3个薄壳山核桃品种叶片结构特征和枝条导水功能比较
2021-07-01王兆成周梦钰何的明毕慧慧沈军城傅松玲
王兆成, 王 磊, 周梦钰, 何的明, 毕慧慧, 葛 翔, 沈军城, 傅松玲,①
(1. 安徽农业大学林学与园林学院, 安徽 合肥 230036; 2. 安徽万利生态农业有限公司, 安徽 合肥 231200;3. 阜阳市新丰种业有限公司, 安徽 阜阳 236000; 4. 安徽省顺源农业有限公司, 安徽 合肥 231100)
植物生长过程中水分的消耗量巨大,其中,蒸腾作用散失的水分是光合作用水分用量的数百倍,因此,植物的水分利用效率(WUE)非常低[1-2]。前人的研究发现,高约21 m乔木的最大蒸腾散失水量可达200 L·d-1[3]。气候模型预测21世纪内全球气温将升高1.0 ℃~4.0 ℃,亚热带地区将更加频繁地发生由极端天气引发的干旱[4]。20世纪80年代以来,受人为活动影响,中国气温波动上升,且增温量和增温态势均较世界平均水平更严峻[5]。植物的组织结构特征在干旱少水环境中会发生适应性改变[6]5,可作为植物抗旱性的评价标准[7]。其中叶片的形态和结构可能影响植物的蒸腾作用和抗旱性[8],气孔导度和蒸腾作用强度受气孔的规格和密度影响[9-10]。叶片脉络结构不仅提供机械支撑、保障植物叶片的正常生理机能,而且负责转运水分至叶片各细胞中,气孔特性和叶脉密度能体现植物在长期缺水环境中的抗旱性[11-12]。导管是树木养分输送的主要渠道,导管越粗,导水能力越强[13-14],但导管内径增加,导管间气体透过纹孔渗入管腔而发生栓塞的概率越大,易导致植株缺水,甚至死亡[15-18]。
薄壳山核桃〔Caryaillinoinensis(Wangenh.) K. Koch〕具有深根性的特点,对水分的需求量大。目前,国内对薄壳山核桃的研究主要集中在开花物候期[19-20]、果实发育[21]、果实品质[22]、枝条生长及枝条和叶片碳氮代谢物积累[23]、光合特性[24]、种仁中酚类成分含量和抗氧化能力差异以及优株筛选[25]等方面,而对薄壳山核桃的抗旱性研究较少,仅有少量研究干旱胁迫下不同品种薄壳山核桃幼苗的生长及生理指标[26-28]。薄壳山核桃进入始果期后,若生长旺盛时期和果实灌浆期(8月至9月)水分匮乏将直接限制薄壳山核桃营养生长和果实膨大,导致果实充盈度不足,进而增加了空壳率和坏果率,果实的质量和产量严重受损[29]。目前,有关薄壳山核桃对干旱胁迫响应的研究较少且试材多为幼苗[30],不足以反映丰产林分对干旱胁迫的响应,指导意义有限。此外,年降水量1 000~1 600 mm对薄壳山核桃生长较为理想[31],安徽省薄壳山核桃主栽地区(江淮丘陵及皖北平原地区)年降水量950~1 188 mm,相比较低,夏季伏旱阻碍薄壳山核桃丰产优产。
目前,薄壳山核桃品种‘波尼’(‘Pawnee’)、‘马罕’(‘Mahan’)和‘威斯顿’(‘Western’)在安徽地区的引种栽植面积较大、规模栽培时间较长,其中,‘波尼’和‘威斯顿’原产于美国得克萨斯州(年降水量约700 mm),‘马罕’原产于美国密西西比州(年降水量约1 600 mm)。本研究对进入始果期后这3个薄壳山核桃品种的叶片结构特征和枝条导水功能进行比较和研究,以自然状态下导水损失率为主要依据对3个薄壳山核桃品种进行抗旱性评价,旨在对薄壳山核桃抗旱性品种进行早期诊断,为薄壳山核桃耐旱品种筛选和精准推广提供理论支持。
1 材料和方法
1.1 研究地概况
研究地分别位于安徽省合肥市肥西县、合肥市长丰县以及阜阳市颍泉区,属亚热带季风气候区。3个样地的基本情况和土壤(0~20 cm)理化性质见表1。

表1 薄壳山核桃3个样地的基本情况和土壤理化性质
1.2 材料
于2019年9月(安徽省特大旱情期间且薄壳山核桃处于果实灌浆期),分别在3个样地选择8年生(砧木2年生,嫁接后生长6年)、长势健康的薄壳山核桃品种‘波尼’、‘马罕’和‘威斯顿’母株各3株,每株选取南向、长势基本一致、粗度约6 mm的当年生枝条2根,剪取长度约30 cm的枝条,立即插入盛有纯水的塑料桶,并用修枝剪在桶内水面以下剪去末端2 cm左右的枝条,消除原有栓塞,最后用纯黑塑料袋将整个枝条套住后带回实验室。
