寄主植物对扶桑绵粉蚧体型及体内能源物质含量的影响
2021-09-06桂富荣覃江梅蔡晓燕陈红松
覃 武,胡 雍.,桂富荣,覃江梅,蔡晓燕,陈红松*
1广西农业科学院植物保护研究所/广西作物病虫害生物学重点实验室,广西 南宁 530007;2云南农业大学植物保护学院,云南 昆明 650201
扶桑绵粉蚧PhenacoccussolenopsisTinsley隶属于半翅目Hemiptera粉蚧科Pseudococcidae绵粉蚧属Phenacoccus(武三安和张润志,2009),是一种重要的入侵害虫。该虫原产于北美大陆,现已扩散至全球51个国家和地区(https:∥www.cabi.org/isc/datasheet/109097)。2005—2009年,扶桑绵粉蚧在印度和巴基斯坦暴发成灾,导致棉花减产30%~60%,至此,该害虫已在世界范围内造成严重危害并受到广泛关注(Fand & Suroshe,2015)。在中国,扶桑绵粉蚧于2008年6月在广州首次被发现,目前已分布至广东、江西、湖南、广西、福建、四川、云南、海南、浙江、安徽、江苏、新疆、湖北、河北、上海、天津、重庆、香港和台湾等19个省、市、区(李金峰等,2020;农业农村部,2021)。扶桑绵粉蚧寄主范围广泛,已报道寄主植物267种,包含大田作物、蔬菜、观赏植物、杂草、灌木和树木等(Nagrareetal.,2020)。该虫通过刺吸细胞汁液抑制植物生长,导致植物过早死亡,还会产生蜜露,在叶片上形成烟熏霉菌,从而干扰光合作用(Banazeeretal.,2019)。
寄主植物在害虫暴发成灾过程中发挥着至关重要的作用(Umbanhowar & Hastings,2002)。昆虫取食植物后,通过复杂的生理代谢活动,把植物的营养物质转变成自身身体的构成成分,同时储存能源物质,为其繁殖、越冬及远距离迁飞等生命活动提供能量(韩海斌等,2019)。因此,寄主植物不仅影响昆虫的生长发育和繁殖,而且影响其体内生化物质的组成(Gilesetal.,2002)。昆虫体内的主要能源物质包括可溶性蛋白质、可溶性糖、脂肪等(韩海斌等,2019),取食不同寄主植物后昆虫体内储存的能源物质含量往往存在明显差异(曹玲,2007)。由于物种间营养水平的差异,取食不同寄主后,昆虫的体型也可能存在差异(Parisetal.,2016)。通常,体型大小可以指示昆虫的整体健康状况,大个体通常存活时间更长、繁殖成功率更高、获得的资源更多(Beukeboom,2018)。成虫体型减小,在一定程度上意味着其发育适合度的降低(Chenetal.,2014)。昆虫表型变异可归结为遗传变异和表型可塑性(Zivetal.,2017),昆虫的遗传变异主要发生在较大的地理范围内,表型可塑性主要受营养源影响(Grussetal.,2019)。