异变化机理研究及其应用[D]. 福州:福建农林大学,2016.
[108]Steffen W,Richardson K,Rockstrm J,et al. Planetary boundaries:guiding human development on a changing planet[J]. Science,2015,347(6223):736.
[109]Fowler D,Steadman C E,Stevenson D,et al. Effects of global change during the 21st century on the nitrogen cycle[J]. Atmospheric Chemistry and Physics,2015,15(2):1747-1868.
[110]李水祥. 有机肥替代部分化肥对蜜柚树体营养及果实品质的影响[D]. 福州:福建农林大学,2019.
[111]姜佳琦. 生物有机肥对大蒜连作土壤酶、微生物及养分的影响[D]. 哈尔滨:东北农业大学,2013.
[112]陈 希,赵 爽,史亚东,等. 生物有机肥对‘滁菊连作障碍的影响[J]. 南京农业大学学报,2015,38(1):50-56.
[113]王博林,倪方方,徐建中,等. 蚕沙发酵肥对连作白术产量和品质的影响[J]. 浙江中医药大学学报,2016,40(4):258-264.
[114]石雷磊,徐建中. 蚕沙发酵肥对连作杭白菊药材产量和品质的影响[J]. 浙江中西医结合杂志,2018,28(6):513-515.
[115]鲍 瑾. 微生物菌肥对薄壳山核桃容器苗生长的影响[D]. 合肥:安徽农业大学,2015.
[116]吴莉雅. 微生物菌肥对连作太子参的生长状况及品质提高的探究[J]. 海峡药学,2019,31(11):56-59.
[117]沈宝宇,李 玲. 不同微生物菌肥对豌豆形态性状及产量的影响[J]. 辽宁农业科学,2019(2):88-90.
[118]沈玉聪. 酚酸和皂苷类物质对三七及3种主要轮作作物幼苗的化感作用研究[D]. 郑州:河南农业大学,2016.
[119]孙窗舒. 连作对黄芪品质形成和根际土壤微生物的影响及黄芪轮作换茬方式的研究[D]. 呼和浩特:內蒙古大学,2017.
[120]李图宝. 橡胶砂仁间作套种可行性分析[J]. 农业开发与装备,2019(6):211,228.
[121]王 鹏,祝丽香,陈香香,等. 桔梗与大葱间作对土壤养分、微生物区系和酶活性的影响[J]. 植物营养与肥料学报,2018,24(3):668-675.
[122]侯 慧,董 坤,杨智仙,等. 间作系统根-土互作与连作障碍缓解机制[J]. 中国农学通报,2016,32(29):105-112.
[123]曾文丹,严华兵,谢向誉,等. 木薯间作套种不同作物栽培模式及经济效益研究概况[J]. 农学学报,2016,6(12):11-15.
[124]吕丰娟,肖运萍,汪瑞清,等. 15个芝麻品种(品系)红壤连作耐性分析[J]. 湖南农业大学学报(自然科学版),2018,44(2):130-135.
[125]任永权,张国辉,周江菊. 土壤热处理对连作太子参产量的影响[J]. 北方园艺,2016(7):140-143.
[126]王 峰,吕 艺,刘 洋,等. 氯化苦土壤消毒对三七种植后细菌群落多样性及烤烟轮作的影响[J]. 昆明理工大学学报(自然科学版),2019,44(4):90-96.
[127]Berendsen R L,Pieterse C M,Bakker P A. The rhizosphere microbiome and plant health[J]. Trends in Plant Science,2012,17(8):478-486.
[128]Zhang R F,Vivanco J M,Shen Q R. The unseen rhizosphere root-soil-microbe interactions for crop production[J]. Current Opinion in Microbiology,2017,37:8-14.
