棉花花青素能量耗散过程维持光合机构稳定性分析
2021-06-07李霞飞李自良贾梦梦陈云瑞张旺锋张亚黎
李霞飞,李自良,贾梦梦,陈云瑞,向 导,张旺锋,张亚黎
(1.石河子大学农学院/新疆生产建设兵团绿洲生态农业重点实验室,新疆石河子 832003;2.新疆乌兰乌苏农业气象试验站/乌兰乌苏国家综合气象观测专项试验外场,新疆乌兰乌苏 832199)
0 引 言
【研究意义】花青素主要通过过滤、衰减和反射高强度的蓝紫光从而调节对光能的吸收[1],陈静等[2]研究低温弱光下光化学能力降低导致过量激发能的积累,而花青素的存在能够维持一定的光合能力。花青素的存在能够减少PSII中的过剩激发能,有效保护PSII的光合活性[3-4];薛占军等[5]研究花青素通过吸收400~600 nm的可见光,有效保护PSI的活性。张开明等[6]研究发现,含有花青素的红叶光合速率较低,主要是由于花青素限制了叶绿体对光能的吸收,影响光反应中心的开放从而影响碳同化过程,花青素与植物光合作用有密切联系[7]。分析含有花青素的棉花光合参数的变化,对研究花青素作为光保护机制的研究有重要意义。【前人研究进展】植物生育进程中常出现各种逆境胁迫,将导致光合器官吸收的光能不能全部用来进行光化学反应,过剩的光能会产生大量活性氧自由基,破坏植物光合机构,影响光合作用的稳定运行[8]。为了防止光损伤的发生,植物激发了各种光保护机制来减少过剩激发能的积累,包括叶片运动[9]、以无害热量形式散失的热耗散[10]、通过光化学途径产生能量物质促进D1蛋白的合成,修复光反应中心[11-12],以及围绕PSI的环式电子传递[13-14]。花青素也可以作为一种光保护机制维持光合机构的稳定运行[15]。依赖叶黄素循环的热耗散[16-18]是保护光合机构免受损伤的重要的光保护机制,类黄酮类物质在长期的光保护过程中比叶黄素循环更为重要[19],但也有研究表明花青素并不具有光保护作用[20]。【本研究切入点】花青素可以作为一种光保护机制保护光合机构免受损伤。有关花青素和其他光保护机制环式电子流和叶黄素循环的热耗散之间关系的研究较少。以不同花青素含量的棉花为材料,研究花青素含量的差异对其光合性能的影响以及为维持光合机构运行所启动的光保护机制。【拟解决的关键问题】分析花青素含量差异与光保护机制的关系,为棉花的高光效育种提供理论依据。
1 材料与方法
1.1 材 料
试验于2018年在新疆中国气象站乌兰乌苏生态与农业气象站(44°17’N,86°03’E)进行。供试材料由国家棉花种质资源中期库(National Medium-term Gene Bank of Cotton in China)和国家棉花种质资源平台(National cotton germplasm resources platform)提供。图1,表1

图1 供试材料

表1 供试材料编号及名称
1.2 方 法
1.2.1 试验设计
小区面积4.5 m2,随机区组排列,重复3次。4月22日布滴灌带,铺膜后人工点播,2 d后进行少量灌溉,保证出苗。宽窄行种植,株距12 cm,密度为22×104株/hm2,其他管理措施同一般大田管理。