1.3 方法
1.3.1 叶片气孔特征和叶脉密度观测 参照王磊[32]18的方法,每根枝条选取1对复叶中部的成熟叶片(小叶)2枚,在叶片背部涂抹指甲油,待指甲油风干后撕下,在Olympus BX53光学显微镜(日本Olympus公司)下观察并用Image J软件测量气孔长度和气孔密度,结果为2个视野的平均值。然后将叶片置于质量体积分数5%NaOH溶液中浸泡至叶肉消失、叶片透明,在每枚叶片的两端及中间部位(避开主脉)选择3个视野在Olympus BX53光学显微镜下拍照,使用Image J软件测量视野内叶脉总长度和叶片面积,计算叶脉密度。
1.3.2 叶片横切面解剖结构观测 参照王磊[32]18的方法,每根枝条选取1对复叶中部的成熟叶片2枚,将叶片剪切并固定、脱水、包埋、染色,使用Leica SM2000R滑走切片机(德国Leica公司)过主脉切厚7 μm的叶片横切片,然后在Olympus BX53光学显微镜下观察和拍照,每个切片对称选取6个视野观测,并用Image J软件测量叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度,然后计算栅栏组织/海绵组织比(栅栏组织厚度与海绵组织厚度的比值)、叶片组织结构紧实度(栅栏组织厚度与叶片厚度的比值)、叶片组织结构松弛度(海绵组织厚度与叶片厚度的比值)。
1.3.3 叶片比叶质量测量 每根枝条选取1对复叶中部的成熟叶片2枚,使用LI-COR 3100C叶面积仪(美国LI-COR公司)测量叶片表面积,然后将该叶片置于60 ℃烘箱烘干72 h后称量叶片干质量,计算比叶质量(叶片表面积与叶片干质量的比值)。
1.3.4 枝条横切面导管特征观测 将枝条剪成长约2 cm的小段,使用Leica SM2000R滑走切片机切厚25 μm的枝条横切片,使用番红-固绿法染色后在Olympus BX53光学显微镜20倍镜下拍照,用Image J软件测量导管直径和导管密度,在40倍镜下拍照后用Image J软件测量导管壁厚(VWT)和导管内径跨度(IDSV),导管壁加固度(VWR)根据公式“VWR=(VWT/IDSV)2”计算。每张照片选取5个视野观测。
1.3.5 自然状态下导水损失率和木材密度计算方法
参照王磊[32]16的方法,取下薄壳山核桃枝条后,依次测定同一枝条的瞬时导水率、最大导水率和边材平均面积,然后计算自然状态下导水损失率和边材比导率。随后将该枝条剪成长2~4 cm的小段,参照王磊[32]16和白重炎等[33]的方法,测量茎段的干质量(m)和鲜体积(V),木材密度(ρ)根据公式“ρ=m/V”计算。
1.4 数据处理与分析
参照王磊[32]17的方法,计算薄壳山核桃不同品种某一抗旱指标的隶属函数值(Xi)。使用EXCEL 2010和SPSS 22.0软件进行数据处理和多重比较(LSD法,P<0.05),采用SigmaPlot 13.0软件绘图。
2 结果和分析
2.1 3个薄壳山核桃品种叶片结构特征的比较
3个薄壳山核桃品种叶片气孔特征和叶脉密度见图1,叶片解剖结构和比叶质量见图2。
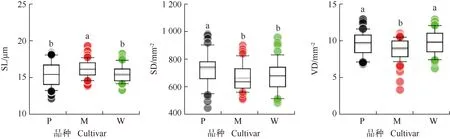
SL: 气孔长度Stomatal length; SD: 气孔密度Stomatal density; VD: 叶脉密度Vein density. P: ‘波尼’‘Pawnee’; M: ‘马罕’‘Mahan’; W: ‘威斯顿’‘Western’. 同一指标不同小写字母表示差异显著(P<0.05)Different lowercases of the same index indicate the significant (P<0.05) difference.
