3个温带阔叶树种根系长期分解速率研究
2021-08-07王传宽王兴昌
潘 君,王传宽,王兴昌,*
1 东北林业大学生态研究中心,哈尔滨 150040 2 东北林业大学森林生态系统可持续经营教育部重点实验室,哈尔滨 150040
根系是森林生态系统中重要的碳库和养分库,对生态系统碳和养分循环起着重要作用[1- 3]。与季节性集中凋落的落叶相比,温带森林的根一年四季都有可能发生死亡并逐渐分解[4]。植物凋落物向土壤输入的物质,很大一部分来自根系,但根系凋落物在生态系统有机物循环中的作用近年来才逐渐受到重视[5]。根系分解是碳和养分归还土壤的主要途径[6],森林中地下凋落量平均约占年总凋落量的48%[5, 7]。因此,根系分解是养分、碳和能量在生态系统内和生态系统之间、生物圈和大气之间循环的重要基础[8-9]。
与地上凋落物分解相比,根系分解研究还很不足。尽管细根(< 2 mm)[5, 7, 10]分解研究已有很多,但粗根分解动态研究仍非常匮乏,绝大多数根系分解的研究持续时间短,不足以反映根系分解的整个过程。凋落物分解过程分为早期的快速分解阶段和后期的慢速分解阶段[1-2, 8, 11-12]。相对于地上凋落物,根系分解缓慢[13],因此短期研究只能反映早期快速分解阶段情况而高估分解速率。而Moore等[14]对温带、寒带和亚北极生态系统中分解袋法研究时间的调查显示,大多数(69%)的分解袋法研究时间为1—2 a,84%的分解袋法研究时间为3 a或更短,只有5%的分解袋法研究时间超过5 a。此外,很多研究表明细根是根系凋落物库的最大贡献者,细根对土壤养分有效性具有重要作用[5, 7]。然而,有证据表明粗根对土壤有机物质的贡献也不能忽略[5, 10, 15],但以往研究对粗根分解模式知之甚少。而且根系长期分解研究仅局限在细根上[8, 13, 16-17],我国温带森林仍缺乏不同直径根系长期分解研究。
本文通过我国东北地区3个温带树种不同径级根系7 a53 d的分解实验,比较长期和短期根系分解及其随根直径、树种的变化模式。研究结果可为探究地下部分的根系凋落物分解以及物质循环和能量流动提供依据。
1 材料和方法
1.1 研究区域概况
研究区域在黑龙江省东北林业大学帽儿山森林生态站(45°25′N,127°38′E)。该地区属于长白山山系支脉,张广才岭西北部的余脉。温带大陆性季风气候,夏季温暖湿润,冬季寒冷干燥。2008—2018平均年降雨量为676 mm,年平均气温为2.0℃[18]。其主要土壤类型为暗棕壤。
