色季拉山急尖长苞冷杉种群不同龄级立木的空间分布格局
2021-08-06任毅华周尧治方江平罗大庆
任毅华,周尧治,侯 磊,方江平,*,罗大庆
1 西藏农牧学院高原生态研究所,林芝 860000 2 西藏林芝高山森林生态系统国家野外科学观测研究站,林芝 860000 3 西藏高原森林生态教育部重点实验室,林芝 860000 4 西藏农牧学院资源与环境学院,林芝 860000
种群是植物群落的基本组成单位,其空间分布格局一直是生态学研究的热点之一[1- 3]。种群空间分布格局是指种群个体在水平空间的分布状况或配置状况[4],是种群个体自身生物学特性、种内种间竞争关系、种群与环境相互作用过程的外在表现[5-6],对于揭示森林群落结构的形成、演替趋势、种内种间关系及环境适应机制具有重要意义[7-11]。点格局分析法由Ripley于1977年首次提出[12],之后由张金屯引入我国[13]。该方法是以植物个体的空间坐标为基础,将每个个体视为二维空间中的一个点,由此形成植物个体的空间分布点图,在空间分布点图的基础上进行空间分布格局分析,客服了传统方法对样地大小的依赖[2,14],是目前植物种群空间格局分析最常用的方法之一[15]。
急尖长苞冷杉(Abiesgeorgeivar.smithii)是松科冷杉属常绿乔木,为我国特有种,主要分布于我国滇西北、川西南和藏东南地区。在西藏主要分布于察隅、波密、林芝、米林等地[16],是藏东南暗针叶林的主要建群树种之一,对于维系林区生物多样性、保育水土具有重要意义,是西藏生态安全的重要屏障[17]。但由于过去的过度采伐和栖息地的破坏,导致该种群更新困难,自然分布区日益缩减,已被列入国家二级保护植物,处于易危状态[18]。
急尖长苞冷杉在西藏色季拉山多以成过熟原始林存在,人为干扰少,是从事森林生态学研究的理想对象。鉴于该种群重要的生态功能及自身的濒危状态,有关种群生态学研究近年来受到学者们的广泛关注[19- 21]。该种群的空间分布格局也有涉及[22- 23],但相关研究还存在一些不足:一是只分析了固定空间尺度下该种群的空间分布格局,无法全面阐述其空间分布规律;二是在点格局分析中,未将幼苗纳入研究范畴,幼苗作为植物群落的重要组部分,对环境变化异常敏感,其数量特征、分布规律以及对环境的适应能力在很大程度上决定植物种群的发展趋势[21],这对濒危物种更为重要。为此,本研究以西藏色季拉山急尖长苞冷杉原始林为研究对象,将幼苗纳入研究范畴,用点格局法分析该种群的空间分布规律及不同发育阶段在空间分布上的关联性,探讨环境对不同发育阶段个体的影响,以期为该森林生态系统的维持机制及该种群的保护与实践提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
色季拉山位于西藏东南部雅鲁藏布江大峡谷西北侧,属于念青唐古拉山向南延伸的余脉,与喜马拉雅山向东发展的山系相连,主峰海拔高度5200 m,属于亚高山温带半湿润气候区。主要植被类型有高山稀疏垫状植被、以云冷杉为主的暗针叶林、以川滇高山栎(Quercusaquifolioides)为主的常绿硬阔叶林和以杨桦林为主的落叶阔叶林等。试验地位于色季拉山东坡,海拔3800 m,年平均气温2.0—4.5 ℃,最高月(7月)平均气温11.1 ℃,最低月(1月)平均气温-14.0 ℃,极端最低气温-31.6 ℃,极端最高气温24.0 ℃。