鼓粒期淹水胁迫对大豆叶片AsA-GSH 循环的损伤及烯效唑的缓解效应
2021-07-19王诗雅郑殿峰冯乃杰梁喜龙项洪涛冯胜杰王新欣左官强
王诗雅 ,郑殿峰 ,冯乃杰 ,梁喜龙 ,3,项洪涛 ,冯胜杰 ,王新欣 ,左官强
(1. 黑龙江八一农垦大学农学院,黑龙江大庆163319;2. 广东海洋大学滨海农业学院,广东湛江524088;3. 黑龙江八一农垦大学植物生长调节剂工程技术研究中心,黑龙江大庆163319;4. 黑龙江省农业科学院耕作栽培研究所,黑龙江哈尔滨150086)
大豆(Glycine max)起源于中国,随后于18 世纪传入欧洲和美洲,是世界上最重要的豆科作物之一[1−2]。世界大豆总种植面积为1.2 亿hm2,而中国大豆的种植面积为0.065 亿hm2,约占全球总种植面积的5.4%[3]。黑龙江是中国优质大豆的主产区,种植面积占全国的50.1%[4]。黑龙江地处中国最北部,是典型的“气候脆弱区”,夏季降水率变大,时空分布不均,且大雨、暴雨较为集中,加之黑龙江地势平坦,农田排水困难,较易形成涝灾[5−6],对大豆生长构成严重威胁。
涝渍胁迫是季节性发生的环境灾害,也是作物生长发育过程中遭受的主要非生物胁迫之一,对作物产量和品质的提升造成严重的负面影响[7]。正常条件下,植物体内活性氧(reactive oxygen species,ROS)在抗氧化物酶调节下维持在一个恒定水平,但在逆境胁迫下ROS 的产生和清除机制受到破坏,可导致植物细胞受损[8]。研究指出,作物在胁迫下产生的ROS 和过氧化氢(hydrogen peroxide,H2O2)具有破坏性,可导致酶失活和膜脂过氧化,甚至使细胞死亡[9−10]。抗坏血酸−谷胱甘肽(ascorbate-glutathione,AsA-GSH)循环是植物抗氧化防御系统的重要组成部分,主要由抗氧化物质还原型抗坏血酸(ascorbic acid,AsA)和还原型谷胱甘肽(glutathione,GSH)与抗坏血酸过氧化物酶(aseorbate peroxidase,APX)、谷胱甘肽还原酶(glutathione reductase,GR)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase,MDHAR)及脱氢抗坏血酸还原酶(dehydroascorbatereductase,DHAR)组成[11−12]。研究表明,植物体内 AsA-GSH 循环的快速运转,能减轻 ROS 对植物的伤害[13]。郭欣欣等[14]研究指出,淹水胁迫导致不结球白菜(Brassica rapassp.chinensis)根系中H2O2含量和超氧阴离子产生速率(superoxide anion production rates,O2—·)显著增加,且随胁迫时间延长,对不结球白菜的伤害越大,在淹水胁迫初期通过保持较高的关键酶活性,来抵御胁迫造成的损伤。齐玉军等[15]研究表明,淹水胁迫可使大豆膜脂过氧化程度增加和ROS 积累,使细胞膜受损,AsA 和GSH 作为非酶类可直接清除O2—·,APX、GR 等酶活性的增加可减少 H2O2含量对细胞膜的氧化攻击。因此,AsA-GSH 循环是植物清除ROS 的酶促催化系统,在植物抵抗氧化胁迫、清除逆境ROS 积累方面具有重要作用[16−17]。