1.2 野外取样与处理
试验地点位于通量塔周围天然次生林,平均海拔400 m,平均坡度为9°。选择主要树种白桦(Betulaplatyphylla,BP)、春榆(Ulmusdavidianavar.japonica,UJ)、水曲柳(Fraxinusmandshurica,FM)[18],研究其不同直径的根系分解速率变化规律。每个树种随机选择3—5株样木,于5月26日春季根系死亡高峰后[19],在0—20 cm土层挖取足够的根混合带回实验室。将取回的根用自来水将其表面附着的泥土清洗干净,明显的死根剪掉,然后将根系分为5个直径等级:< 1 mm、1—2 mm、2—5 mm、5—10 mm、10—20 mm,自然风干。实验采用分解袋法[20],分解袋的网眼大小非常重要[21],本实验选择网眼0.1 mm的分解袋能够防止活根进入,也可避免分解后期根系碎屑掉出分解袋。测量根系风干质量与70℃烘干质量的比值,用于计算根系的初始干质量。对每个树种每个直径等级的根系,称取2—5 g的风干根,放入网眼为0.1 mm的分解袋中并用记号笔记录初始风干质量(精确到0.001 g)。根直径< 1 mm、1—2 mm和2—5 mm各60袋,3个树种共540袋;根直径5—10 mm和10—20 mm各30袋,3个树种共180袋。于2012年6月3日将每个树种根系分解袋埋入各自树种树冠下土壤(10 cm深),把计划同时取样的网袋穿在一条塑料绳上以便后期取回分解袋,每个树种重复5株样木。为减少因分解环境改变而产生的差异,布设分解袋后表层覆盖林地凋落物,尽量恢复原样。按照分解前期取回分解袋的间隔时间较短,分解后期取回分解袋的间隔时间较长,在分解的第1年进行5次取样,取样时间分别为2012年6月30日、7月31日、9月1日、10月1日、11月2日;第2年进行3次取样,取样时间分别为2013年5月9日、7月20日、9月22日;第3、5、8年分别只进行1次取样,分别在2014年09月15日、2016年6月25日、2019年7月26日进行取样,历时7年53天(即7.15 a)。样品取回后先清洗分解袋,70℃烘干,再称取干质量(精确到0.001 g)。
1.3 数据处理