待棉花进入盛花期,叶片颜色均匀一致,取主茎上倒数第1片完全展开叶,每个材料取3次重复进行试验。
1.2.2 指标测定
1.2.2.1 叶绿素
将取回的棉花叶片用打孔器(孔径5 mm)打成小圆片4个,放入 5 mL离心管,用约4 mL 80%的丙酮溶液浸提,置于完全遮光处浸提,定时振荡至圆片完全褪绿。取其上清液用分光光度计UV-2041 型分光光度计(日本岛津)比色,叶绿素含量的计算公式参考Lichtenthaler[21]。
1.2.2.2 花青素
选颜色均匀的棉花叶片,在叶脉两侧用5 mm打孔器取15个叶圆片,用15 mL 1%盐酸甲醇溶液提取,在黑暗环境中浸提24~48 h,25 mL容量瓶定容,1%盐酸甲醇溶液为空白对照。在波长530和652 nm处测其吸光度。花青素含量花青素(Ant)=A550-0.25A652[22]。
1.2.2.3 光合参数
Li-6400便携式光合测定系统(Li-COR,USA)测定光合气体参数净光合速率(Pn)。测定时,设置光强 2 000 μmol/(m2·s)、CO2浓度(400±2) μmol/(m2·s)、气体流量300 μmol/s,叶片温度(30±2)℃,参比室的水分(20±2) μmol/(m2·s)。将棉花叶片左上部加入叶室,待数据稳定后,记录数据。
1.2.2.4 吸光系数
用Imaging-PAM荧光成像仪(Walz,德国)测定吸光系数。首先校准仪器,然后将被测材料放置到成像区域,用样品架上的细线固定材料,在材料中上部叶脉两侧选择5~6个区域,测定吸光系数(Abs)。
1.2.2.5 NDH活性
利用Dual-PAM-100叶绿素荧光仪(Walz,德国)测量NDH活性和P700暗还原速率。将取回的棉花材料放置在室温条件下进行0.5~1 h黑暗适应,打开光化光AL(1 828 μmol/(m2·s))进行荧光诱导,待荧光信号趋于平缓状态(大概4 min左右),关掉光化光荧光信号先下降后立即上升,荧光信号立即上升速率能够反映NDH介导的环式电子活性[23]。
1.2.2.6 P515吸收变化
单周转饱和闪光诱导活体棉花叶片的P515快速驰豫动力学可以反映类囊体膜的完整性和ATP合成酶活性[24]。经过1 h暗适应后,P515吸收信号先快速上升,缓慢上升,达到峰值后,缓慢下降。先打开光化光(1 800 μmol/(m2·s))照光10 min,后暗适应4 min,P515吸收信号先快速上升后快速下降,缓慢上升相消失。暗适应后P515吸收信号缓慢下降,膜的完整性,照光后P515吸收信号快速下降的初始斜率可以用来表示ATP酶活性[25]。
利用Dual-PAM-100(Walz,Germany)的新模块P515/535 检测双波长550~515差示吸收的信号变化。将供试材料充分暗适应(至少1 h),测定的515信号能够反映类囊体膜的完整性[24],然后打开光化光(1 828 μmol/(m2·s))诱导棉花叶片10 min,关掉光化光黑暗处理4 min,再测定P515信号吸收变化能够反映ATP合成酶活性[25]。