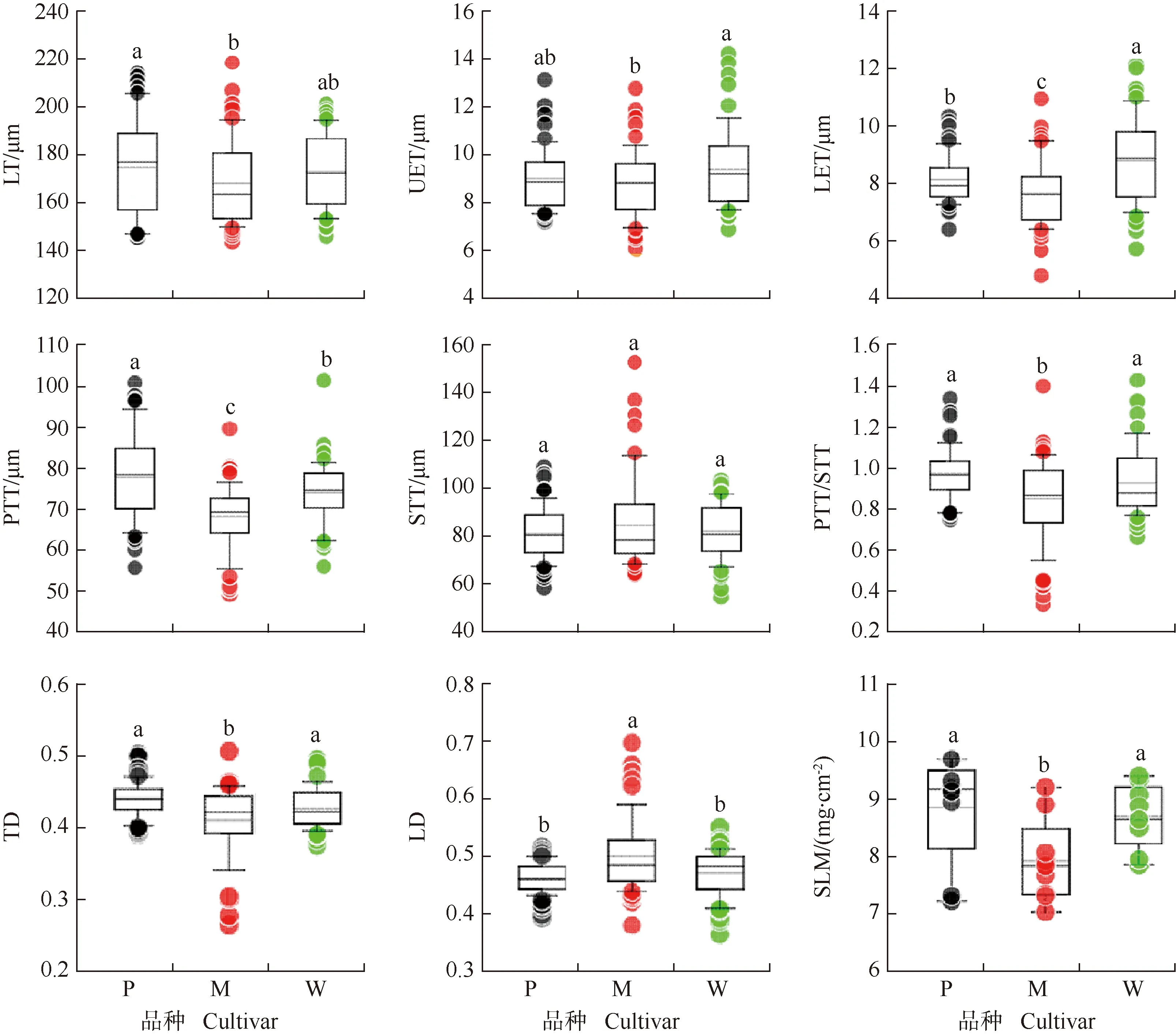
LT: 叶片厚度Leaf thickness; UET: 上表皮厚度Upper epidermis thickness; LET: 下表皮厚度Lower epidermis thickness; PTT: 栅栏组织厚度Palisade tissue thickness; STT: 海绵组织厚度Spongy tissue thickness; PTT/STT: 栅栏组织/海绵组织比Ratio of palisade tissue to spongy tissue; TD: 叶片组织结构紧实度Tightness degree of leaf tissue structure; LD: 叶片组织结构松弛度Loose degree of leaf tissue structure; SLM: 比叶质量Specific leaf mass. P: ‘波尼’‘Pawnee’; M: ‘马罕’‘Mahan’; W: ‘威斯顿’‘Western’. 同一指标不同小写字母表示差异显著(P<0.05)Different lowercases of the same index indicate the significant (P<0.05) difference.