质量保持率是分解剩余干质量占初始干质量的百分比,其值越大分解越慢;分解率为100%减去质量保持率。3个树种根系质量保持率时间动态球形检验(Mauchly′s Test of Sphericity)P<0.05,说明根系质量保持率之间存在时间自相关性,因此采用树种和直径及其交互作用的双因素重复测量方差分析,通过组内和组间变异计算时间、直径、树种以及它们间的交互作用。根据同一树种不同直径质量保持率随时间的变化差异,将根系分解过程分为快速分解阶段和慢速分解阶段。采用Olson指数衰减模型Mt/M0=e-kt[22]拟合质量保持率平均值随时间的变化过程,Mt代表t时刻(a)剩余干质量,M0代表初始干质量,Mt/M0为分解t时刻根系质量保持率(%),即根系在t时刻剩余干质量占初始干质量的比例,t为分解时间,k为根系分解系数。该模型设置初始质量保持率为1,避免了初始质量估计偏差导致的k偏差[23]。k直接反映年分解速率,k值越大表明根系分解越快[3]。除了分解系数,还用最终平均分解率/7.15 a直接计算了根系年分解速率。采用单因素方差分析和Duncan多重比较判断直径、树种间分解系数和年分解速率以及快慢速阶段分解百分数的差异显著性。采用非线性回归分析3个树种分解系数、年分解速率随直径的变化规律。利用SigmaPlot 14.0绘图,用SPSS 22软件进行统计分析。
2 结果
2.1 树种和根直径对根系质量损失的影响
重复测量方差分析表明,时间、树种、直径(仅< 1 mm、1—2 mm、2—5 mm)与树种交互作用、树种与时间交互作用显著影响根系质量保持率,但直径、直径与时间交互作用以及直径、树种与时间三者交互作用均不显著(表1)。

表1 三个树种根系分解的重复测量方差分析结果
7 a53 d分解过程中,3个树种根系质量保持率随时间增加呈下降趋势,且生长季根系质量保持率下降速率明显快于非生长季。在快速分解阶段(0—1.30 a),根系质量损失26%—47%;之后慢速分解阶段(1.30—7.15 a)继续损失12%—33%(图1;表2)。在快速分解阶段,白桦、春榆直径间质量损失差异不显著;但直径影响水曲柳根系质量损失,直径5—10 mm根系质量损失(47%)显著高于直径< 1 mm根系(29%)。在慢速分解阶段,水曲柳根系直径间质量损失差异不显著;白桦根系质量损失为直径5—10 mm、10—20 mm均高于直径< 1 mm、1—2 mm、2—5 mm;春榆根系质量损失则为直径1—2 mm显著低于直径2—5 mm、5—10 mm、10—20 mm。即使慢速分解阶段分解时间远长于快速分解阶段,但春榆各径级根系慢速分解阶段的质量损失均显著低于快速分解阶段(表2)。根系分解7 a53 d后仍未完全分解,直径1—2 mm的春榆根质量剩余达56%,分解最快的水曲柳直径10—20 mm根系仍剩余24%(图1)。