测定550~515 nm信号的暗-光-暗诱导瞬变,取充分暗适应几个小时的棉花材料,打开光化光AL(1 828 μmol/(m2·s))诱导10 min后515信号趋于稳定(玉米黄质合成的测定),然后关掉光化光AL,515信号先快速下降随后上升逐渐趋于稳定(跨类囊体膜质子梯度的测定)[26-27]。
1.3 数据处理
采用Microsoft Excel 2019进行数据处理,运用Rstudio软件进行方差分析及作图。试验处理间的多重比较采用“Agricolae”程序包进行最小显著差法(Least-significantdifference,LSD)分析,显著水平为0.05,并用英文小写字母标记。图表中的数据均为平均值±标准误。
2 结果与分析
2.1 不同棉花花青素含量的差异
研究表明,绿叶棉花无花青素的存在,而红叶棉花有明显花青素存在,随颜色加深,花青素含量增多。低花青素含量的棉花Chla、Chlb、ChlT均较高花青含量的棉花低,而无花青素的棉花 Chla、Chlb、ChlT和高花青素含量棉花几乎无差异,低花青素棉花类胡萝卜素(Car)较高花青素棉花低,含花青素的棉花较无花青素的棉花低。图2
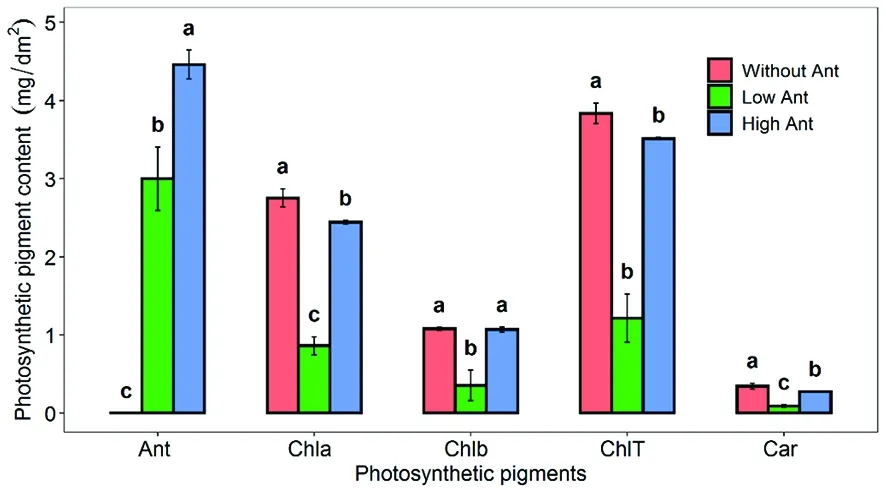
图2 不同棉花花青素含量和叶绿素含量的差异
2.2 不同花青素含量的棉花光合性能的差异
研究表明,含花青素和不含花青素的棉花Fv/Fm均无差异,各棉花可能均相应启动了光保护机制。含花青素的棉花Abs较不含花青素的棉花高,含花青素的棉花吸光系数(Abs)为0.87明显高于较不含花青素的棉花0.83,低花青素棉花Abs 0.86和高花青素棉花0.87无差异;随花青素含量增加,净光合速率(Pn)从31.19到31.43无差异,而无花青素的棉花Pn 34.25显著高于含花青素的棉花31.19。表2
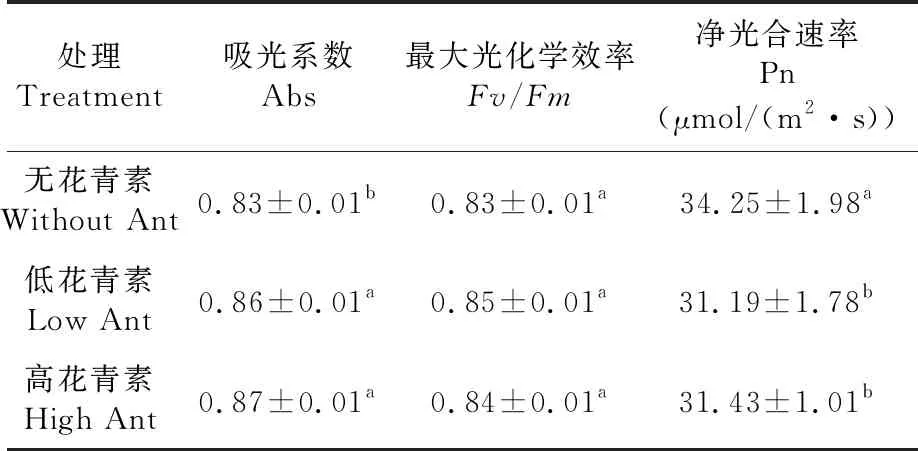
表2 不同花青素含量的棉花气体交换参数变化
2.3 不同花青素含量的棉花NDH介导的环式电子活性的差异
研究表明,随花青素含量增加,荧光信号的快速上升速率显著增强,而花青素含量较低的棉花和无花青素的棉花荧光信号的快速上升无明显差异。花青素含量越高NDH介导的环式电子活性越高,高花青素的棉花伴随较强的NDH介导的环式电子流。图3

A:叶绿素荧光动力学曲线;B:作用光关闭后叶绿素荧光的瞬时上升
2.4 不同花青素含量的棉花ATP合成酶活性的差异
研究表明,在Before AL条件下高花青素含量的棉花和无花青素的棉花515信号缓慢上升几乎无差异,类囊体的完整性都相对较好;而After AL条件下高花青素含量的棉花515信号快速下降的初始斜率显著高于无花青素的棉花。高花青素含量的棉花较无花青素的棉花有较高的ATP合成酶活性,能够合成更多的ATP,而低花青素含量的棉花和无花青素的棉花有相同的趋势。图4
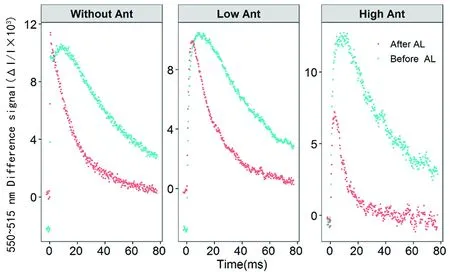