年均降水1134.1 mm,春夏季节降水占全年79.4%,年均蒸发量544.0 mm,年均相对湿度78.8%。植被乔木层为单一的急尖长苞冷杉纯林,林下灌木主要有猴斑杜鹃(Rhododendronfaucium)、杯萼忍冬(Lonicerainconspicua)、柳叶忍冬(Loniceralanceolata)、西南花楸(Sorbusrehderiana)、长尾槭(Acercaudatum)、冰川茶藨子(Ribesglaciale)、小舌紫菀(Asteralbescens)、峨眉蔷薇(Rosaomeiensis)、光秃绣线菊(Spiraeamollifoliavar.glabrata)等;草本主要有大钟花(Megacodonstylophorus)、五裂蟹甲草(Parasenecioquinquelobus),管花鹿药(Smilacinahenryi)、扭柄花(Streptopusobtusatus)、蛇莓(Duchesneaindica)等。试验地郁闭度0.4—0.9,地形起伏较大,地表苔藓层发达,土壤为漂灰化山地暗棕壤,腐殖质化程度明显。
1.2 研究方法
1.2.1外业调查
森林群落空间分布的点格局分析要求样地以长方形或正方形为宜,且边长在50 m以上,如果小于50 m,则大尺度上的格局有可能被忽略[13]。鉴于此,于2018年11月,在色季拉山东坡海拔3800 m处设置100 m×100 m固定样地,并将其划分为25个20 m×20 m调查单元,记录每个调查单元的坡度、坡向、盖度、植物种类等信息。采用相邻网格法对样地内所有基径≥0.1 cm的急尖长苞冷杉进行调查,并编号。对于树高<130 cm的植株只测基径;对于树高≥130 cm、胸径<15 cm的植株同时测量基径与胸径;对于胸径≥15 cm的植株,只测胸径。此外,记录所有植株的高度、冠幅、坐标及生长状况等信息。
1.2.2急尖长苞冷杉林的龄级划分
林龄结构是植物种群的重要特征之一,但由于树木生长周期长,不可能追踪到所有个体的生活史,因此学者们常用径级来表征植物群落的龄级[24-27]。虽然径级和龄级是两个不同的概念,但二者对环境的反应具有一致性[24]。本研究依据调查结果及种群生长特性,选用基径和胸径两个指标对急尖长苞冷杉种群进行径级划分:首先,对于胸径<10 cm的植株个体,以基径为标准,划分为2个等级,第1级0.1 cm≤基径<3.0 cm,第2级基径≥3.0 cm;对于胸径≥10 cm的植株个体,以胸径为标准,以10 cm为步长值进行径级划分,即第3级10 cm≤胸径<20 cm、第4级20 cm≤胸径<30 cm,等等,此方法将急尖长苞冷杉种群共划分为12个径级。由于植物种群点格局分析对样本数量有一定要求,为客观分析不同发育阶段种群在不同空间尺度上的分布规律及其关联性,参考桂亚可对赣中闽楠(Phoebebournei)天然种群的分类方法[25],并结合种群自身特性,将该种群划分为6个年龄段,分别为幼苗(1级)、幼树(2级)、小树(3—4级)、中树(5—6)、大树(7—8级)、老树(8—12级),如此便保证了不同发育阶段种群的个体数量。
1.2.3数据处理