烯效唑(uniconazole,S3307)作为一种重要的植物生长延缓剂,可提高作物的抗逆性[18],目前被广泛应用,前人对S3307 在各种作物上的应用效果也进行了大量研究和报道,发现其可有效提高作物的光合作用[19−20],提高酶活性[21],增强作物对逆境的抵抗力[22−24]。S3307 通过提高作物内非酶抗氧化剂和渗透调节物质含量,提高抗氧化酶活性,进而减少ROS 的积累,维持ROS 代谢平衡,进而可缓解逆境对作物造成的伤害[25−26]。目前,淹水胁迫对大豆AsA-GSH 循环的影响研究较少,且关于S3307 缓解大豆淹水伤害的相关研究也尚未报道。因此,本试验以耐涝性不同的2 个大豆品种为材料,探究S3307 对淹水胁迫下大豆叶片ROS 及AsA-GSH 循环中关键酶和抗氧化剂的影响,旨在为S3307 提高大豆的耐涝性提供理论依据。
1 材料与方法
1.1 试验材料
选用耐涝品种‘垦丰14’和涝渍敏感品种‘垦丰16’为大豆试验品种[27]。供试生长调节剂选用烯效唑(S3307),由黑龙江八一农垦大学植物生长调节剂工程技术研究中心提供。
1.2 试验设计与处理
试验于2019 年在黑龙江八一农垦大学国家杂粮工程技术研究中心盆栽场进行,采用带有隔水层和排水口的树脂花盆(上口径×底径×高=32 cm×23 cm×31 cm)进行盆栽试验。试验用土为栽培土、腐殖土和沙子按7∶2∶1 体积比例混合组成,每盆装土18 kg。5 月19 日进行播种,试验共设6 个处理,4 个单元为一个处理,每盆播种10 粒,子叶期定苗5 株,具体设计见表1。待植株生长至R5 期(鼓粒期,播种后95 d),进行叶面喷施S3307(浓度为 50 mg·L−1,喷施量为 225 L·hm−2),并于喷药后第 5 天(播种后 100 d,淹水胁迫处理第 0 天,简称 R5+5)进行淹水处理(套盆淹水,水淹没土表面高于土面2~3 cm 为准),淹水持续5 d(播种后第105 天,淹水第5 天,简称R5+10),5 d 后(播种后110 d,简称R5+15)放水恢复正常水分管理。处理期间,在上述3 个时间点进行取样,每次选取4 株并采集功能叶片(倒三叶),各处理分别取样后立即放入液氮中,而后置于−80 ℃冰箱中保存,供测定生理指标使用。

表1 试验设计方案Table 1 Experiment design
1.3 测定项目及方法
按照高俊凤[28]的方法测定丙二醛(malondialdehyde,MDA)含量、超氧阴离子(O2—·)产生速率、还原型谷胱甘肽(GSH)、氧化型谷胱甘肽(glutathiol,GSSG)含量和抗坏血酸过氧化物酶(APX)活性;采用碘化钾法[29]测定过氧化氢(H2O2)含量;采用 Zhang 等[30]的方法测定还原型抗坏血酸(AsA)和氧化型抗坏血酸(dehydroascorbate,DHA)含量;根据 Zhu 等[31]的方法测定谷胱甘肽还原酶(GR)活性;根据 Murshed 等[32]的方法测定脱氢抗坏血酸还原酶(DHAR)活性。采用试剂盒法(Solarbio 公司)测定单脱氢抗坏血酸还原酶(MDHAR)活性。
1.4 数据处理
采用Microsoft Excel 2013 进行数据录入及整理,用SPSS 25 作方差分析,用Origin 2018 软件绘图。