图1 三个树种不同直径根系7 a分解过程中的初始质量保持率比较Fig.1 Initial mass remaining in different root diameter classes of three species during the seven years′ decomposition

表2 三个树种根系快速分解阶段(0—1.30 a)和慢速分解阶段(1.30—7.15 a)的分解百分数
分解百分数为均值±标准差;不同小写字母代表同一树种的根直径间差异显著;不同大写字母代表同一树种和直径分解阶段间差异显著(P<0.05),n=5;BP—Betulaplatyphylla, UJ—Ulmusdavidianavar.japonica, FM—Fraxinusmandshurica
2.2 根系分解系数
方差分析和Duncan多重比较表明根系分解系数随树种和根直径而变(表3)。不同树种同一直径的根系分解系数不同,根直径< 1 mm的3个树种的分解系数中春榆和水曲柳显著大于白桦;而其它根直径则均为白桦最小、春榆次之、水曲柳最大,差异显著性因直径而异,总体上水曲柳根系分解系数显著大于白桦和春榆(图2;表3)。同一树种不同直径之间相比,白桦直径10—20 mm根系分解系数显著大于直径< 1 mm、1—2 mm、2—5 mm、5—10 mm根系;春榆直径< 1 mm根系分解系数显著大于直径1—2 mm、2—5 mm、5—10 mm、10—20 mm根系;水曲柳直径< 1 mm与2—5 mm、5—10 mm、10—20 mm以及直径1—2 mm与5—10 mm、10—20 mm根系间差异显著(表3)。3个树种各直径根系拟合得到的分解系数置信区间较大;白桦和水曲柳方程拟合度均较好,而春榆直径< 1 mm、1—2 mm根系决定系数较小,拟合度较低(表3)。
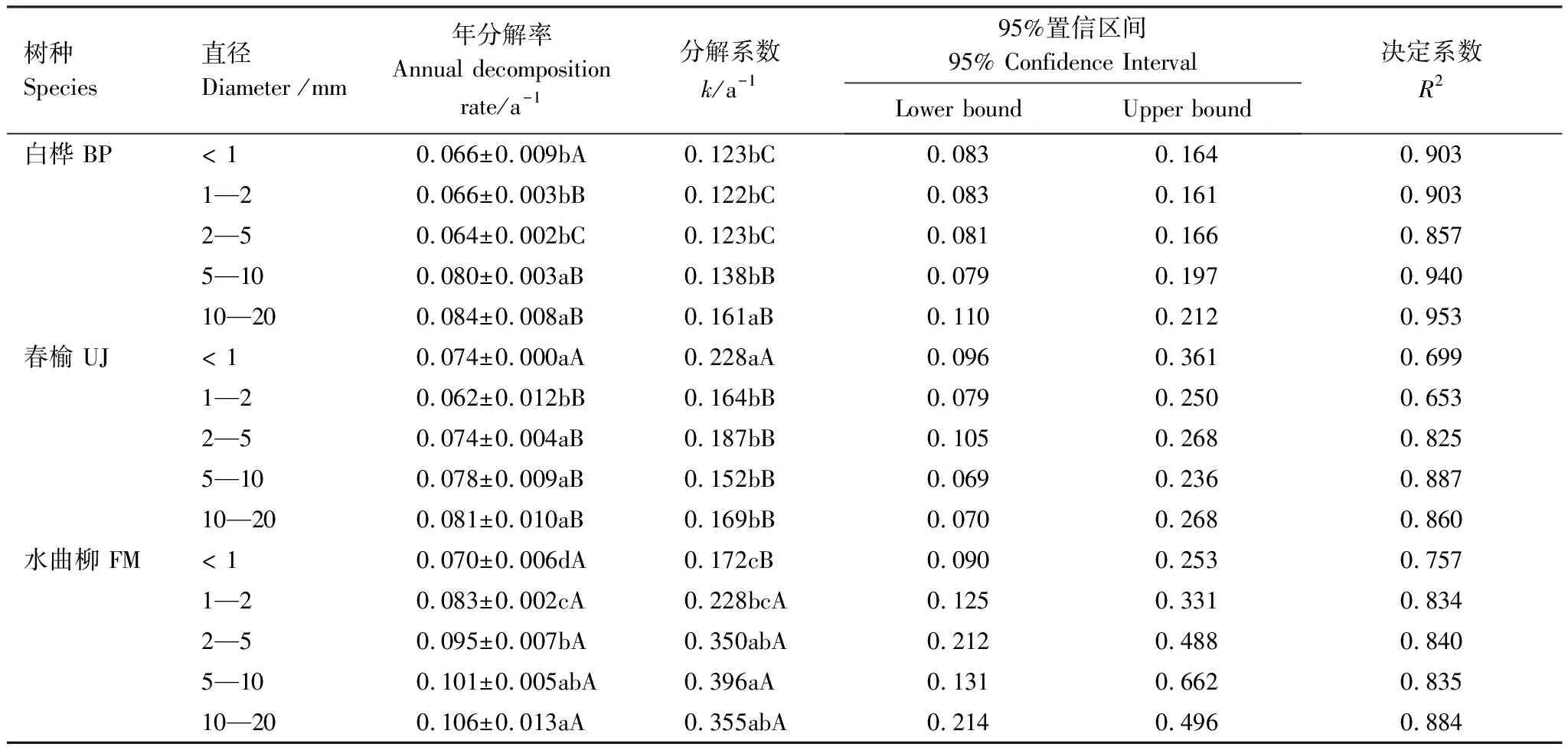
表3 三个树种不同直径根系年分解速率、Olson指数衰减模型得到的分解系数(k)及其95%的置信区间与决定系数
3个树种根系7 a分解系数随直径变化趋势迥异(图2)。白桦根系分解系数与直径(各直径区间的中间值)呈显著线性正相关关系(R2=0.967,P=0.003);春榆各直径根系分解系数与根系直径间的关系不显著(P=0.390);水曲柳根系分解系数与直径呈显著对数正相关关系(R2=0.829,P=0.032)。而短期分解系数则完全不同,白桦、春榆、水曲柳与直径关系均不显著(图2)。7 a平均看,白桦、春榆、水曲柳根系年分解速率随直径的变化(图3)与长期分解系数随直径的变化形式(图2)类似。

