甘蓝型油菜钾离子转运载体HAK/KUP/KT家族的全基因组鉴定与分析
2021-07-10朱乐赵鑫泽蒋立希
朱乐,赵鑫泽,蒋立希
(浙江大学农业与生物技术学院作物科学研究所/浙江省作物种质资源重点实验室,杭州 310058)
钾(K)是植物内一种重要的大量营养元素,占植物干物质量的10%。钾参与多种生化过程,包括蛋白质合成、碳水化合物代谢和酶激活,以及许多生理过程,如气孔调节和光合作用[1]。此外,钾还可以增强作物对各种生物和非生物胁迫的抵抗力[2-3]。
植物对K+的需求量很大而植物所处的环境又非常复杂,使得K+转运系统十分庞大,以至于植物从土壤中吸收的K+可以通过不同的组织进行再分配[4-5]。几乎在所有的植物组织中,K+转运系统都是一个大的基因家族,编码多种功能的蛋白质,K+转运载体家族基因通常被分为4 大家族:HAK/KUP/KT、Trk/HKT、KEA和CHX家族。通过与细菌KUP和真菌HAKs 的序列进行相似性比较,在拟南芥中鉴定出第1 个植物HAK/KUP/KT基因[6-7]。目前,HAK/KUP/KT家族数目庞大,对植物的生长发育调节起着重要作用。植物根系对钾的吸收及其在植物体内的转运是由许多K+通道和转运子共同完成的[8-9]。K+通道在外部高K+浓度(>0.5 mmol/L)下起作用,被认为是低亲和转运系统;K+转运体在外部低K+浓度(<0.2 mmol/L)下也可以发挥作用,属于高亲和转运系统;但是,随着K+通道和转运体数目的增加,这2 类系统之间的分界线变得越来越模糊。
植物中存在多种HAK/KUP/KT转运蛋白,它们在K的吸收和转运以及植物生长发育、耐盐性和渗透势的调节中发挥着不同的作用[10]。在拟南芥中,K+转运蛋白AtHAK5 和K+通道蛋白AtAKT1 主要在根细胞中表达,促进根细胞吸收外界环境中的K+[11]。在正常K+浓度条件下,在拟南芥根系中可检测到AtHAK5转录本,而在芽中检测不到;但在K+缺乏的情况下,AtHAK5转录本在根系中的表达水平升高,而且在芽中也能检测到AtHAK5转录本[12]。在水稻中,AtHAK5和AtAKT1的功能是由其同源基因OsAKT5和OsHAK1实现的。研究发现,水稻木质部汁液中的K+浓度以及OsHAK1突变体的K+净输出率明显低于野生型植物,特别是低K+处理(0.1 mmol/L)的植株[13]。K+转运蛋白OsHAK5 可提高K+吸收亲和力,在K+浓度高于OsHAK1 蛋白浓度的情况下,OsHAK5基因在根部维管组织的木质部薄壁组织和韧皮部中强烈表达,在缺K条件下尤为明显,表明OsHAK5可能参与了根与芽之间的K 分配[14]。OsHAK21 具有K+转运活性,但不直接参与K+吸收。OsHAK21主要在中柱的根维管系统中表达,可能在根与芽的长距离K+分配中起作用[15]。
植物体内K+/Na+比值的增大可增强其耐盐性。OsHAK5的过表达增大了植株的K+/Na+比值,从而提高了植株的耐盐性。相反,OsHAK5基因敲除突变体的K+/Na+比值降低,结果转基因植株对盐胁迫的敏感性增加[14]。OsHAK21在盐胁迫反应中也发挥着重要作用。在高盐胁迫条件下,OsHAK21的表达量显著增加,特别是在盐胁迫4 h后,OsHAK21在根中的表达水平增加了600倍。在高盐胁迫下,OsHAK21突变体在地上部和根部积累的K+量均低于野生型(对照)。此外,HAK/KUP/KT转运蛋白也在抗旱性中发挥作用。OsHAK1在水稻营养期和生殖期的过表达均增强了水稻的抗旱性,与对照相比,OsHAK1过表达幼苗的脂质过氧化水平降低,脯氨酸积累量增加,抗氧化酶活性提高[15]。在干旱条件下,OsHAK1过表达植株的产量比野生型植株高出35%[16]。