2 结果与分析
2.1 烯效唑对R5 期淹水胁迫下大豆叶片膜脂过氧化程度的影响
由图 1 可知,(R5+5)d 时,与各自对照相比,S3307 处理有效降低了两大豆品种叶片的MDA 含量,但未达显著差异水平。(R5+10)d 时,淹水胁迫下两大豆品种叶片内MDA 含量显著增加,其中,T2较T1显著增加37.69%,T5较T4显著增加70.09%;而与淹水胁迫相比,S3307 处理显著降低淹水胁迫下两大豆品种叶片内MDA 含量,其中,T3较T2降低4.57%,T6较T5降低24.69%,各处理间均达显著差异水平。恢复正常水分处理5 d(R5+15)后,T2和T5的叶片内MDA 含量虽有降低,但仍高于T3和T6,且二者显著高于两品种的对照(T1和T4),说明叶面喷施S3307 可缓解淹水胁迫对大豆叶片膜脂过氧化的损伤,减轻淹水胁迫造成的伤害。

图1 烯效唑对R5 期淹水胁迫下大豆叶片MDA 含量的影响Fig. 1 Effects of S3307 on MDA content in leaves of soybean under waterlogging stress at R5 stage
2.2 烯效唑对R5 期淹水胁迫下大豆叶片O2—·产生速率和H2O2含量的影响
由图 2 可知,(R5+5)d 时,与对照(T1和 T4)相比,S3307 处理有效抑制垦丰 14 和垦丰 16 叶片内 O2—·产生速率,其中T3较T1降低6.77%,达显著差异水平;T6较T4降低5.77%,未达显著差异水平。(R5+10)d 时,两大豆品种叶片内O2—·产生速率在淹水胁迫下与对照相比显著增加,其中T2较T1显著增加55.75%,T5较T4显著增加32.05%。S3307 处理则有效抑制了两大豆品种叶片内O2—·的产生速率,其中与T2相比T3略有降低,未达显著差异水平;T6较T5显著降低15.81%。恢复正常水分处理[(R5+15)d]后,淹水胁迫处理的垦丰14 T2和垦丰16 T5叶片内O2—·产生速率明显降低,但高于两品种S3307 处理,且均高于对照,其中垦丰14 T3处理恢复至T1水平,说明S3307 处理可抑制叶片内O2—·产生速率,且S3307 处理后,耐涝品种恢复能力强于涝渍敏感品种。

(R5+5)d 时,与对照(T1和 T4)相比,S3307 处理显著降低了 T3和 T6叶片内 H2O2含量,其中 T3较 T1显著降低2.72%;T6较 T4显著降低 1.85%(图 2)。(R5+10)d 时,垦丰 14 和垦丰 16 变化趋势相同,淹水胁迫处理显著增加了两大豆品种叶片内H2O2含量,垦丰14 T2较T1显著增加30.28%,垦丰16 T5较T4显著增加15.74%,S3307 处理可减少H2O2的积累,其中,垦丰14 T3较T2降低3.52%,垦丰16 T6较T5降低2.4%。恢复正常水分处理[(R5+15)d]后,垦丰14 T2和T3与R5+10 d 时相比变化趋势相同,但T2和T3的H2O2含量明显降低,方差分析结果表明T2和 T3均显著高于 T1;垦丰 16 T5和 T6与(R5+10)d 后相比略有降低,T5显著高于 T6和 T4,T6显著高于 T4。说明,叶面喷施S3307 后,耐涝品种垦丰14 在恢复正常水分处理后可有效抑制H2O2含量的积累,减缓胁迫对叶片的伤害;而对于涝渍敏感品种垦丰16 来说,淹水胁迫后叶片恢复能力较慢,淹水胁迫所造的伤害较大。
