敦煌李光杏花芽分化与发育研究
2021-08-27张继强罗文莉陈翠莲赵通郭荣王俊杰
张继强,罗文莉,陈翠莲,赵通,郭荣,王俊杰
(1.甘肃省林业科学研究院,甘肃 兰州 730020;2.敦煌市林业技术推广中心,甘肃 敦煌 736200)
花芽分化是果树生殖生长最为关键的发育阶段,分化的结果直接影响果树开花的数量和质量[1-2]。关于花芽分化的研究是果树栽培学的基础,它与大田生产实践联系最为密切,越来越受到人们的重视。科研工作者已经对杏Armeniaca vulgaris的花芽分化特征进行了观测和研究[3-5],取得了很多重要成果。李光杏A.vulgarisvar.glabra[6]是普通杏的果实无毛变种,在我国甘肃敦煌、阿克塞、肃北及新疆哈密等地区都有种植[7-8],其中以敦煌李光杏最为著名。李光杏雌蕊败育严重影响果实产量,目前关于李光杏的研究主要集中在栽培管理、生物学特性、抗寒性等方面[9-12],关于生殖生物学、花芽分化方面的研究相对较少[13-15]。本研究采用摘叶试验、石蜡切片及扫描电镜观测等技术,对李光杏花芽分化过程进行观察研究,以期掌握分化临界期和分化进程,为今后李光杏相关研究和栽培管理提供理论参考。
1 研究区概况
试验在甘肃省敦煌市七里镇三号桥村杏园进行,位于敦煌市西郊,海拔1 130 m;年均降水量42 mm,蒸发量2 500 mm;年均气温9.4 ℃,全年日照3 246 h,年均无霜期152 d,属典型的暖温带干旱性气候。土壤类型为沙壤土,pH值7.8。
2 材料及方法
2.1 试验材料
供试材料为李光杏,树龄10 a,处于盛果期。试验园密度750株/hm2,与授粉树按5∶1行间配置,授粉品种为五月黄和毛杏。样株高3.5~4.0 m,冠幅3.0~3.5 m,平均产量50 kg/株。
2.2 试验方法
摘叶试验参照黄海等人[16]的研究方法,选取盛果期的李光杏若干株,于枝条停长后进行,以枝条上最后一片叶片刚刚展平为准。2018年5月15、22日,6月1、8、15、22、29日,7月6、14、22、29日,8月6日对标记的样枝进行摘叶处理。试验取3次重复,每个处理标记长、短样枝各50个,每个样枝上有叶5片以上,保证样枝能形成花芽。去叶时除骨干枝的延长梢外,所有叶片全部摘除,只保留顶端1片。不摘叶处理的枝条作为对照。次年4月上旬,采用人工记数法统计样枝的花芽分化数量。
花芽分化进程观察每隔约10 d采样1次,从2019年6月1日开始至10月31日结束。样株树冠外围采取短枝顶芽30个为样芽,FAA固定液固定,小瓶密封保存。实验室内爱氏苏木精整染样芽,常规石蜡纵向切片制片[17-18],中性树胶封片,取中区切片电子显微成像系统观察确认分化时期并拍照。每次最少切片观察样芽15个。参照[文献19-20]标志特征的解剖学描述确定花芽形态分化和发育阶段起止时间。对于特定阶段,以1个样芽呈现标志特征定为始期,以3个样芽呈现标志特征定为盛期,10个样芽呈现标志特征定为末期。
3 结果与分析
3.1 花芽分化时期
图1显 示,5月15、22日 和6月1、8日 摘 叶样枝上,次年都没观察到花芽形成,说明6月8日之前,摘叶完全抑制花芽的形成。在6月15日摘叶样枝上开始有少量花芽形成,并且随着摘叶时间的推移花芽呈现出逐渐增多的趋势。说明花芽形态分化最早开始于6月8日至15日之间,即枝条停止生长5周后开始花芽形态分化。7月6日,即第8周,成花率达到50%以上;7月29日,即第11周,成花率达到最高65%,与对照基本持平,表明这一时期花芽形态分化基本结束。