上述研究表明,HAK/KUP/KT 家族成员对作物的生长发育具有潜在价值,可以提高作物对干旱和盐胁迫的耐受力。同时,随着我国缺K土壤面积日益扩大,科学合理地施用K肥是提高K肥利用效率的有效措施[17-18],而筛选K 高效基因型作物品种是缓解K肥需求压力的另一有效途径。近年来,越来越多的植物HAK/KUP/KT 家族被研究,如梨[19]、甘蔗[20]、辣椒[21]、小麦[22]、大豆[23]和水稻[24]等。油菜是世界上最重要的油料作物之一,其中甘蓝型冬油菜生育期长,对K 素需求量大,但是目前关于甘蓝型油菜(Brassica napusL.)HAK/KUP/KT基因家族系统的鉴定和分析研究未见报道。
本研究利用已经发布的甘蓝型油菜基因组数据,对油菜中HAK/KUP/KT类K转运蛋白家族进行系统鉴定,并对HAK/KUP/KT 家族成员的基因特征、系统发育树、蛋白质结构域、基因结构、染色体位置和顺式作用元件等进行了分析,为深入了解HAK/KUP/KT基因家族的功能及培育K高效油菜品种提供理论依据。
1 材料与方法
1.1 甘蓝型油菜HAK/KUP/KT 家族成员的鉴定与分析
甘蓝型油菜的蛋白质序列来源于已公布的数据库(http://www.genoscope.cns.fr/brassicanapus/);分别从拟南芥的数据库[The Arabidopsis Information Resource(TAIR),https://www.arabidopsis.org/]和水稻的数据库(Rice Genome Annotation Project, http://rice.plantbiology.msu.edu)中下载拟南芥和水稻的蛋白质序列;从Phytozome 数据库(https://phytozome.jgi.doe.gov/pz/portal.html)中下载甘蓝、白菜和大麦的蛋白质序列;随后,以其他物种的HAK/KUP/KT蛋白质序列为查询序列,与甘蓝型油菜的蛋白质序列进行Blastp比对,筛选阈值小于10-10、相似性大于50%的蛋白质序列;再在Pfam 数据库(http://pfam.xfam.org/)中下载HAK/KUP/KT基因家族的隐马尔可夫模型(PF02705)。利用在线分析软件HMMER(https://www.ebi.ac.uk/Tools/hmmer/)和SMART(http://smart.embl-heidelberg.de/)分别对蛋白质序列的保守结构域进行鉴定,最终确定甘蓝型油菜特异的HAK/KUP/KT家族成员。通过ExPASy网站(https://www.expasy.org/)的在线分析软件对蛋白质的分子质量、等电点等特征信息进行分析。
1.2 甘蓝型油菜HAK/KUP/KT 蛋白家族的系统分析和进化树构建
对甘蓝型油菜中HAK/KUP/KT 蛋白家族进行系统进化分析。将从其他物种和甘蓝型油菜中鉴定出来的HAK/KUP/KT 蛋白序列利用Clustal X 进行多序列比对,并将比对结果利用MEGA X软件进行格式转换对齐,然后利用邻接法绘制系统发育树,其中自展值(bootstrap)设置为1 000[25]。
1.3 甘蓝型油菜HAK/KUP/KT 基因的染色体定位与分析
根据甘蓝型油菜HAK/KUP/KT基因序列信息,获得每个基因在油菜染色体上的位置信息,并利用MapChart 软件(https://www.wur.nl/en/show/Mapchart.html)进行染色体定位和作图[26]。