2.3 烯效唑对R5 期淹水胁迫下大豆叶片AsA 和DHA 含量的影响
由图 3 可知,(R5+5)d 时,垦丰 14 和垦丰 16 在 S3307 处理下(T3和 T6)与各自对照(T1和 T4)相比,显著增加了叶片内 AsA、DHA 和 AsA+DHA 含量,其中 T3较 T1分别显著增加 4.44%、17.25%、3.02%;T6与 T4相比分别显著增加8.65%、2.50%、6.05%。(R5+10)d 时,垦丰14 和垦丰16 各处理之间高低趋势表现为:S3307>淹水胁迫>对照,且各处理间均达显著差异水平,说明淹水胁迫提高了叶片内AsA、DHA 和AsA+DHA 含量,S3307 可进一步促进上述指标的增加。恢复正常水分处理[(R5+15)d]时,两品种淹水胁迫和S3307 处理均有所降低,但S3307 处理仍显著高于淹水胁迫处理,说明S3307 在恢复正常水分胁迫仍可使上述指标保持较高的含量,可有利于ROS 的清除。
2.4 烯效唑对R5 期淹水胁迫下大豆叶片GSH 和GSSG 含量的影响
由图 4 可知,(R5+5)d 时,S3307 处理提高了 T3叶片内 GSH、GSSG 和 GSH+GSSG 含量,分别较 T1增加11.97%、15.35%、14.12%,达显著差异水平;S3307 处理也提高了 T6叶片内 GSSG 和 GSH+GSSG 含量,T6较T4分别显著增加19.16%和9.35%,但对提高GSH 含量效果不明显。(R5+10)d 时,垦丰14 和垦丰16 各处理之间高低趋势分别表现为T3>T2>T1和T6>T5>T4,方差分析结果表明各处理之间差异显著,说明叶面喷施S3307 可提高淹水胁迫下大豆叶片内GSH、GSSG 和GSH+GSSG 含量。恢复正常水分处理[(R5+15)d]后,垦丰14 和垦丰16 淹水胁迫和S3307 处理与(R5+10)d 时相比明显降低,但高低顺序保持不变,说明恢复正常水分处理后,S3307 可维持垦丰14 和垦丰16 叶片内GSH、GSSG 和GSH+GSSG 含量,进而防止淹水胁迫对细胞膜的损伤,提高大豆的耐涝性。

图4 烯效唑对R5 期淹水胁迫下大豆叶片GSH、GSSG 和GSH+GSSG 含量的影响Fig. 4 Effects of S3307 on GSH and GSSG and GSH+GSSG content in leaves of soybean under waterlogging stress at R5 stage
2.5 烯效唑对R5 期淹水胁迫下大豆叶片关键酶活性的影响
2.5.1 烯效唑对R5 期淹水胁迫下大豆叶片APX 活性的影响 由图5 可知,(R5+5)d 时,与各自对照(T1和T4)相比,S3307 处理(T3和 T6)显著提高了垦丰 14 和垦丰 16 叶片内 APX 活性,其中,T3较 T1增加 23.53%,T6较T4增加 26.83%。(R5+10)d 时,两大豆品种淹水胁迫处理(T2和 T5)的叶片内 APX 活性显著增加,S3307 处理(T3和T6)后可进一步促进淹水胁迫下APX 活性的增加。恢复正常水分处理[(R5+15)d]后,垦丰14 和垦丰16 淹水胁迫和S3307 处理APX 活性均有所下降,且高低趋势与(R5+10)d 时相同,但 S3307 处理仍保持较高水平,说明恢复正常水分后,S3307 处理可维持较高的APX 活性,进而有效清除淹水胁迫产生的H2O2。