扶桑绵粉蚧寄主范围广,取食不同寄主后,其生长发育和繁殖参数存在差异(黄芳等,2011;鹿东东和商晗武,2016;吕茂翠等,2013;Nagrareetal.,2018;Sana-Ullahetal.,2011)。取食不同寄主后,扶桑绵粉蚧雌成虫体重也存在显著差异,雌虫产卵量与体重密切相关,虫体越重产卵量越高(Dogaretal.,2018)。目前,尚无关于寄主植物影响扶桑绵粉蚧体内能源物质含量的报道,影响体型的报道仅涉及蛹重和雌成虫体重。本文以扶桑绵粉蚧雌成虫的体长、体宽、体重以及可溶性蛋白质、可溶性糖、脂肪为指标,明确不同寄主植物对扶桑绵粉蚧体型大小和体内能源物质含量的影响,以期为其科学防控提供参考。
1 材料与方法
1.1 供试寄主
选取8种扶桑绵粉蚧常见寄主,其中农作物6种:锦葵科的Bt棉、常规棉Gossypiumsp.;茄科的马铃薯SolanumtuberosumL.、番茄LycopersiconesculentumMiller、茄子SolanummelongenaL.、烟草NicotianatabacumL.。园林观赏植物2种:锦葵科的朱槿Hibiscusrosa-sinensisLinn.、木槿HibiscusmutabilisLinn.。从市场购买朱槿和木槿小苗(高10 cm),移至花盆(21 cm×17 cm)中,置于室外防虫网内,用常规花土培养,长至50 cm高备用。从市场购买Bt棉、常规棉、马铃薯、番茄、茄子、烟草的种子或块茎,播种于花盆(21 cm×17 cm)内,置于室外防虫网内,用常规花土培养,长至5片真叶时备用。每月施适量复合肥(N∶P∶K质量比 = 13∶7∶15),并及时浇水。
1.2 供试昆虫
从广西南宁市街道绿化带朱槿上剪取被扶桑绵粉蚧寄生的枝条,带至实验室(注意隔离,防止粉蚧逃逸,造成人为扩散),取适量扶桑绵粉蚧接种于上述8种寄主上,于(26±1)℃、相对湿度(70±5)%、光周期14L∶10D的人工气候箱中继代饲养5代,取7日龄雌成虫用于实验。
1.3 体型测量方法
取每种寄主各5株,视为5次重复,每次重复取雌成虫100头,用数显卡尺测量单头雌成虫的体长、体宽,用精密电子分析天平测体重。
1.4 能源物质含量测定
可溶性蛋白质:参考王孝平和邢树礼(2009)的考马斯亮蓝染色法测定。取冷冻的雌成虫5头,称重,加入2 mL蒸馏水,4 ℃下充分碾磨,将混浆转移到刻度离心管中,再用6 mL蒸馏水充分洗涤研钵,洗涤液收集于同一离心管中,加水至10 mL,静置30 min后,在4 ℃、3500 r·min-1条件下离心10 min,取上清液,置于4 ℃冰箱内备用。取提取液0.1 mL于试管中,加入5 mL考马斯亮蓝试剂,充分混合,放置2 min后,于595 nm处测光密度,根据标准曲线计算蛋白质含量。