图2 三个树种根系长期分解系数和短期分解系数随根直径的变化Fig.2 Change of the long-term and short-term decomposition constants with root diameter for the three tree species实线代表显著,虚线代表不显著
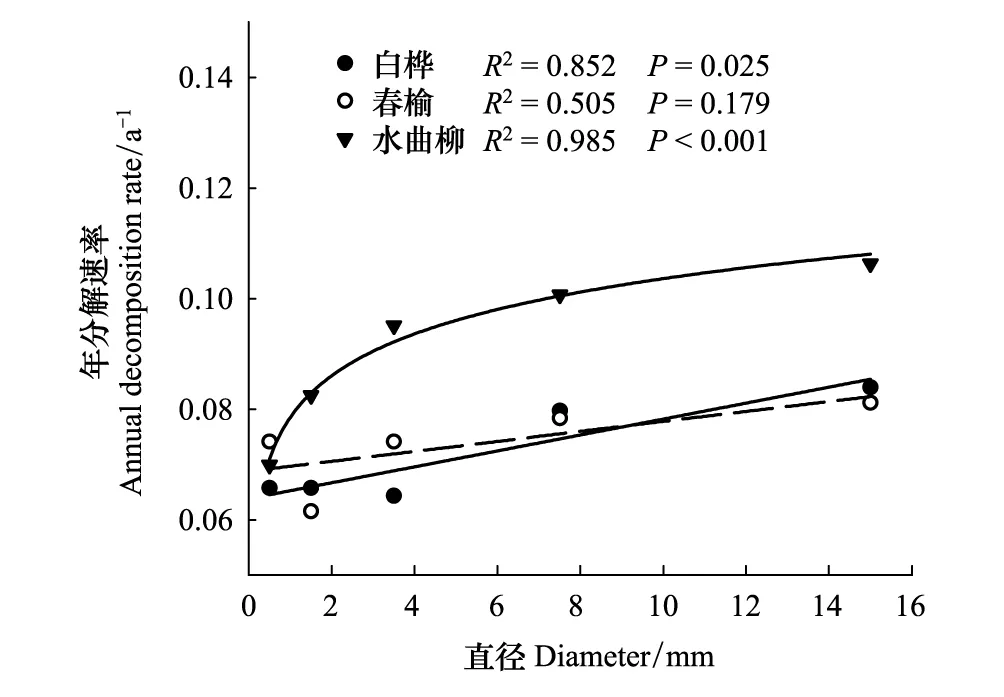
图3 三个树种根系年分解速率与根直径的关系Fig.3 Relationships of the annual decomposition rate with root diameter for the three tree species实线代表显著,虚线代表不显著;年分解速率由分解率/分解时间(a)得到
3 讨论
3.1 根系分解质量损失过程
本研究3个树种不同直径根系在分解初期阶段(0—1.30 a)质量保持率快速降低,之后趋于平缓,这与之前的研究结论相一致[1-2, 11-12]。根系质量损失快慢主要与根系分解在不同阶段养分释放的速率和组分差异有关[2, 8, 15-16, 24],快速分解阶段主要由凋落物中可溶性和相对不稳定的化合物的损失主导,包括初始总非结构性碳水化合物;后期质量损失逐渐减少,酸不溶性物质、木质素对分解有很强的主导作用。生长季气候温暖湿润,淋溶作用增强,微生物酶的活性提高,根系质量保持率快速降低,因而生长季根系质量保持率减少快于非生长季[5, 11, 24- 26];底物质量差异也会影响微生物活性,进而影响根系质量损失速率。
即使经过7 a53 d分解,根系仍剩余相当部分初始质量(24%—56%)。目前长期研究仍然不多[14],为了比较本文3个树种根系分解速率差异,收集文献发表数据作进一步比较。从这些数据可以看出,绝大多数白桦、春榆、水曲柳根系分解研究为短期研究。而短期研究[2, 12, 16, 27]普遍发现根系分解实验结束时仍剩余大部分初始质量。黑龙江省丰林自然保护区35种木本植物的一级根系6 a研究结果[13]与此类似。国外研究表明,10 a分解后根系仍剩余初始质量的0.3%—79.7%[8],寒冷地区12 a分解后仍剩余其初始质量的50%以上[14]。本研究前2个生长季的根系分解系数明显大于整个分解阶段的分解系数,进一步证明了根系短期分解系数会高估根系分解速率,10 a以上的研究结果[8, 14]均表明用短期分解系数估计的凋落物全球储量将被低估至少三分之一。这些研究均表明根系分解是一个长期过程,今后应加强根系分解长期研究(表4)。