图1 处理样枝成花率变化
上述结果可以判定,6月8日之前为李光杏的芽体生理分化期,即从枝条停止生长到花芽形态分化开始,这一整个时期都是花芽的生理分化时期。生理分化期的芽体进行着节位增长和复杂的生理转化过程,黄海等人认为这时的芽体处于可塑状态,根据条件不同可以向花芽转化,也可以发育为成熟叶芽;当芽体发育到一定节位时丧失可塑性,或者停止发育而成为叶芽,或者转化为花芽继续花器官的分化和发育[16]。
由于成花率以摘叶处理后次年4月开花数量为统计基础,此时样枝开放的花朵形成于摘叶之前,即其花芽的形态分化完成于摘叶之前,说明摘叶仅仅抑制李光杏芽体向花芽的转化,即花芽形态分化,而没有阻断花芽的发育过程,即其萼片、花瓣、雄蕊、雌蕊等花器官的分化和发育。需要说明的是,花芽形态分化在狭义上指芽体转化为花芽的过程,在广义上还包括花器官的分化和发育,即花芽发育。为此,本文仅在狭义上使用花芽形态分化一词,而将跨越花芽形态分化期之后的花器官分化和发育统称为花芽发育。
试验中发现,摘叶处理的样枝当年至次年夏季几乎光秃无叶,未见侧芽萌发抽枝。说明摘叶处理阻断了芽体继续发育为成熟叶芽的过程,即摘叶阻断叶芽发育。
3.2 花芽形态分化与发育进程
镜检观察结果表明,样芽形态分化与发育进程分为以下几个阶段。
(1)花芽形态分化先期。在6月20日以前,样芽体积较小,纵切面可见生长点呈锥状,被鳞片紧抱,分生细胞体积较小(封二图1)。
(2)花芽形态分化期,标志特征是花原基形成。样芽鳞片松开,纵切面可见生长锥膨大,向上隆起呈馒头状,即芽体生长锥转化为花原基(封二图2)。花芽形态分化初期6月20日,盛期7月20日,末期8月10日;8月10日仍观察到少数芽体处于花芽形态分化期。
(3)花萼分化期。标志特征是花原基顶部平坦,中心部分相对凹下,四周产生突起,即分化形成萼片原基(封二图3)。花萼分化初期7月20日,盛期8月10日,末期8月30日。
(4)花瓣分化期。标志特征是伸长的萼片原基内侧基部产生突起,即分化形成花瓣原基(封二图4)。花瓣分化初期8月10日,盛期8月30日,末期9月10日。
(5)雄蕊分化期。标志特征是伸长的花瓣原基内侧基部出现多个突起体,上下两层排列,即分化形成雄蕊原基(封二图5、图6)。雄蕊分化初期8月20日,盛期8月30日,末期9月10日。
(6)雌蕊分化期。标志特征是雄蕊原基形成时,花原基中心部位向上突起,即分化形成雌蕊原基(封二图5、图6)。雌蕊分化初期8月20日,盛期9月10日,末期9月30日。
(7)雌、雄蕊发育期。从10月开始,整个花芽体积增大,花粉囊壁内出现花粉母细胞,呈多角形且紧密状排列,同时雌蕊原基膨大形成子房,子房内分化形成胚珠原始体(封二图7、图8)。
3.3 花芽形态分化与发育特点
结合摘叶试验和样芽切片显微观察,整理李光杏花芽形态分化与发育历程如表1。在整个花芽形态分化与发育期,花原基分化历时最长,平均51 d;其次为萼片分化期和雌蕊分化期,约42 d;花瓣分化历时较短,约31 d;雄蕊分化历时最短,约21 d。观察还发现,成龄树样芽开始形态分化期的时间明显早于幼龄树。不同结果枝上的样芽进入形态分化与发育各阶段的时间和历时长短也存在差异,一般短果枝条上的样芽较早,长果枝上的样芽相对较晚。