本研究采用ADE4点格局分析软件分析急尖长苞冷杉种群的空间分布格局及不同发育阶段个体在空间分布上的关联性。该软件的理论基础为Ripley的L函数,L函数由Ripley的K函数改进而来,详细数学原理见参考文献[13, 24, 28- 30]。空间分布格局及空间关联性采用0—50 m空间尺度,步长值为1 m。采用Monte-Carlo检验拟合计算上下包迹线,拟合检验100次,确定99%的置信水平。在点格局分析图中,观测值在上包迹线以上为集群分布,在上下包迹线中间为随机分布,在下包迹线以下为均匀分布;在关联性分析图中,观测值在上包迹线以上为正关联,在上下包迹线中间关联不显著,在下包迹线以下为负关联。用Excel 2016软件绘制点格局分析图。
2 研究结果
2.1 色季拉山急尖长苞冷杉种群的径级与林龄结构特征
样地内共有急尖长苞冷杉431株,分别分布于12个径级上,无缺失径级,最大胸径105.5 cm。冷杉个体在径级上的分布总体呈现倒“J”型(图1),小径木(1级、2级)数量占整个种群的47.80%,即具有较高的更新苗木储备,是典型的稳定增长型种群。该种群在从第1径级到第2径级,第2径级到第3径级数量均有较大幅度的下降,之后趋于稳定状态,直至第10径级又开始大幅下降,可见该种群从第9径级开始趋于老龄化。种群数量在龄级上的分布与径级相似,呈现近似的倒“J”型(图2),其中幼苗、幼树、小树、中树、大树、老树的数量分别为127、79、77、60、61、27株,占种群总数量的29.47%、18.33%、17.87%、13.92%、14.15%、6.26%。
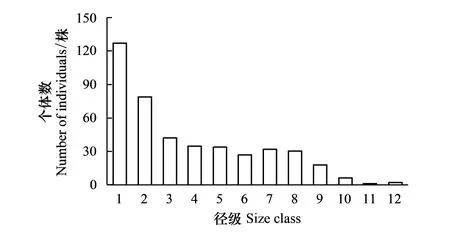
图1 急尖长苞冷杉种群的径级结构 Fig.1 Diameter at breast height (DBH) class of Abies georgei var. smithii forest

图2 急尖长苞冷杉种群的龄级结构Fig.2 Age class of Abies georgei var. smithii forest
2.2 急尖长苞冷杉种群空间分布的点格局分析
对色季拉山急尖长苞冷杉种群进行空间点格局分析发现(图3):色季拉山急尖长苞冷杉种群整体在0—28 m空间尺度内呈现显著的集群分布,在28—40 m空间尺度内呈现随机分布,随着空间尺度的进一步增大,又转为均匀分布,其最高聚集强度为2.412,对应空间尺度为5 m;幼苗在0—42 m空间尺度内呈现显著的集群分布,在42—47 m空间尺度内呈现随机分布,随着空间尺度的继续增大,之后转为均匀分布,其最高聚集强度为10.574,对应空间尺度为16 m;幼树在0—28 m空间尺度内呈现显著的集群分布,在28—42 m空间尺度内呈现随机分布,随着空间尺度的继续增大,又转为均匀分布,其最高聚集强度为6.541,对应空间尺度为5 m;小树、中树、大树、老树在0—50 m空间尺度内均呈现随机分布。从急尖长苞冷杉种群不同发育阶段的空间分布格局看,幼苗聚集强度明显大于幼树,之后的小树、中树、大树、老树均呈现随机分布,说明色季拉山急尖长苞冷杉种群在发育过程中经历了从集群分布到随机分布或均匀分布的演变过程。
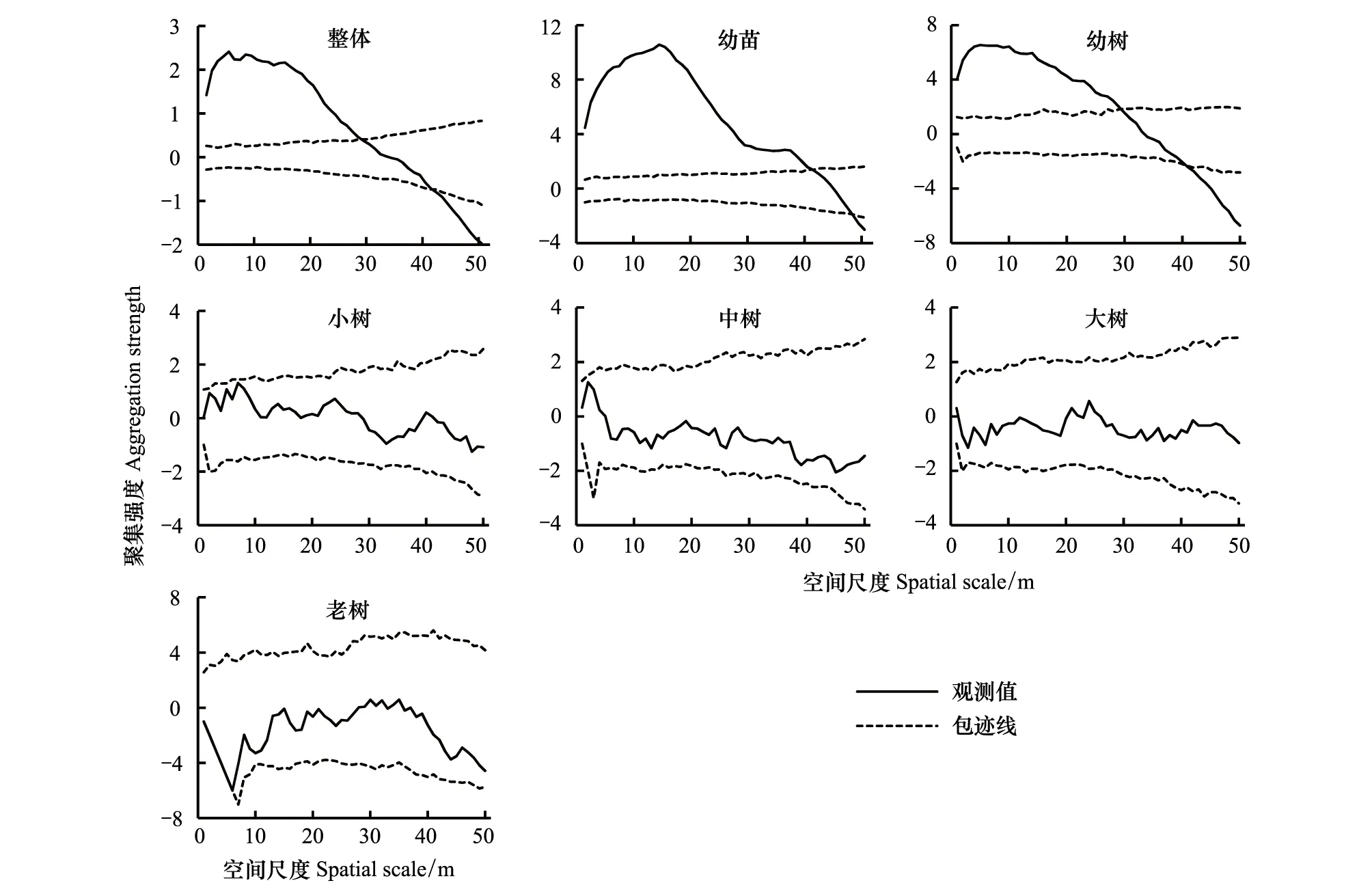
图3 急尖长苞冷杉种群空间点格局分析Fig.3 Point pattern analysis of Abies georgei var. smithii forest