2.1.1 气孔特征和叶脉密度的比较 结果(图1)显示:‘马罕’的气孔长度为15.8 μm,分别较‘波尼’(15.0 μm)和‘威斯顿’(15.2 μm)高5.3%和3.9%,差异显著(P<0.05)。‘波尼’的气孔密度为712.2 mm-2,分别较‘马罕’(648.2 mm-2)和‘威斯顿’(662.2 mm-2)高9.9%和7.6%,差异显著。‘马罕’的叶脉密度为9.0 mm-2,分别较‘波尼’(9.7 mm-2)和‘威斯顿’(9.9 mm-2)低7.2%和9.1%,差异显著。
2.1.2 叶片解剖结构和比叶质量的比较 结果(图2)显示:‘波尼’的叶片最厚(175.4 μm),较‘马罕’(168.4 μm)厚4.2%,差异显著。‘威斯顿’的上表皮(9.3 μm)较‘马罕’(8.7 μm)厚6.9%,差异显著。‘威斯顿’的下表皮(8.8 μm)分别较‘波尼’(8.1 μm)和‘马罕’(7.6 μm)厚8.6%和15.8%,差异显著。‘波尼’的栅栏组织厚度为77.6 μm,显著厚于‘马罕’(68.1 μm)和‘威斯顿’(74.0 μm)。‘波尼’、‘马罕’和‘威斯顿’的海绵组织厚度无显著差异。‘马罕’的栅栏组织/海绵组织比(0.85)显著低于‘波尼’(0.97)和‘威斯顿’(0.93)。‘波尼’和‘威斯顿’的叶片组织结构紧实度分别为0.44和0.43,显著高于‘马罕’(0.41);而‘马罕’的叶片组织结构疏松度为0.50,显著高于‘波尼’(0.46)和‘威斯顿’(0.47)。‘波尼’和‘威斯顿’的比叶质量均为8.9 mg·cm-2,显著高于‘马罕’(7.7 mg·cm-2)。