可溶性糖:参考仵均祥等(2004)的硫酸蒽酮法测定。取5头供试雌成虫称重,用1 mL硫酸钠饱和乙醇溶液充分研磨,于1000 r·min-1下离心10 min,弃上清液。将沉淀溶于0.5 mL 70%乙醇溶液,如前述离心,弃上清液。80 ℃水浴加热除去残留的乙醇,然后加入30%氢氧化钾水溶液0.5 mL,100 ℃水浴加热15 min。冷却,加入1 mL无水乙醇,1000 r·min-1下离心10 min,弃上清液,将沉淀溶于1 mL蒸馏水中,加入4 mL蒽铜试剂(0.2 g蒽铜+100 mL浓硫酸),混匀后,100 ℃水浴加热10 min,冷却,于620 nm处测光密度,根据标准曲线计算可溶性糖含量。
脂肪:利用索氏提取器,参考陈永杰等(2005)的差重法测定。每种寄主设5次重复,分别取各寄主上的雌成虫样品置于60 ℃烘箱内烘72 h至恒重,磨碎成粉,称取50 mg磨碎样品(M样),将样品放入抽提器内,样品上面用脱脂棉封盖,抽提瓶称重(M1),接着量取90 mL乙醚加入其中,然后放入水浴槽内,60 ℃水浴抽提12 h,抽提结束后,旋转活塞开关至横向,自动进行乙醚回收,全部无水乙醚回收完成之后,取出抽提瓶,擦净瓶外壁。将抽提瓶放入80 ℃烘箱中烘干24 h至恒重,于干燥器中冷却30 min,再次称重,得到抽提瓶和脂肪的总重量(M2),计算样品中脂肪含量。脂肪含量=(M2-M1)/M样。
1.5 数据分析
用Excel整理数据,用SPSS 21.0 One-Way ANOVA对数据进行方差分析,并用Duncan′s法比较不同处理间平均值的差异程度(P<0.05)。
2 结果与分析
2.1 取食不同寄主扶桑绵粉蚧雌成虫体型
取食不同寄主后,扶桑绵粉蚧雌成虫体长、体宽和体重均存在着显著差异(p<0.05)。取食马铃薯后,扶桑绵粉蚧雌成虫在体长、体宽、体重上均显著优于取食其他7种寄主(表1)。取食马铃薯后,雌成虫体长达4.80 mm,显著长于取食其他寄主的雌成虫,其次为取食番茄(4.45 mm)、茄子(4.31 mm)、朱槿(4.24 mm),取食烟草体长最短(3.52 mm)。取食马铃薯后,雌成虫的体宽为3.07 mm,显著大于取食其他寄主的体宽,其次为取食番茄(2.82 mm)、茄子(2.80 mm)、木槿(2.71 mm),取食烟草最小(2.27 mm)。取食马铃薯后,雌成虫体重为7.11 mg,显著高于取食其他寄主的雌成虫,其次为取食番茄(5.67 mg)、茄子(5.16 mg),取食木槿体重最轻(3.12 mg)。
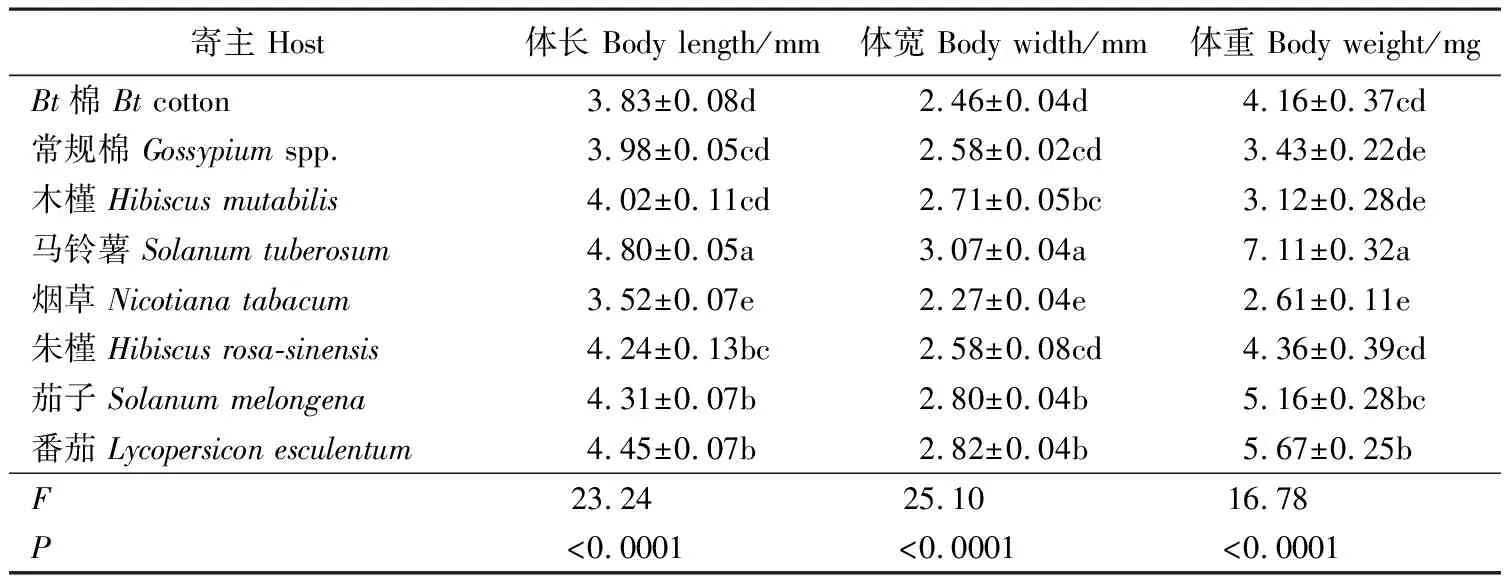
表1 取食不同寄主后扶桑绵粉蚧雌成虫的体型