2.3 急尖长苞冷杉种群不同发育阶段的空间关联性
种群不同发育阶段的空间关联性可直观反映种内关系的变化[22,31]。对急尖长苞冷杉种群不同发育阶段空间关联性进行分析发现(图4):幼树、小树、中树、大树、老树在小、中尺度内均与幼苗呈显著的负关联,随着空间尺度的增大,又转为不关联,小树、大树在极窄的大空间尺度内与幼苗还表现出了显著的正关联;小树、中树、大树、老树在空间分布上与幼树空间关联性相对比较一致,即在小、中尺度表现为显著的负关联,随着空间尺度的增大,再依次呈现不关联和正关联;小树、中树、大树、老树相互之间在0—50 m空间尺度内无显著关联。
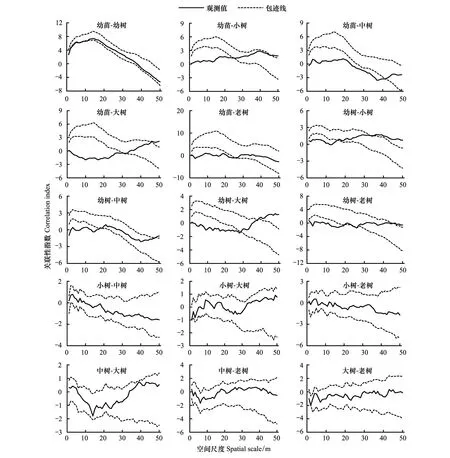
图4 急尖长苞冷杉种群不同发育阶段的空间关联性Fig.4 Spatial associations of Abies georgei var. smithii forest among different growth stages
3 讨论
3.1 急尖长苞冷杉种群的径级结构
植物种群的径级结构是种群生存能力与外界环境相互作用的结果[32],在一定程度上可反映种群的结构现状与更新策略,同时也是探索种群动态和发展趋势的有效方法[27,33]。一般情况下,濒危植物种群幼龄个体少,老龄个体多,多为衰退性[10, 34]。但在本研究中,急尖长苞冷杉种群的径级结构与龄级结构均呈现典型的倒“J”型,幼苗个体数量丰富,随着径级的增大,个体数量逐渐减少,各径级无个体缺失,是典型的稳定增长型种群,与赣中闽楠天然种群[25]、甘南闽江冷杉(Abiesfaxoniana)种群[35]、长白山臭冷杉(Abiesnephrolepis)种群[36]研究结果一致。说明急尖长苞冷杉种群虽为二级濒危物种,但比较适合色季拉山东坡环境。在该环境下,只需控制人为干扰,加强生境保护,种群便可完成自我更新和恢复。
3.2 急尖长苞冷杉种群的空间分布格局
植物种群空间分布格局是种群对环境适应的生存策略及适应机制的反映[37],对空间尺度具有很强的依赖性[13,38-39],同时又受自身生物学特性、发育阶段、种内种间关系、环境等因素综合作用影响[27,39]。一般而言,在较小尺度上,种群的分布格局主要由种子扩散机制、个体繁殖特性以及种内竞争等生物学特性决定;而在较大尺度上,可能受到地形、土壤、水分、光照等环境因素的影响[40-41]。本研究中,幼苗、幼树均随空间尺度的增大依次呈现集群分布、随机分布和均匀分布,且以集群分布为主要特征,而小树、中树、大树、老树则在0—50 m尺度内均呈现随机分布,这种分布格局也出现在其他区域森林生态系统中[10,27,42-44]。究其原因可能是因为:(1)种群初期的分布格局可能与种子传播方式及环境因素有关[45],该种群球果成熟后易脱落、易碎裂,进而形成以母树为中心的集群分布格局,同时集群分布有利于小径木个体间相互庇护,提高种间竞争能力[46];随着空间尺度的增大,又受环境异质性影响,因此在大尺度呈现出随机分布或均匀分布。(2)当种群进入生活史中、后期,植株对光照、水分、养分、空间等资源需求不断增加,种内、种间竞争加剧,自疏与它疏作用增强,导致植株大量死亡,最终趋于随机分布[44]。