2.2 3个薄壳山核桃品种枝条导管特征的比较
3个薄壳山核桃品种枝条导管特征见图3。结果显示:‘马罕’枝条的导管直径(45.1 μm)较‘波尼’(42.2 μm)大6.9%,差异显著(P<0.05)。‘波尼’和‘威斯顿’枝条的导管密度均为26.3 mm-2,较‘马罕’(20.0 mm-2)高31.5%,差异显著。‘威斯顿’枝条的导管壁厚为13.2 μm,显著高于‘波尼’(11.9 μm)。‘马罕’枝条的导管内径跨度为56.5 μm,显著高于‘威斯顿’(49.9 μm)和‘波尼’(44.5 μm);‘波尼’和‘威斯顿’枝条的导管壁加固度分别为0.074和0.072,显著高于‘马罕’(0.055)。
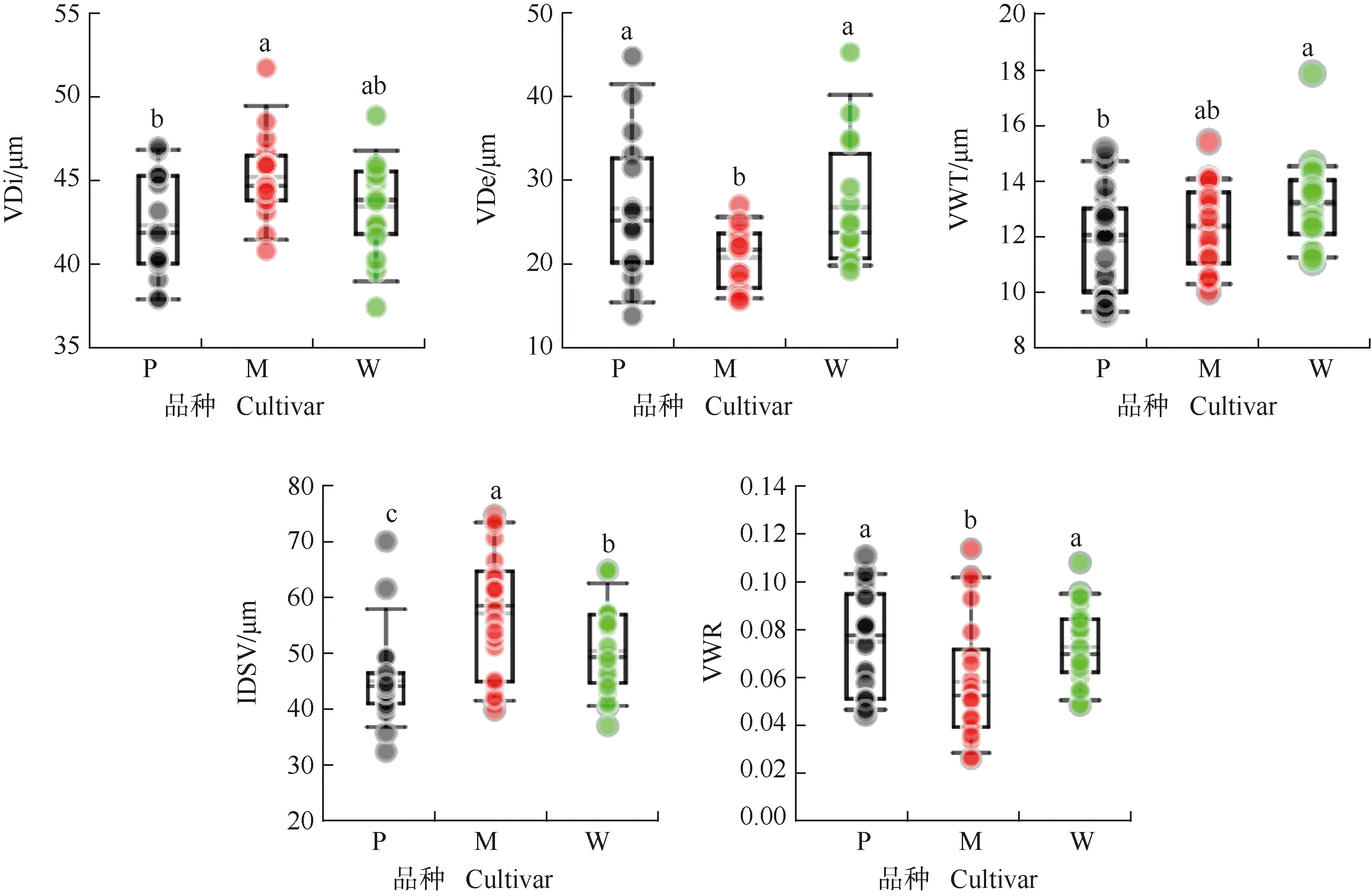
VDi: 导管直径Vessel diameter; VDe: 导管密度Vessel density; VWT: 导管壁厚Vessel wall thickness; IDSV: 导管内径跨度Inside diameter span of vessel; VWR: 导管壁加固度Vessel wall reinforcement. P: ‘波尼’‘Pawnee’; M: ‘马罕’‘Mahan’; W: ‘威斯顿’‘Western’. 同一指标不同小写字母表示差异显著(P<0.05)Different lowercases of the same index indicate the significant (P<0.05) difference.