1.4 甘蓝型油菜HAK/KUP/KT 蛋白序列的基序分析
利用MEME 在线工具(http://meme-suite.org/)对甘蓝型油菜的HAK/KUP/KT 蛋白序列进行基序(motif)分析,基序个数设置为10,其他参数为默认数值。
1.5 甘蓝型油菜HAK/KUP/KT 基因结构分析
从甘蓝型油菜基因组数据库中获取BnHAKs基因的编码序列(coding sequence, CDS)和非编码区(untranslated region, UTR)信息,利用GSBS(http://gsds.cbi.pku.edu.cn/index.php)和TBtools(https://github.com/CJ-Chen/TBtools)对基因的内含子和外显子进行可视化展示。
1.6 甘蓝型油菜HAK/KUP/KT 基因顺式作用元件分析
从甘蓝型油菜基因组数据库中提取BnHAKs基因的启动子序列(转录起始位置为上游1 500 bp 序列处)。顺式作用元件的预测通过PLANTCARE在线网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行。
1.7 甘蓝型油菜HAK/KUP/KT基因表达模式分析
从公开的RNA-seq 数据库[27]下载甘蓝型油菜根、茎、叶、萼片、果实和花等6个组织的表达谱数据文件,进行标准均一化处理后,使用MeV 4.9软件绘制基因表达热图。
1.8 实时荧光定量聚合酶链式反应(real‑time quantitative polymerase chain reaction, RT-qPCR)分析
利用快速通用植物RNA提取试剂盒[华越洋生物(北京)科技有限公司]提取样品RNA。第1 链cDNA 利用GoScriptTM反转录系统(GoScriptTMReverse Transcription Mix)、Oligo(dT)[普洛麦格(北京)生物技术有限公司]合成。20 μL RT-qPCR扩增体系:2 μL cDNA 模板,10 μmol/L 上、下游引物各1 μL(引物信息如表1 所示)、10 μL Promega Eastep qPCR 主混合物和6 μL ddH2O。PCR 程序:50 ℃预变性600 s;95 ℃变性180 s;95 ℃变性15 s,60 ℃退火和延伸60 s,共45 个循环;从60 ℃升温到95 ℃,进行熔解曲线分析。每个样品设置3 个生物学重复,以油菜ACTIN7为内参基因,各基因相对表达量使用2-△△CT法计算得出,并且使用软件Excel 2016进行数据统计和分析。

表1 RT-qPCR引物序列Table 1 Primer sequences for RT-qPCR
2 结果与分析
2.1 甘蓝型油菜HAK/KUP/KT 家族成员的鉴定与分析结果
使用Blastp 方法对已报道的拟南芥、水稻与甘蓝型油菜的HAK/KUP/KT蛋白序列进行比对,将所得的序列比对Pfam 数据库,初步确定了45 个编码HAK/KUP/KT蛋白的基因。根据这些基因在染色体上的分布位置,分别将其命名为BnHAK1~BnHAK45(附表1,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2020.08.251)。对鉴定到的45 个BnHAKs 蛋白的理化性质进行分析,发现甘蓝型油菜中BnHAKs蛋白的氨基酸长度在204~1 526个氨基酸残基之间,蛋白质分子质量为22.61~171.55 kDa,等电点范围为4.88~9.63,大部分BnHAKs编码碱性蛋白。