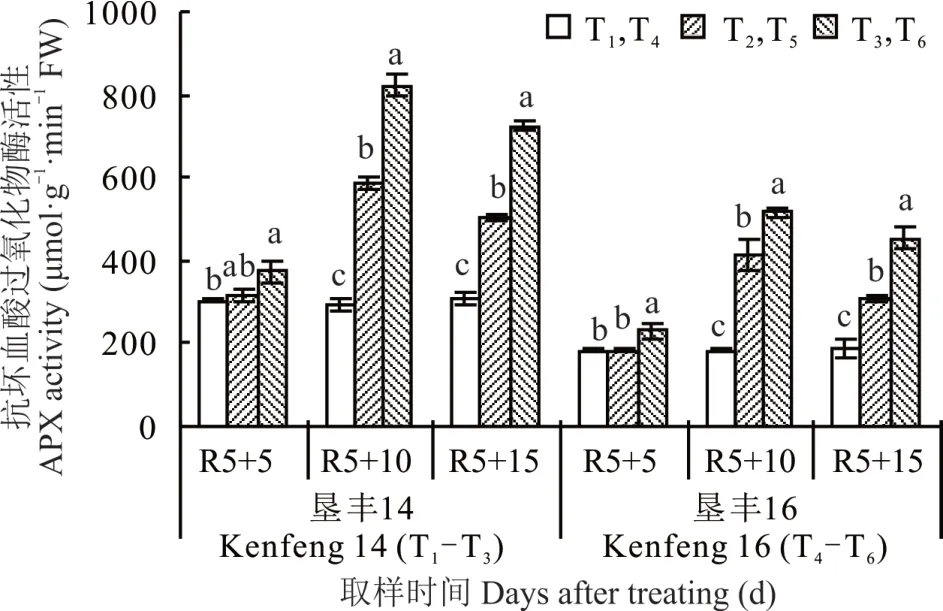
图5 烯效唑对R5 期淹水胁迫下大豆叶片APX 活性的影响Fig. 5 Effects of S3307 on APX activity in leaves of soybean under waterlogging stress at R5 stage
2.5.2 烯效唑对R5 期淹水胁迫下大豆叶片GR 活性的影响 由图 6 可知,(R5+5)d 时,S3307 处理对两大豆品种叶片内GR 活性无明显影响。(R5+10)d时,垦丰14 和垦丰16 各处理之间高低趋势表现为T3>T2>T1和 T6>T5>T4,各处理间均达显著差异水平,说明淹水胁迫可提高叶片内GR 活性,以抵御淹水胁迫造成的伤害,喷施S3307 后,可进一步促进叶片内GR 活性的提高。恢复正常水分处理[(R5+15)d]后,T3和 T2处理与(R5+10)d 时相比显著下降,但趋势相同;垦丰 16 T6和 T5与(R5+10)d 时相比略有降低,但T6仍保持较高水平。S3307 可延缓两大豆品种叶片内GR 活性降低,但恢复正常水分处理后,涝渍敏感品种垦丰16 淹水胁迫处理降幅略小,说明涝渍敏感品种淹水后恢复能力较差。
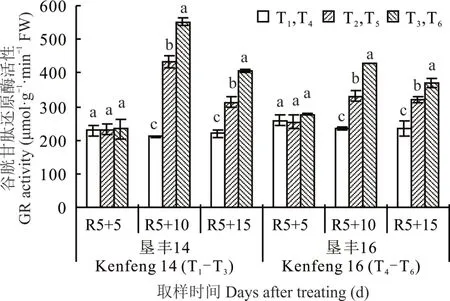
图6 烯效唑对R5 期淹水胁迫下大豆叶片GR 活性的影响Fig. 6 Effects of S3307 on GR activity in leaves of soybean under waterlogging stress at R5 stage
2.5.3 烯效唑对R5 期淹水胁迫下大豆叶片MD⁃HAR 活 性 的 影 响 由 图 7 可 知 ,(R5+5)d 时 ,S3307 处理后T6叶片内MDHAR 活性与T4相比增加24.61%,达显著差异水平;但S3307 处理对垦丰14 叶片内MDHAR 活性的增加效果不明显。(R5+10)d时,显著增加两品种叶片内MDHAR 活性,且叶面喷施烯效唑后可进一步促进MDHAR 活性的提高,其中垦丰14 各处理之间高低趋势表现为T3>T2>T1,垦丰16 各处理之间高低趋势表现为T6>T5>T4,方差分析表明各处理之间差异显著。恢复正常水分处理[(R5+15)d]后,垦丰 14 T3降幅小于 T2,T2恢复至 T1水平;垦丰16 T6降幅小于T5,T5恢复至T4水平。说明恢复正常水分处理后,叶面喷施S3307 具有延缓叶片内MDHAR 活性的作用。
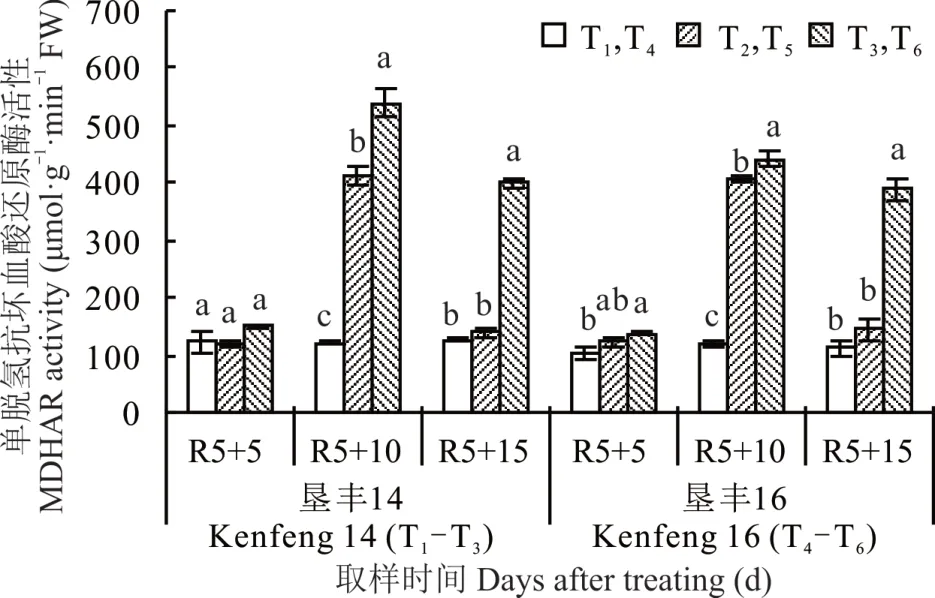
图7 烯效唑对R5 期淹水胁迫下大豆叶片MDHAR 活性的影响Fig. 7 Effects of S3307 on MDHAR activity in leaves of soybean under waterlogging stress at R5 stage