2.2 取食不同寄主扶桑绵粉蚧雌成虫体内能源物质含量
取食不同寄主后,扶桑绵粉蚧雌成虫体内能源物质含量存在显著差异(P<0.05)。取食棉花(Bt棉和常规棉)后,扶桑绵粉蚧雌成虫体内可溶性蛋白质含量最高(58.77和57.70 mg·g-1),并显著高于取食其他寄主的雌成虫;取食木槿和烟草时可溶性蛋白质含量最低,分别为6.53和4.81 mg·g-1。取食常规棉后,扶桑绵粉蚧雌成虫体内可溶性糖含量最高,为75.02 mg·g-1;其次为取食番茄和木槿,分别为69.00和65.69 mg·g-1;取食茄子时可溶性糖含量最低(30.26 mg·g-1)。取食马铃薯后,扶桑绵粉蚧雌成虫干重脂肪含量最高(349.00 mg·g-1);其次为取食茄子(276.53 mg·g-1);取食Bt棉时脂肪含量最低,为139.17 mg·g-1。总体比较,取食马铃薯后,扶桑绵粉蚧雌成虫体内脂肪含量最高,可溶性蛋白质和可溶性糖含量适中(表2)。
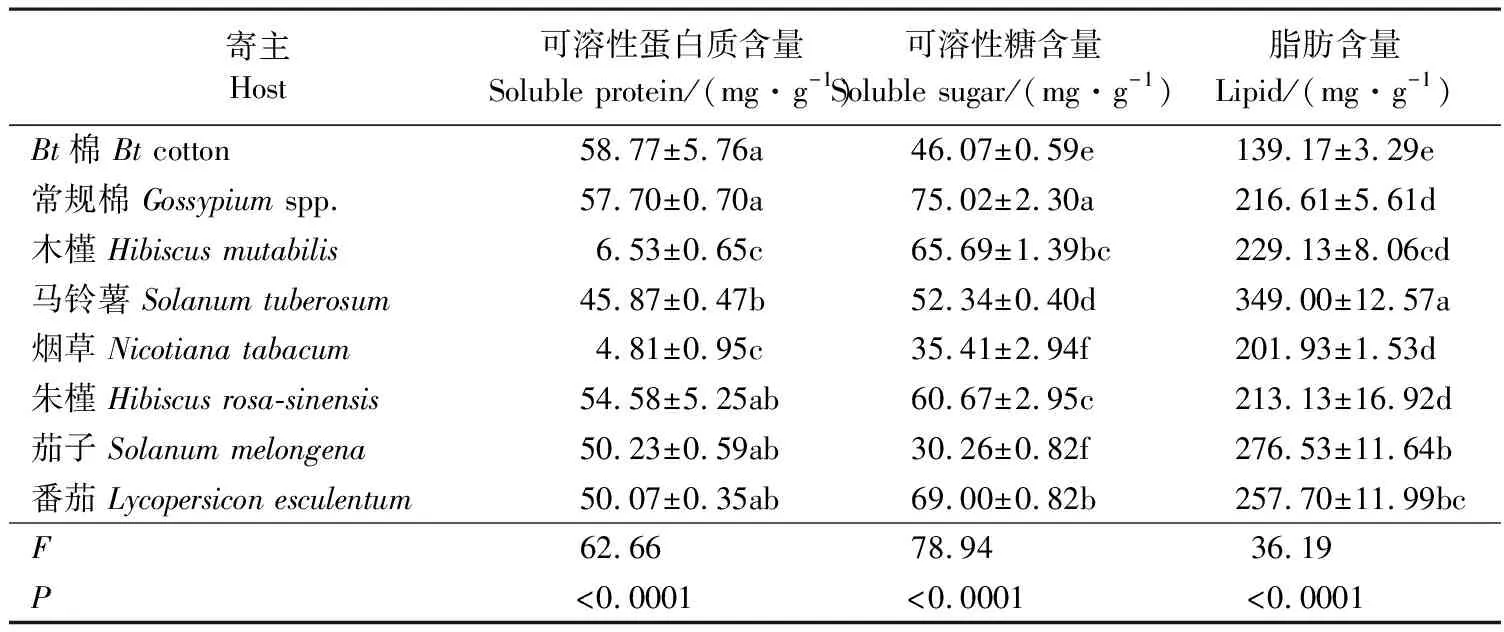
表2 取食不同寄主后扶桑绵粉蚧雌成虫体内能源物质的含量
3 讨论
植食性昆虫所需的营养物质直接来源于寄主植物。因此,植食性昆虫的生长发育、生存繁殖、体型大小和能源物质含量受寄主植物影响。
体型是昆虫最基本的表型特征,从解剖学、生理学到生活史、行为和生态学等方面影响昆虫的适应性(Chenetal.,2014; Henryetal.,2009)。本研究中,取食不同寄主后,扶桑绵粉蚧在体型上存在一定的差异,这可能是由于不同寄主植物营养水平差异所致。取食马铃薯后,扶桑绵粉蚧雌成虫在体长、体宽、体重上均优于取食其他7种寄主,这与朱艺勇等(2011)报道的结果相近,但与黄玲等(2011)及Dogaretal.(2018)的结果差异较大,可能是地理种群、寄主植物、饲养条件等原因所致。Parisetal.(2016)研究发现,寄主植物影响柑橘木虱DiaphorinacitriKuwayama的大小和形状,推测不同寄主饲养的柑橘木虱形态差异,可能由于物种之间营养水平差异所致。西花蓟马FrankliniellaoccidentalisPergande取食黄瓜CucumissativusL.后的体型大于取食菜豆PhaseolusvulgarisLinn.的体型,感性品种上的体型也大于抗性品种上的体型,大个体蓟马繁殖力更强,因此,体型可以作为植物抗性的一个标志(de Kogeletal.,1999)。通常认为,昆虫体型越大,其适合度越高(Chenetal.,2014)。多数昆虫的繁殖力随体型增大而增加,体型可以预测繁殖力,而繁殖力是适合度的重要组成部分(de Kogeletal.,1999)。