2.3 薄壳山核桃枝条木材性状和水力性状的比较
3个薄壳山核桃品种枝条的木材性状和水力性状见图4。结果显示:‘波尼’枝条的木材密度(0.49 g·cm-3)高于‘马罕’(0.45 g·cm-3)和‘威斯顿’(0.45 g·cm-3),差异显著(P<0.05)。‘波尼’、‘马罕’和‘威斯顿’枝条的边材比导率无显著差异。‘波尼’和‘威斯顿’枝条的自然状态下导水损失率分别为23.3%和22.2%,显著低于‘马罕’(32.9%)。
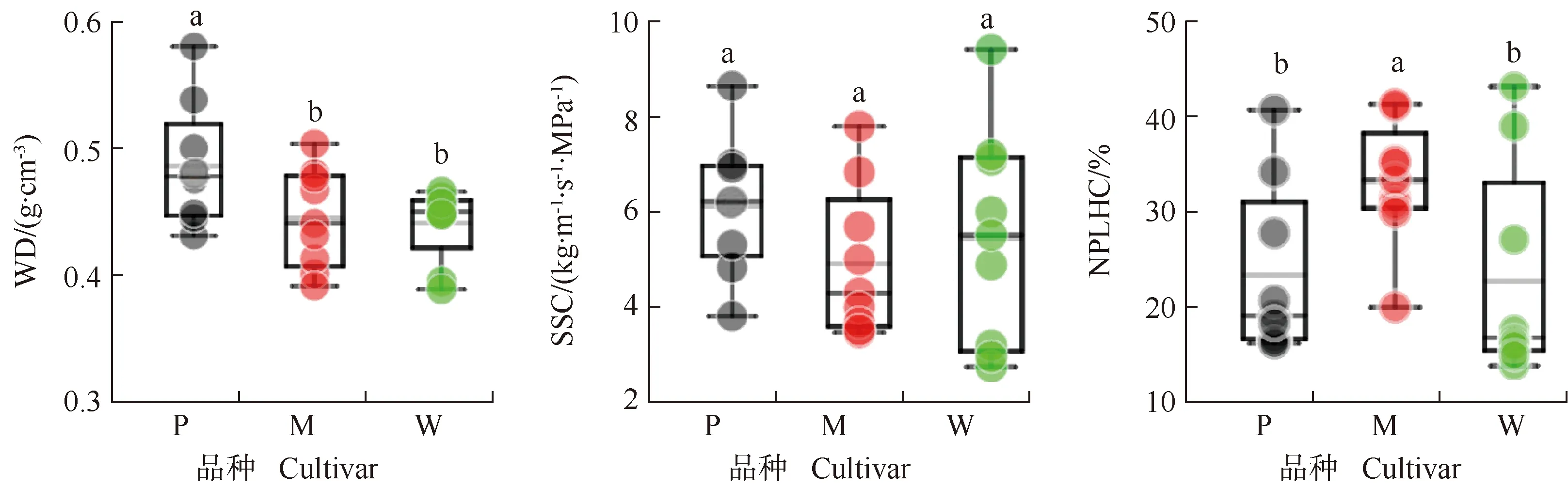
WD: 木材密度Wood density; SSC: 边材比导率Sapwood specific conductivity; NPLHC: 自然状态下导水损失率Natural percentage loss of hydraulic conductivity. P: ‘波尼’‘Pawnee’; M: ‘马罕’‘Mahan’; W: ‘威斯顿’‘Western’. 同一指标不同小写字母表示差异显著(P<0.05)Different lowercases of the same index indicate the significant (P<0.05) difference.