2.2 甘蓝型油菜HAK/KUP/KT 蛋白家族的系统进化分析
为充分研究甘蓝型油菜HAK/KUP/KT基因家族的进化关系,分别利用甘蓝(23个)、白菜(19个)、拟南芥(13 个)、水稻(27 个)和大麦(25 个)的HAK/KUP/KT 蛋白序列与甘蓝型油菜的45 条BnHAKs蛋白序列构建系统发育树。
结果(图1)表明,HAK/KUP/KT 家族分成2 大类,双子叶植物油菜HAK/KUP/KT家族成员与双子叶植物拟南芥、甘蓝和白菜的亲缘关系较近,而与单子叶植物水稻和大麦的亲缘关系较远。在同一亚家族内的蛋白序列保守度较高,而不同亚家族间的差异较大,在同一亚家族内、不同亚组之间也存在一定差异。亚家族Ⅱb的甘蓝和单子叶植物水稻聚集在一起,说明该基因家族出现得较早,随着物种进化,功能比较保守;甘蓝型油菜(B.napusL.,2n=AACC=38)是由甘蓝(B. oleraceaL.,2n=CC=18)和白菜(B. rapaL.,2n=AA=20)通过天然种间杂交而形成的异源四倍体。甘蓝型油菜和甘蓝处于同一姊妹分支的比例高于白菜,表明甘蓝型油菜HAK/KUP/KT 家族成员与甘蓝的亲缘关系更近,HAK/KUP/KT 家族基因可能更多的来自A 染色体,与前人对甘蓝型油菜的全基因组分析结果[28]一致。
2.3 甘蓝型油菜HAK/KUP/KT基因染色体定位结果
从HAK/KUP/KT在甘蓝型油菜基因组上的分布来看,45个BnHAKs基因分布在油菜的1 922条染色体上(图2)。其中:A01号染色体上分布得最多,有5个BnHAKs基因;A03、C04和C07这3条染色体上分布的其次,均有4 个BnHAKs基因;A02、A05、A08、C01、C02、C03、C06、C09、Cnn_random和C01_random这10条染色体上均分布2个BnHAKs基因;其余的8条染色体上均分布1 个。本研究中的45 个甘蓝型油菜HAK/KUP/KT基因没有形成基因簇分布,说明甘蓝型油菜HAK/KUP/KT基因家族无法通过串联重复来实现扩增,因此,HAK/KUP/KT基因家族的扩增可能主要为分散复制和片段复制的形式。
2.4 甘蓝型油菜HAK/KUP/KT 基因家族的结构域分析
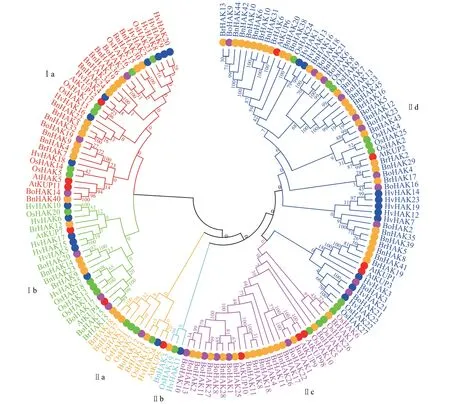
图1 甘蓝型油菜与其他植物HAK/KUP/KT蛋白的系统进化分析Fig.1 Phylogenetic analysis of HAK/KUP/KT proteins in B.napus and other plants
根据系统发育分析,可以将甘蓝型油菜HAK/KUP/KT基因家族分成4个亚家族(图3A)。分析表明:BnHAKs基因序列中至少含有4个外显子和3个内含子,至多含有20 个外显子和19 个内含子。BnHAK10和BnHAK23都只含有4 个外显子和3 个内含子,是所有已鉴定的BnHAKs基因中含外显子和内含子最少的基因;BnHAK42含有20 个外显子和19个内含子,是所有已鉴定的BnHAKs基因中外显子和内含子数量最多的基因。甘蓝型油菜HAK/KUP/KT基因家族含有9和10个外显子的基因各有10 个(22.2%),含有8 和9 个内含子的基因数量最多。在同一个组群中,外显子和内含子的数目也不相同(图3B)。