2.5.4 烯效唑对R5 期淹水胁迫下大豆叶片DHAR 活性的影响 由图8 可知,(R5+5)d 时,S3307 处理对垦丰 14 和垦丰 16 叶片内 DHAR 活性无显著影响。(R5+10)d 时,垦丰 14 各处理之间高低趋势表现为 T3>T2>T1,垦丰16 各处理之间高低趋势表现为T6>T5>T4,方差分析表明各处理之间差异显著,淹水胁迫增加了两大豆品种叶片内DHAR 活性,S3307 处理可进一步促进两大豆品种叶片内DHAR 活性的提高。恢复正常水分处理[(R5+15)d]后,垦丰 14 和垦丰 16 淹水胁迫和 S3307处理DHAR 活性均有所下降,但淹水胁迫处理的降幅大于S3307 处理,且各处理间高低趋势与(R5+10)d时相同,说明在恢复正常水分处理后,S3307 处理的两大豆品种叶片内DHAR 活性可保持较高水平,延缓了DHAR 活性的降低,增强清除ROS 的能力。

图8 烯效唑对R5 期淹水胁迫下大豆叶片DHAR 活性的影响Fig. 8 Effects of S3307 on DHAR activity in leaves of soybean under waterlogging stress at R5 stage
3 讨论
3.1 R5 期淹水胁迫及S3307 处理对大豆膜脂过氧化程度的影响
MDA 是膜脂过氧化的主要产物,会对植物细胞产生氧化胁迫并导致膜系统损伤,造成氧化伤害,因此,MDA 含量的高低可直接反应膜脂过氧化的程度,常用作判断胁迫损伤程度的指标[33]。MDA 可与膜上的蛋白质、酶等结合,从而引起蛋白质分子内和分子间的交联,最终破坏膜的完整性以及丧失选择透过性,引起质膜透性的升高,膜系统受到损伤[34]。本试验结果表明,R5期淹水胁迫导致垦丰14 和垦丰16 叶片内MDA 含量大幅增加,加剧了膜脂过氧化程度,且涝渍敏感品种垦丰16受到的胁迫程度大于耐涝品种垦丰14,这与于奇等[24]对绿豆(Vigna radiata)的研究结果一致。研究表明,S3307能够影响植物体内自由基和△-二氢吡咯-5-羧酸合成酶的合成,自由基作用于膜脂过氧化合成,最终产物是MDA,S3307 处理可抑制自由基的合成,并降低MDA 含量,提高作物的耐寒性[22]。本试验结果表明,叶面喷施S3307 后,可抑制淹水胁迫下两品种叶片内MDA 含量的增加,并在恢复正常水分处理后显著降低,与耐涝品种垦丰14 相比涝渍敏感品种垦丰16 恢复较慢,说明外施S3307 可缓解淹水胁迫对大豆叶片膜脂过氧化程度的影响,增强大豆的耐涝性。
3.2 R5 期淹水胁迫及S3307 处理对大豆活性氧代谢的影响
ROS 是植物生长发育中非常重要的角色,正常条件下浓度较低,主要作为信号分子调节植物的生长发育;当受到逆境胁迫时,植物体内的ROS 会急剧积累,而高浓度的ROS 会直接影响细胞的正常发育[35],加快O2—·产生速率和增加H2O2含量,进而破坏膜脂过氧化程度,抑制植物正常生长发育。本试验结果表明,淹水胁迫导致两品种叶片内O2—·产生速率显著增加,这与于奇[36]研究淹水胁迫对绿豆O2—·产生速率影响的结果类似。同时,淹水胁迫还增加了两大豆品种叶片内H2O2含量,与对照相比,分别增加了1.48 和1.43 倍。研究指出,S3307 处理后可降低ROS 过度积累造成的膜损伤,提高作物在逆境胁迫下的耐受性[37]。本试验表明,S3307 可有效降低淹水胁迫下两大豆品种叶片内O2—·产生速率和H2O2含量的积累,说明S3307 对维持ROS 平衡具有积极作用。