2.4 3个薄壳山核桃品种的隶属函数值
3个薄壳山核桃品种的隶属函数值见表2。结果显示:‘波尼’和‘威斯顿’各指标隶属函数值的均值分别为0.55和0.52,远大于‘马罕’(0.44),说明‘波尼’和‘威斯顿’的抗旱性较强。
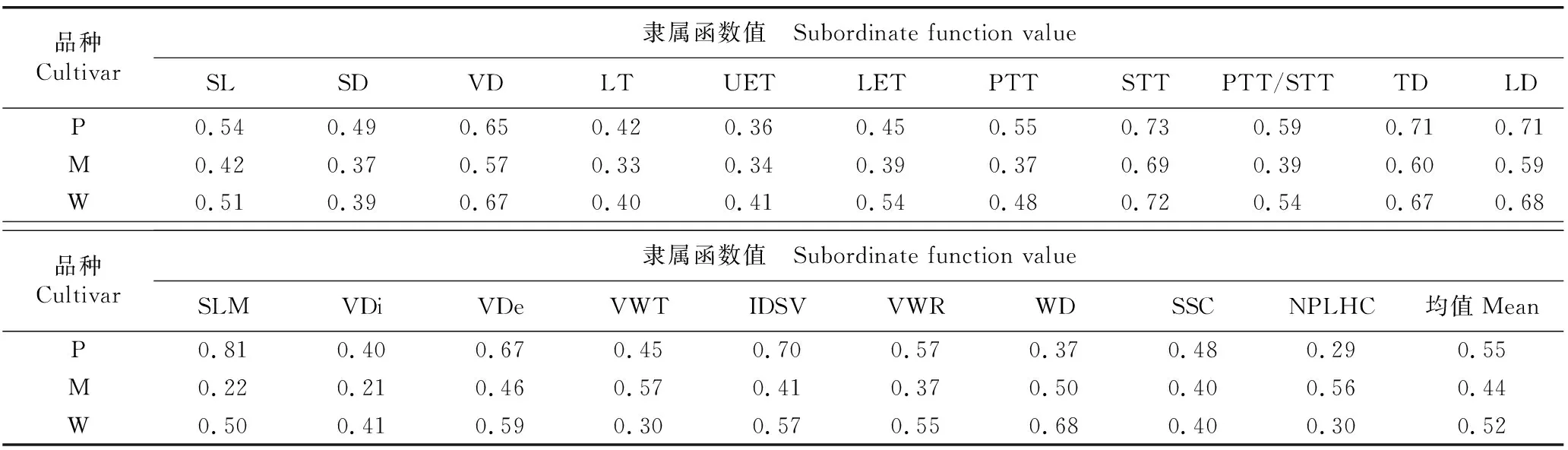
表2 3个薄壳山核桃品种的隶属函数值1)
3 讨论和结论
叶片是光合作用和呼吸作用的主要场所,也是树木表面积最大的器官,对立地条件变化非常敏感[34-35],环境变化和品种差异在叶片形态和结构上均有直观反映,例如:小而密的气孔特征更能适应干旱环境[36]。王树森等[37]认为小而密的气孔特征和更发达的叶脉有利于灌木在干旱环境中生存。本研究中,薄壳山核桃品种‘波尼’和‘威斯顿’拥有小而密的气孔特征和更发达的叶脉,使二者在最大程度保证自身光合作用的前提下能够维持蒸腾作用,当遇到干旱胁迫时,可有效降低叶片水势,达到保护植株的目的。植物叶片对植物体水分供给高度敏感,在长期的进化过程中形成许多特殊结构,并能够通过改变这些结构的形态和数量应对环境水分变化,植物叶片解剖结构可作为评价植物抗旱性的重要依据[34-37]。叶片厚度为体现植物对外界环境适应性的重要表型性状,常用于衡量植物的抗旱性,叶片厚度与叶片贮藏水分的能力呈正相关,通常叶片越厚,抗旱能力越强[38]。除直接测量外,比叶质量能够反映叶片厚度差异。栅栏组织和海绵组织与植物抗旱性密切相关,植物抗旱性随栅栏组织厚度的增加而增强,随海绵组织厚度增加而减弱[39]。白重炎等[40]对12个核桃(JuglansregiaLinn.)品种的研究发现,栅栏组织厚度与植物抗旱性正相关。本研究中,原产于较干旱生境的‘波尼’和‘威斯顿’的栅栏组织厚度显著高于‘马罕’,海绵组织厚度低于‘马罕’,推测‘波尼’和‘威斯顿’叶肉细胞排列紧致,渗透调节能力和水分利用能力更强,更适应较干旱环境。