扶桑绵粉蚧雌虫体重与其产卵量密切相关,取食花烛AnthuriumandraeanumLinden、茉莉花Jasminumsambac(L.)Aiton和朱槿后,扶桑绵粉蚧雌虫较重,繁殖力更强(Dogaretal.,2018)。已报道多种昆虫的产卵量与体重相关,如斜纹夜蛾SpodopteralituraFabricius(Greenbergetal.,2001)、白纹伊蚊AedesalbopictusSkuse和埃及伊蚊AedesgeniculatusOliv(Armbruster & Hutchinson,2002)、小菜蛾PlutellaxylostellaL.(Saeedetal.,2010)、家蝇MuscadomesticaL.(Khanetal.,2012)等。通过食物链能量传递,寄主植物与天敌昆虫的生长发育和形态关系密切(Gols & Harvey,2009; Hopkinsetal.,2009)。如间纹蓟马Aeolothripsintermedius(Bagnall)幼虫和成虫主要捕食昆虫,成虫取食田间花粉补充营养,成虫的体长和体重与寄主植物密切相关(Grussetal.,2019)。可见,寄主植物不仅直接影响植食性昆虫的体型,还通过不同营养级关系,间接影响寄生性和捕食性天敌的体型,从而影响整个生态系统的各个环节。
取食不同寄主后,扶桑绵粉蚧雌成虫体内能源物质含量差异显著,即使取食同一寄主植物,不同能源物质含量差异也较大。总体而言,取食马铃薯后,扶桑绵粉蚧雌成虫体内脂肪含量最高,可溶性蛋白质和可溶性糖含量适中。这与王鹏等(2014)研究桃小食心虫CarposinasasakiiMatsumura所得的结果一致,取食不同寄主后,桃小食心虫幼虫总脂肪、总蛋白和总糖含量差异显著,但取食同一种寄主植物均不能使3种能源物质的含量同时保持最高。寄主植物对植食性昆虫的影响,主要取决于植物组织所含营养成分的种类和质量及昆虫的嗜食程度、消化吸收和转化效率(王鹏等,2014)。甜菜夜蛾SpodopteraexiguaHübner取食不同施肥水平的白菜BrassicachinensisL.后,其幼虫体内蛋白质、可溶性糖、脂类3种能源物质的含量差异显著(王思芳等,2017)。韩海斌等(2019)研究发现,黏虫Mythimnaseparate(Walker)幼虫体内能源物质含量与寄主植物粗蛋白含量呈正相关关系,与寄主植物粗纤维含量呈负相关关系。这些研究结果表明,影响植食性昆虫体内能源物质含量的主要因素是寄主植物的营养水平。
取食马铃薯后,扶桑绵粉蚧雌成虫体型较大,体内能源物质含量较高,从形态学和营养水平方面验证了王伟兰(2014)关于马铃薯是扶桑绵粉蚧适宜寄主的结论。马铃薯受害风险较高,因此应加强马铃薯上扶桑绵粉蚧的种群监测,早发现早防治,减轻该虫对马铃薯的危害和扩散蔓延的风险。