对每个组群进行详细分析发现,在组群Ⅰ的2个亚组群中,只有BnHAK23和BnHAK19蛋白基序缺少较多;组群Ⅰ的基序与其他3 组基序整齐度一致;组群Ⅱ的4 个亚组群中BnHAK3、BnHAK5、BnHAK10、BnHAK20、BnHAK23、BnHAK38 和BnHAK43蛋白基序缺少较多,特别是Ⅱb亚组群的BnHAK3 蛋白基序只有2 个,保守性较差。甘蓝型油菜BnHAKs 蛋白结构域基序整体保守性较好,只有个别蛋白结构域的基序保守性差(图3C)。
通过MEME 在线工具对甘蓝型油菜BnHAKs蛋白序列进行分析,共鉴定出10 个保守基序(图3D),大多数的保守基序位于K+超级家族结构域的序列中。总的来说,这些基序几乎均匀分布,在所有组群中,基序1、2、3、4、5、6、8、9 和10 都是保守的,只有1个基因因缺少特定基序而例外。
2.5 甘蓝型油菜HAK/KUP/KT 基因家族顺式作用元件分析结果
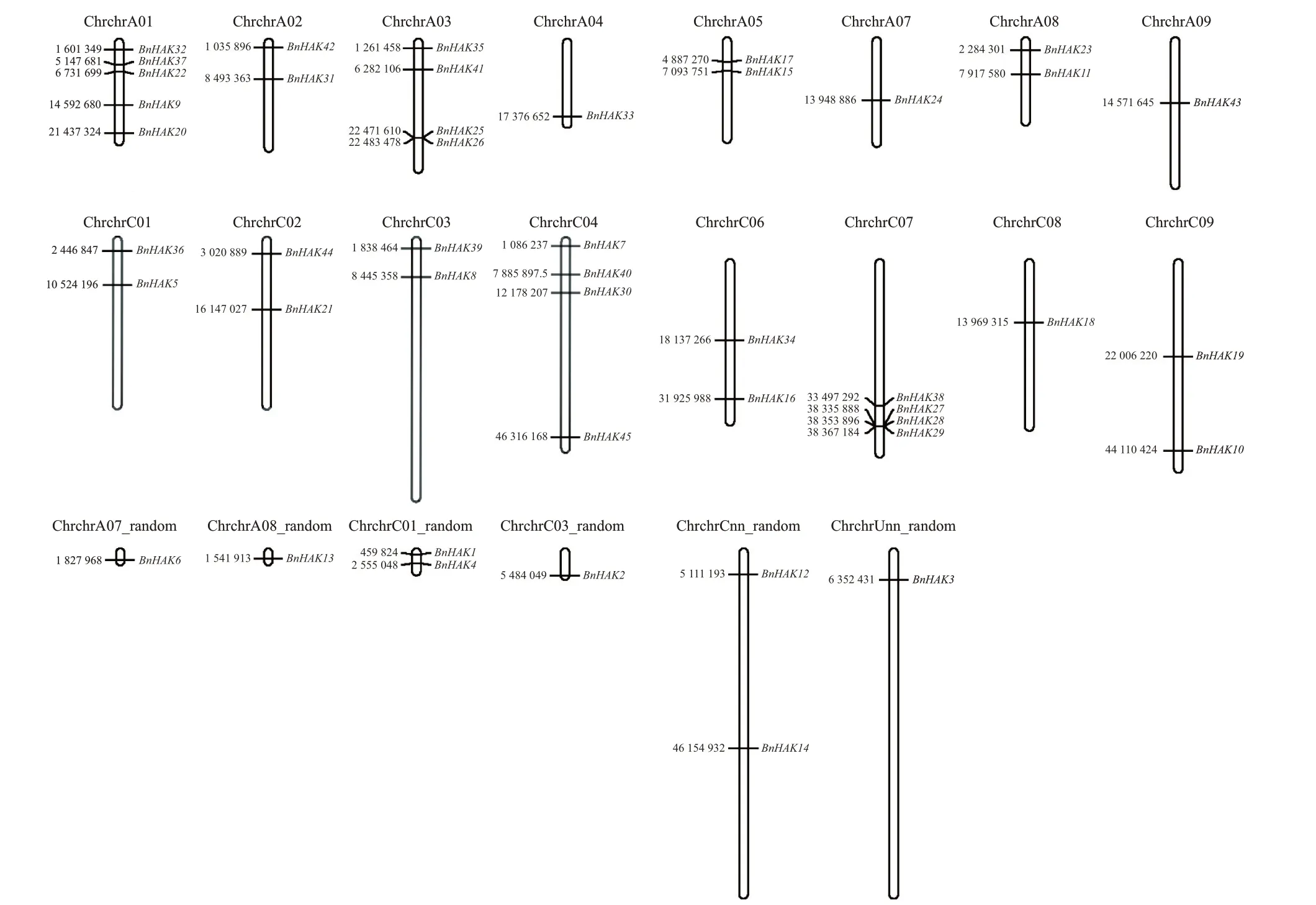
图2 甘蓝型油菜HAK/KUP/KT基因家族的染色体定位Fig.2 Chromosome location of HAK/KUP/KT gene family in B.napus
为了进一步研究HAK/KUP/KT基因家族在非生物胁迫反应中的调控机制,对45 条甘蓝型油菜HAK/KUP/KT家族基因的上游1 500 bp序列进行顺式作用元件分析。结果(图4)显示了11种非生物胁迫响应元件,包括低温应答元件(LTR)、赤霉素应答元件(TATC-box)、防御和胁迫应答元件(TC-rich repeats)、脱落酸应答元件(ABRE)、干旱应答元件(MBS)、茉莉酸甲酯(MeJA)应答元件(CGTCAmotif和TGACG-motif)、水杨酸应答元件(SARE和TCA-element)以及生长素应答元件(AuxRE 和TGA-element)。上述结果表明,多数顺式元件参与激素调控和抗性相关的反应,所以我们推测HAK/KUP/KT基因家族在调控植物的抗性和生长发育的过程中发挥着重要作用。
2.6 甘蓝型油菜HAK/KUP/KT 基因家族在不同组织中的表达分析
利用公开的RNA-seq 数据库预测HAK/KUP/KT基因家族在甘蓝型油菜根、茎、叶、萼片、果实和花中的表达量并绘制热图。在被检测的45 个基因中,42个基因的数据来自甘蓝型油菜已发表的RNA序列数据库,而其他3 个基因(BnHAK20、BnHAK26和BnHAK38)的信息丢失,其在6 个组织中的表达量都设置为0。结果发现,42 个BnHAKs基因的表达模式各不相同,总体而言,它们在根中的表达量最高,在角果中的表达量最低(图5A 和附表1,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2020.08.251)。
在组群Ⅰ的2 个亚组群中,除BnHAK19、BnHAK24和BnHAK34之外,其余基因在6个组织中均有表达;组群Ⅱ的4 个亚组群的大部分基因在不同组织中都有较高的表达水平,其中BnHAK41、BnHAK7和BnHAK8都在花中有较高的表达丰度,而BnHAK14只在根和茎中高表达,在其他组织中的表达量较低。


图3 甘蓝型油菜HAK/KUP/KT基因的系统发育、基因结构和蛋白结构域分析Fig.3 Phylogenetic analysis and structure of HAK/KUP/KT gene and its architecture of conserved protein motifs from B.napus