由表1可见,样芽雄蕊、雌蕊均在8月20日开始分化,雄蕊分化盛期出现在8月30,至9月10日结束;雌蕊分化盛期则出现在9月10,至9月30日结束,说明雄蕊分化迅速而集中,雌蕊分化缓慢而离散。显微观察进一步发现,从10月份开始,雄蕊、雌蕊开始发育。
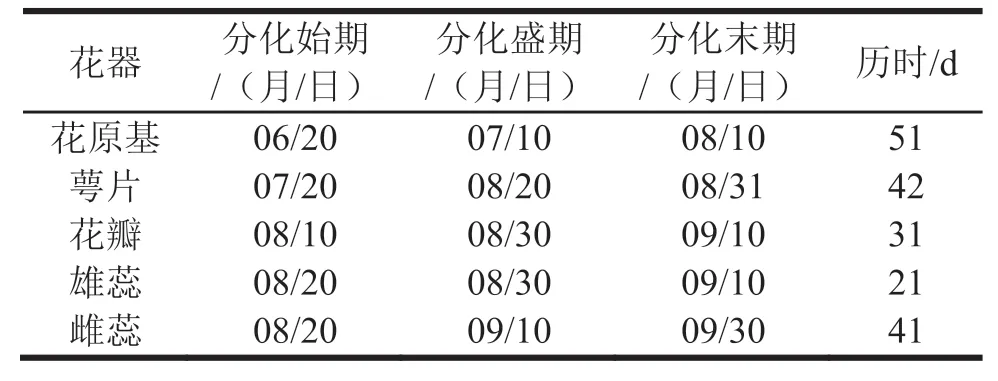
表1 花芽形态分化进程显微观察结果
摘叶试验表明李光杏样株花芽形态分化出于6月15日之前,切片显微观察则表明开始于6月20日。两个结果冲突,分析其原因有2条:一是花芽形态分化开始于芽生长锥不可逆地转化为花原基[16],转化之初形态变化微弱,只有花原基呈现标志特征时才能观察确定;二是摘叶试验样本量大,能较早反映出少量越过形态分化期的花芽,切片显微观察样本量小,取样容易漏过较早进入形态分化期的少量花芽。因此,李光杏样株花芽形态分化始期应以摘叶试验为准,切片显微观察确定的始期滞后约10 d左右。同理,花萼、花瓣、雄蕊、雌蕊等花器官的分化期也应较切片显微观察确定的日期有所提前。
4 结论与讨论
在敦煌,李光杏6月上旬完成花芽生理分化期,开始花芽形态分化,7月下旬花芽形态分化基本结束,8月10日仍有少数花芽形态分化;7月上旬开始花器官分化,次序为萼片、花瓣、雄蕊、雌蕊分化和发育,历时102 d左右。雄蕊、雌蕊分化重叠,同时开始于8月20日,雄蕊分化迅速而集中,盛期出现在8月下旬,至9月10日结束,历时21 d;雌蕊分化缓慢而离散,盛期出现在9月10日,持续至9月底;10月初开始雌、雄蕊发育花粉和珠心细胞分化。李光杏雌蕊败育不是雌蕊未能分化而是分化后的发育异常。摘叶抑制花芽形态分化但不阻断花芽发育。
帅焕丽[22]对陕西关中地区11个杏品种研究发现,萼片分化所需时间最长,约需35~51 d,而花瓣和雌蕊分化时间最短,需要27 d左右。本文研究结果与帅焕丽不一致,可能是品种之间遗传差异所致[16],也可能是研究对象所处地域不同,温度、湿度、土壤等生境不同所致[23-24];刁永强等人[25-26]认为树体营养状况和管理措施也会导致花芽形态分化与发育各阶段开始时间及持续期的不同。
蔺创业[9]多年观察发现,敦煌李光杏有分批开花的特性,第1批花开放于4月上旬,数量大;第2批开放于4月中下旬,数量较少,均着生于1 a生枝基部或细弱枝上,从未见坐果。本项研究表明李光杏花芽形态分化集中在7月中旬,但迟至8月10日仍有少数花芽处于形态分化期,但9月底花芽均能完成雌蕊分化。这些观察事实在一定程度可以解释蔺创业观察到的李光杏分批开花特性,即其第2批花可能形成于8月上旬之后。
摘叶不阻断花芽而阻断叶芽发育,跨越花芽形态分化期的芽体不受摘叶影响而继续发育并最终开放,这一特性被发展为确定花芽形态分化期的试验手段,但差异背后的机制被严重忽视。发育是有机体从其启始到成熟的自我构建过程,离不开充足的营养供应。摘叶阻断叶芽发育,说明叶芽营养与同位叶片息息相关,失去同位叶片则失去营养。王俊杰等人认为,根据“栈糖渠模型”,叶片光合产物一部分运输贮存在枝干根条中以供应来年生长,其余则供应其腋芽发育并贮存在腋芽内[27-28]。