3.3 R5 期淹水胁迫及S3307 处理对大豆AsA-GSH 循环的影响
当植物处于淹水胁迫状态时,细胞内正常的ROS 代谢平衡会遭到破坏,进而影响植物的正常生长代谢[38]。前人曾指出,为应对氧化胁迫,植物体内进化出复杂的防御系统,其中,AsA、GSH 含量和AsA-GSH 循环中关键酶活性可起到清 除 ROS 的作用[16]。APX 可 催化 AsA 清除 H2O2生成 MDHAR,且 MDHAR 可协同 APX 清除H2O2;GR 可生成 AsA 进一步降低氧化应激[39−40]。本研究结果表明,淹水胁迫可导致两大豆品种内 APX、GR 和MDHAR 活性升高,说明在淹水胁迫下,H2O2的产生促使两大豆品种叶片APX、GR 和MDHAR 活性的增加,以清除过量的H2O2,该结果与齐玉军等[15]研究淹水胁迫对大豆AsA-GSH 循环中关键酶活性的影响相似。外施S3307 后,可进一步促进上述3 种酶活性的增加,说明S3307 增强了清除H2O2的能力。DHAR 是存在于AsA 和DHA 之间的一种关键酶,在GSH 存在的条件下,催化DHA 向AsA 转变[41]。本研究表明,淹水胁迫下DHAR 含量也呈增加趋势,且S3307 处理后进一步增加了DHAR 活性,说明S3307 可促进DHAR 活性的提高,有助于AsA的产生。恢复正常水分处理后,淹水胁迫解除,抗氧化酶活性降低,但未能恢复至正常生理状态,说明淹水胁迫对叶片的伤害较大;S3307 处理的叶片内关键酶活性和在胁迫解除后维持较高水平,说明S3307 具有延缓关键酶活性降低的作用。AsA 和GSH 是重要的抗氧化剂,可降低O2—·产生速率,并催化 H2O2生成 H2O 和 O2,促进膜蛋白的稳定性,同时,也可作为清除ROS 的调控信号[42−43]。本试验结果表明,淹水胁迫导致叶片内AsA、GSH、DHA、GSSG、AsA+DHA 和GSH+GSSG 含量显著增加,S3307 处理后对上述指标有更明显的促进作用,维持更高的抗氧化能力。胁迫解除后,S3307 可保持较高的抗氧化剂含量,有助于两大豆品种快速恢复至正常生理状态。
4 结论
R5 期,叶面喷施S3307 能有效提高大豆叶片AsA-GSH 循环中非酶抗氧化剂和关键酶活性,维持大豆的正常生长发育;淹水胁迫后导致大豆叶片内ROS 积累和膜质过氧化程度的提高,且涝渍敏感品种垦丰16 的增幅大于耐涝品种垦丰14,同时,促进AsA-GSH 循环中非酶抗氧化剂和关键酶活性的提高,是大豆对淹水胁迫所产生的应激反应,通过两者的协同作用,抵抗ROS 和MDA 对细胞造成的伤害,两品种非酶抗氧化剂和关键酶活性虽有提高,但垦丰16 的幅度低于垦丰14,说明垦丰16 受到的伤害大于垦丰14。叶面喷施50 mg·L−1S3307 具有抵御淹水胁迫,减缓作物受损的作用,增加淹水胁迫下关键酶活性和抗氧化剂含量,有效减少ROS 积累,并维持ROS代谢平衡,缓解淹水胁迫对大豆造成的氧化损伤,在恢复正常水分处理后,通过保持较高的非酶抗氧化剂含量和关键酶活性,促进两大豆品种恢复至正常生长状态。