此外,本研究中,急尖长苞冷杉种群整体随空间尺度的增加也依次出现了集群分布、随机分布和均匀分布,与幼苗、幼树空间分布规律基本一致,但聚集强度却低了许多,这可能与植株数量在不同龄级上的分布有关。本研究中,该种群数量主要集中于幼苗、幼树上,占种群总数量的47.80%,进而导致种群整体呈现与其相似的分布规律;但因小树、中树、大树、老树均以均匀分布为主,最终导致种群整体的聚集强度明显低于幼苗和幼树。
3.3 急尖长苞冷杉种群不同发育阶段间的空间关联性
同一种群不同发育阶段间的空间关联性能够揭示种群内部个体间在某一时期内的相互作用关系,有助于描述种群的现状与动态[7,47]。本研究表明,小树、中树、大树、老树均在小、中尺度一定范围内与幼苗、幼树呈显著的负关联,在大尺度则呈现一定的正关联(中树与幼苗、老树与幼苗除外),小树、中树、大树、老树之间无显著关联。说明对于急尖长苞冷杉种群,大径木对于林下自然更新在小尺度上起抑制作用,在大尺度则起促进作用,但小树、中树、大树、老树之间无明显相互作用。从径级结构看,该种群在经过幼苗到幼树、幼树到小树,个体数量急剧下降后开始趋于平稳状态,也说明了这点。
本研究中小树、中树、大树、老树与幼苗、幼树在不同空间尺度下的关联性变化基本符合Janzen-Connell假说。该假设认为,种子存活数量与母树的空间距离呈负相关,母树附近的种子和幼苗存活率低,种子死亡率受密度和距离的制约[48-49]。色季拉山急尖长苞冷杉种群大径木个体高大,冠幅发达,对其下种子萌发或幼苗生长抑制作用强,因此在小、中尺度呈现显著的负关联,这与新疆西天山国家级自然保护区天山云杉(Piceaschrenkianavar.tianshanica)种群大树与小树的空间关联性相似[50]。但种子传播不可能无限远,而总是以母树为中心散播其周围,这样离母树稍远距离的种子因受母树抑制作用较弱而能正常生长,因此形成大尺度上的正关联。但天山云杉种群大树与小树在所有尺度均未出现正关联,与本研究结果略有差异,这可能与两种种群的演替阶段和起测径阶不同有关[50]。
本研究中幼树与幼苗、老树与幼苗在所研究空间尺度内并未出现正关联,这可能与该种群的结实特性有关。幼树树龄小,生理上尚未成熟,不具备结实能力,因此与幼苗的关系可能仅限于小尺度的种间竞争,而在大尺度表现为不相关。老树则可能是因为进入老龄期,繁殖能力下降或消失,因此在大尺度上与幼苗也不存在显著的正关联。但老树与幼树在大尺度上呈现一定的正关联,这可能与时间尺度有关。该种群所处环境为高寒环境,加之种内竞争激烈,幼苗生长缓慢[22],因此当现存幼树处于幼苗期时,现存老树可能也处于大树期,还具备一定的繁殖能力,所以在大尺度内与幼树存在一定的正关联。
虽说本研究中小树、中树、大树、老树与幼苗、幼树在不同空间尺度下的关联性变化基本符合Janzen-Connell假说,但总体上仍以负相关和不相关为主要特征,而表现为正关联的大尺度范围较窄,这可能与林下草本、灌木对幼苗、幼树的种间竞争作用有关。色季拉山急尖长苞冷杉林为成过熟原始林,林内林隙发达。林隙因有效改善了林内环境,是冷杉种群自然更新的重要场所,同时也促进了草本与灌木的扩展[51]。由于幼苗、幼树个体较小,其生态位与草本、灌木存在重叠,种间竞争激烈。这种竞争削弱了冷杉种群中大径木个体对小径木个体的促进作用,扩大了抑制作用,最终导致大径木个体与小径木个体在空间分布上表现出以负相关和不相关为主要特征。