为了验证基因表达趋势是否正确,利用RTqPCR 分析了油菜根和叶中的基因表达情况,包括BnHAK1、BnHAK4、BnHAK9、BnHAK13、BnHAK18和BnHAK37。以甘蓝型油菜‘中双11’的第3 片真叶的RNA 为模板,基因表达结果如图5B 所示:HAK/KUP/KT基因家族的表达具有组织特异性,其中BnHAK1在叶中的表达量远高于在根中的表达量,而BnHAK37在根中的表达丰度特别高,与转录表达结果一致。
3 讨论
油菜是我国重要的油料作物,2017年油菜的种植面积约665.3万hm2,总产量132.7亿kg,占全国油料作物年产总量的38.2%,是当前我国食用植物油的主要来源。我国油菜种植面积和总产均居世界第一[29],其产量和品质受K 元素的影响较大。K 是植物生存的必需元素,而K+必须从细胞外摄取以满足植物自身的生理需要,因此,植物对K 的吸收及转运也一直是植物生理研究的重点。
本研究对甘蓝型油菜基因组测序,共鉴定出45个HAK/KUP/KT基因,分别命名为BnHAK1~BnHAK45。对甘蓝型油菜和其他物种的HAK/KUP/KT 的系统进化分析发现,HAK/KUP/KT 家族可以分为2 类,其中:第Ⅰ类包含的成员最少;第Ⅱ类成员最多,可分为4 个亚组,所有物种均有涉及。尽管甘蓝型油菜是由甘蓝和白菜通过天然种间杂交而形成的异源四倍体,但是甘蓝型油菜和甘蓝在同一姊妹分支的比例高于白菜,表明甘蓝型油菜HAK/KUP/KT基因家族成员与甘蓝的亲缘关系更近。大多数的BnHAKs旁系同源基因具有相似的基序(motif),表明它们之间的功能具有一定的保守性;也有部分旁系同源基因具有不同的基序,这部分基因可能在进化过程中产生了功能分化。利用MEME分析共鉴定出10个基序,其中大部分被预测为K+转运域的组成部分。激素信号传导途径及逆境胁迫响应调控相关的顺式元件在HAK/KUP/KT基因启动子区域大量存在,暗示着HAK/KUP/KT基因家族在多种激素信号转导途径与逆境胁迫响应的调控过程中可能发挥重要作用。

图4 甘蓝型油菜HAK/KUP/KT基因家族顺势作用元件分析Fig.4 Analysis of the Cis-elements of HAK/KUP/KT gene family in B.napus
表达模式对基因功能的研究具有重要作用,BnHAKs在不同组织中的表达模式具有显著差异。对甘蓝型油菜6个组织中的HAK/KUP/KT基因家族成员分析发现,这45个成员具有不同的时空表达特异性,如BnHAK4、BnHAK9在叶中特异性高表达,显示出二者对叶发育的重要性;BnHAK7 和BnHAK17在花中的表达量最高,表明它们在花发育中具有重要作用;BnHAK11、BnHAK18和BnHAK23在根中特异性高表达,表明它们在根发育中具有重要作用。
BnHAK2、BnHAK5、BnHAK12、BnHAK22、BnHAK32、BnHAK33、BnHAK36、BnHAK39和BnHAK45在大部分组织中的表达量均较高,而BnHAK3、BnHAK19、BnHAK24、BnHAK25、BnHAK29、BnHAK34和BnHAK43在大部分组织中的表达量均较低,这说明不同HAK/KUP/KT基因家族成员的分工是不同的。
4 结论
本研究鉴定出45 个甘蓝型油菜HAK/KUP/KT基因,并对其蛋白质理化性质、染色体定位、基因结构和蛋白功能结构域进行了分析,预测了BnHAKs启动子区域响应多种激素和胁迫的调控元件,检测了HAK/KUP/KT基因在甘蓝型油菜6个组织中的表达模式,为进一步揭示甘蓝型油菜HAK/KUP/KT基因家族的功能机制奠定了基础。