木质部导管和管胞是高大乔木将水分和营养物质从根系运输到远端叶片的主要通道[41],木质部导水效率与管腔直径的四次方成正比,说明导水效率随导管管腔宽度的增加而提高[42]。栓塞是导管中阻碍水分和养分运输的主要障碍,严重时可导致植株在干旱失水后死亡,栓塞更容易出现在管腔较宽的导管中[6]26。自然状态下导水损失率表明植株在干旱环境中水分运输功能的丧失情况,反映薄壳山核桃在自然失水状态下的栓塞发生情况,可用于比较不同薄壳山核桃品种抗旱性的差异。本研究中,‘波尼’和‘威斯顿’的导管直径明显小于‘马罕’,自然状态下导水损失率也显著低于‘马罕’,与Knipfer等[43]在对原产于干旱和富水生境下核桃幼苗的研究结果类似,导管直径的增加在加快植物输水效率的同时也使植物发生栓塞的概率大大增加,影响植物的抗旱性。
导管间壁的水分通道通过纹孔相互连接,木质部栓塞的产生受纹孔间连接程度和端壁纹孔面积的直接影响,因此纹孔结构在植物抗旱生理研究中尤为重要[43-44]。本研究中,‘波尼’和‘威斯顿’的导管密度较‘马罕’更大,使得‘波尼’和‘威斯顿’在保证导水安全性的情况下兼顾了导水效率。Hacke等[45]研究认为,导管壁加固度与导管发生内爆形成栓塞的风险紧密相关。本研究中,与‘马罕’相比较,‘波尼’和‘威斯顿’导管内径跨度较小,导管壁加固度较大,降低了植物导管中栓塞的发生概率,从而增强‘波尼’和‘威斯顿’导管的安全系数,提高抗旱能力。
综上所述,3个薄壳山核桃抗旱性由强到弱依次为‘波尼’、‘威斯顿’、‘马罕’,在相同的栽培条件下,原产于富水生境下的‘马罕’导水损失率显著高于原产于干旱生境下的‘波尼’和‘威斯顿’。3个薄壳山核桃品种的叶片解剖性状和枝条导水能力均有所不同,这可能是品种长期进化的结果。在实际生产中,应针对立体条件,尤其是降水量和灌溉能力,遵循薄壳山核桃各品种的特性,选择适宜品种,制定合理的供水措施,避免因水分欠缺导致产量及品质下降。