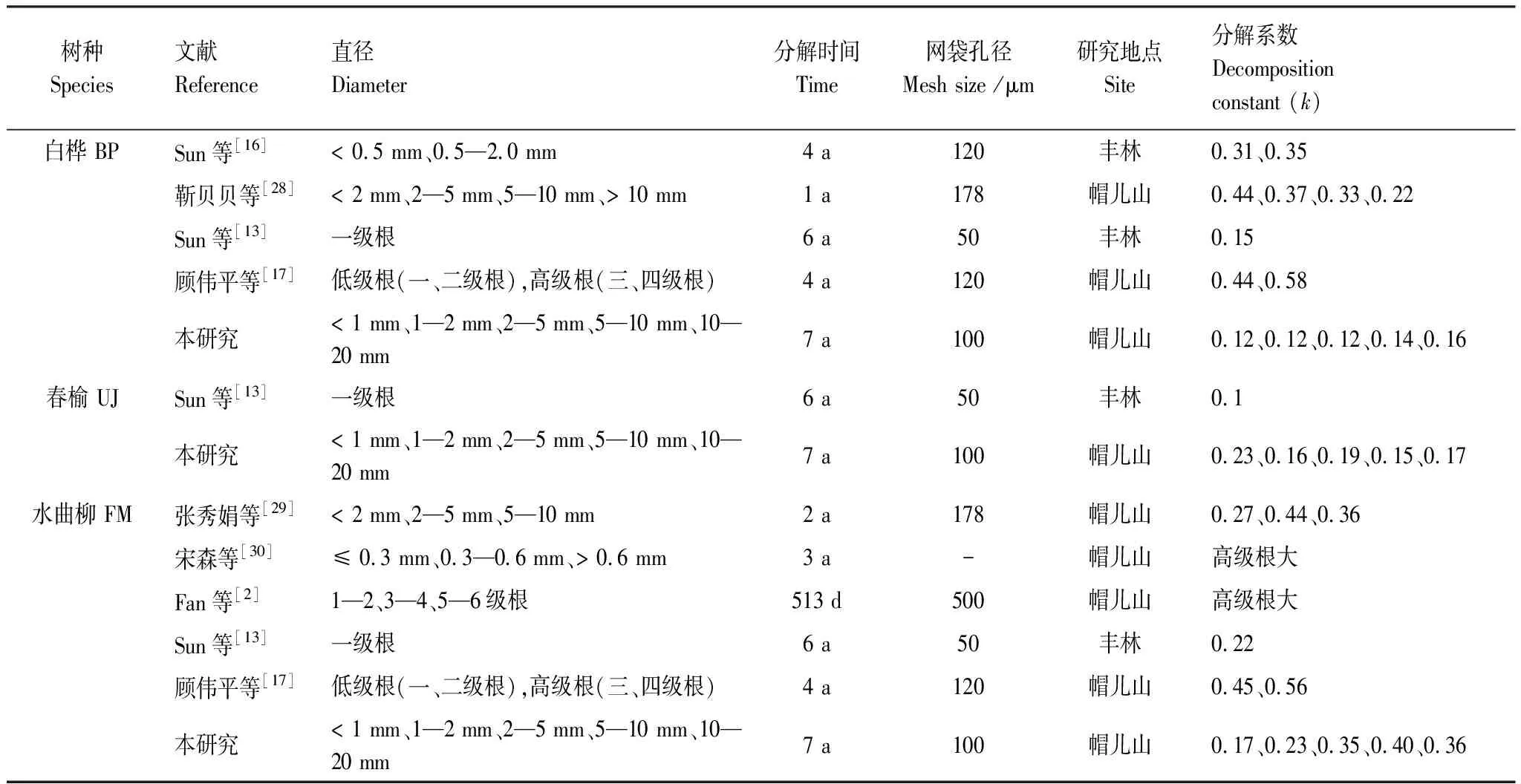
表4 白桦、春榆和水曲柳根系分解相关文献
3.2 根系分解系数种内种间变异
本文和许多文献都表明根系分解速率种间差异很大[11-12, 31]。同一地点不同树种和直径的根系分解研究中,环境因子变异对根系分解的影响较小,而底物质量差异是影响其分解的主要因素[26, 29]。早期研究发现阔叶树根系分解系数大于针叶树[12],如欧洲一项研究发现欧洲桤木(Alnusglutinosa)大于欧洲赤松(Pinussylvestris)和东方云杉(Piceaorientalis)[6],我国东北地区的研究也表明水曲柳细根分解系数大于落叶松(Larixgmelinii)[2],枫桦(Betulacostata)也明显大于红松(Pinuskoraiensis)[1]。这归因于阔叶树的根具有较高浓度的易于分解的水溶性物质,而难分解的木质素浓度较低[12],树种间底物质量不同,从而影响根系分解。最近研究发现根系分解速率还受菌根类型影响,内生菌根树种根系分解速率大于外生菌根树种[2, 13]。本文白桦和春榆为外生菌根,水曲柳为内生菌根[13, 32],同样发现水曲柳分解快于白桦、春榆。此外,菌根类型之间相比,粗根分解也得到了类似规律,但需要更多树种加以验证。
根系分解系数随直径变化因树种而异。白桦和水曲柳根系直径越大,根系分解越快,这与前人对白桦和水曲柳进行研究的结果相一致(表4);而春榆根直径间分解系数没有一致的规律。种间差异可能与Olson指数衰减模型拟合度有关。白桦和水曲柳拟合较好,其分解系数与根系直径具有显著正相关关系;春榆拟合较差,分解系数与根系直径无显著关系。进一步计算根系年分解速率,发现春榆直径间仍没有显著的正相关关系(图3),这基本排除了Olson模型拟合度的影响,春榆根系分解系数与根系直径无显著关系很可能是该树种的固有特性。然而,本文重复之间的差异和Olson模型拟合的不确定性(表3)表明,增加重复数量以及研究局域尺度的根系分解变异对于正确解释凋落物分解机制至关重要[9]。