摘叶不能阻断花芽发育,说明花芽营养与同位叶片无关而与母枝有关。王俊杰提出形成层屏蔽假说解释顶端优势现象,认为叶芽被母枝形成层屏蔽在外,萌动时才穿透母枝形成层建立与母枝的维管联系[29]。借用这一思路,推测花芽形态分化中建立起了与母枝的维管联系,在母枝上扎了“根”,因而能从母枝获得发育所需营养。许明宪等观察发现,花原基中可见初生髓部细胞,甚至初生输导组织细胞,但在叶芽生长锥中不存在异形已分化的细胞[30]。这在一定程度上佐证前述推测属实。
形成层屏蔽假说认为[29],春季根尖合成细胞分裂素随蒸腾液流经木质部向上运输,沿途诱导形成层增殖木质部前体细胞,其在树液中的浓度越积越高,最终超过阈值而击穿形成层屏蔽进入叶芽,促使叶芽萌动并合成生长素。生长素经母枝木质部薄壁细胞向下运输,刺激新根发生并下调根尖的细胞分裂素合成水平,同时沿途促进木质部前体细胞分化,从而建成新生枝与母枝的维管联系,相当于叶芽“扎根”于母枝。生长素浓度越大,分化形成的导管越多越大。树液上升过程中因蒸腾失水细胞分裂素浓度越来越高,在枝条最高处最先击穿形成层屏蔽促使叶芽萌动,获得先发优势;枝条下部的叶芽因此萌动较迟甚至不萌动,屈居后发劣势。先发优势与后发劣势在宏观上就表现为顶端优势。“要想富,先修路”,细胞分裂素诱导增殖木质部前体细胞相当于奠定“路基”,生长素主导前体细胞分化则相当于铺设“高速轨道”,从而满足叶芽抽枝展叶对水分、养分的旺盛需求。借用这一思路,推测花芽的“扎根”过程发生在叶芽发育期间。这期间,芽体利用同位叶片的光合产物一边增生节位,一边合成某种促花激素,即花芽生理分化过程。促花激素浓度超过阈值而诱导生长锥内部分化髓部细胞和初生输导组织,同时释放到母枝,促使母枝形成层活动增殖并分化韧皮部运输组织,与芽体内的初生输导组织连接贯通维管联系,这种维管联系一旦贯通,芽体转而从母枝获取营养,从而跨越形态分化阶段进入花芽发育阶段,因此不再受摘叶影响。
李光杏雌蕊败育应该源于其发育过程与雄蕊之间的营养竞争。花芽“扎根”于形态分化之初,如果这条维管之“根”不够粗大,那么花芽的营养供应就不充裕,难以满足雄蕊、雌蕊同时发育的营养需求,“两败俱伤不如放弃”“两害相较取其轻”,因而优先满足雄蕊发育的营养需求。雄蕊发育只需少量营养就能大量产生花粉,雌蕊利用同样多的营养只能产生少数胚珠。1个花粉与1个胚珠都能生产1粒种子,生殖效率相同。因此在生殖中,雄蕊的营养利用率远远高于雌蕊,优先保证雄蕊发育,即使雌蕊败育也不影响生殖后代。蔺创业观察发现李光杏第2批开放且未见坐果的花朵均着生于1 a生枝基部或细弱枝上[9],在一定程度上佐证了上述推测。细弱枝条,形成层活动能力弱,难以形成深厚的基础供花芽“扎”下粗大维管之“根”;1 a生枝条基部的叶片较小,叶龄较长,光合能力降低,供应同位花芽“扎根”的能力较弱。同样的道理,对于同一枝条上多个花芽来说,先分化的花芽因先发优势而“扎根”粗大,后分化的花芽则因后发劣势而“扎根”细弱。最终,前者雌蕊发育正常,后者雌蕊败育。基于同一思路,推测雄蕊和雌蕊之间的营养竞争同样在于花芽内部的输导组织并受控于某种激素,雄蕊分化早获得先发优势,释放的激素较多,其输导组织相对完善,优先获取营养;雌蕊分化较迟而屈居后发劣势,释放的激素较少,其输导组织相对不完善,拾遗补缺获取营养,因而容易受到营养供应波动的影响而败育。