以往绝大多数细根分解研究也发现根直径越大根系分解系数越大[1, 15, 24, 33- 35],但粗根和细根的分解速率大小仍存在争议[36],且粗根、细根分解的影响因子不同[10, 37]。同一地点,根系分解速率主要取决于底物质量(可溶性碳、N和木质素)差异,N和木质素浓度随直径增加而减小,而N和木质素对根系分解速率有抑制作用[2, 6, 10, 15, 33, 35, 38],因此径级影响底物质量进而影响根系分解速率。也有研究表明分解开始时间与直径存在交互作用[39]。本文不仅为本地区的物质循环提供基础数据,而且粗根分解总体快于细根这一结果对增加根系分解速率的整体认知有重要意义。此外,本文结果也证明粗根分解对生态系统养分循环的贡献不能被忽略。例如,伐根分解可能是采伐初期土壤微生物和植物养分的重要来源[40]。
本研究根系长期分解系数总体小于以往白桦、春榆、水曲柳根系短期分解实验结果(表4)。可能有2个方面原因,一方面可能是因为以往研究时间普遍较短而高估分解系数;另一方面可能与本研究分解袋网眼小有关。网眼大会导致破碎的根系残体在清洗过程中损失,从而高估分解速率,这在后期阶段更加明显;网眼小则会消除这一问题,但可能因为阻止土壤动物的进入而低估分解速率。因此本文的结果可以认为是根系分解速率的下限,而以往大网眼分解袋的短期实验结果可以认为是根系分解速率的上限。而帽儿山春榆直径< 1 mm的细根分解系数大于Sun等[13]的分解系数,这可能与后者研究地点丰林自然保护区的温度低且分解袋网眼直径更小有关(表4)。此外,短期分解系数也不一定能代表长期分解系数的种内、种间差异。短期分解系数和长期分解系数的种内排序存在一定差异,短期分解系数随直径变化趋势与长期分解系数不同。因此,短期研究并不能充分认识整个根系分解过程,这与Moore等[14]研究结果一致,研究表明分解1 a和以后年份的质量损失之间的关系强度随分解时间的延长而减小,说明短期分解系数不能准确的代表长期分解系数。
4 结论
7 a53 d分解过程中,白桦、春榆、水曲柳根系经历快速分解和慢速分解两个阶段。白桦和水曲柳根系分解系数总体随直径增大而逐渐增大,而春榆的分解系数在不同直径间没有显著的变化趋势,总体上细根分解速率低于粗根;不同树种的根系分解速率也存在差异,根直径< 1 mm的分解速率大小排序依次为春榆>水曲柳>白桦,而其它根直径则均按白桦>春榆>水曲柳的种间排序。在分解实验结束后,根系仍剩余相当部分的初始质量(24%—56%),短期根系分解系数明显大于长期根系分解系数,且短期分解系数也不能完全代表长期分解系数的种内、种间差异。本研究7 a分解实验为东北温带森林不同树种根系分解速率随直径的长期变化模式提供了重要数据。