图4 花青素含量的差异对单周转饱和闪光诱导P515的吸收变化
2.5 不同花青素含量的棉花叶黄素循环的热耗散的差异
研究表明,在515信号暗-光诱导下,无花青素的棉花515信号显著较含花青素的棉花强,而低花青素含量的棉花较高花青素含量的棉花强,在含花青素的棉花中玉米黄质的合成速率显著较无花青素的棉花弱,随花青素含量的增多,玉米黄质的合成速率变慢,花青素的存在可能抑制了玉米黄质的合成,并且随花青素含量的增多,抑制作用越强。图5
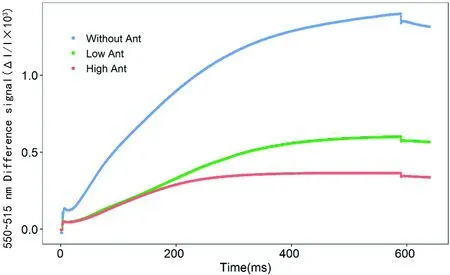
图5 花青素含量的差异下550~515 nm信号的暗-光-暗诱导瞬变的影响诱导瞬变
3 讨 论
3.1 花青素含量增多伴随叶片光合能力的减弱
花青素可通过过滤、衰减和反射高强度的蓝紫光从而来减少光能的吸收[15、27],可以调节叶绿素对光量子的吸收[1]。而光合色素在植物光合作用中主要起对吸收、传递和转化光能的作用,其中Chlb具有捕获和吸收光能的作用,而Chla能够执行能量转换,类胡萝卜素不仅可以作为辅助色素捕获光能,也可以猝灭活性氧防止光合器官受损[28]。高花青素含量的棉花Chla、Chlb、ChlT、Car均低花青素含量的高,吸光系数之间无差异,而相对无花青素的棉花而言,高花青素含量的棉花Chla、Chlb、ChlT较低,吸光系数较高,Car较低。花青素含量越低,吸收、转换光能的能力较弱,猝灭活性氧的能力也较弱,花青素含量的多少能够影响植物对光能的吸收以及猝灭活性氧的能力。
分析净光合速率,低花青素棉花的Pn与较高花青素棉花无差异,相对无花青素的棉花,有花青素的棉花Pn较弱,有花青素的存在并且随着花青素含量逐渐增多,光合性能越弱,可能含有花青素的棉花吸收的光能被花青素吸收、过滤使得转换为光化学的能量减弱,使得光合能力减弱。
3.2 随花青素含量升高NDH介导的环式电子流及其调控能力逐渐增强
环式电子传递是植物体内重要的光合电子传递途径[13、29],研究中,很明显高花青素含量的棉花中NDH介导的环式电子活性显著高于无花青素的棉花,ATP合成酶活性也较无花青素的棉花强;同时随花青素含量的增加,Y(ND)逐渐增强,Y(NA)逐渐减弱,较强的环式电子能够耗散受体侧的电子,减少氧化态的P700,保护PSI免受光损伤,有效保护PSI的活性。进一步分析高花青素含量的棉花ATP合成酶活性较低花青素含量的棉花强,并且高花青素含量的棉花能够较好的保护PSII的活性。随花青素含量的增加,棉花光合能力减弱,而吸光能力增强,将面临较多的过剩激发能,花青素含量的增加伴随环式电子流及其调控能力的增强,有效保护了PSI和PSII的活性。
3.3 花青素能够起到耗散过剩光能保护光合机构的作用
P515信号的暗-光-暗诱导瞬变曲线能够反映玉米黄质的合成速率和类囊体腔内H+的积累程度[26]。515光-暗诱导下,515信号最低点和稳态点之间的差值能够反映ΔpH[27],无花青素的棉花ΔpH明显较含有花青素的棉花高,不含花青素条件下类囊体腔内的H+浓度显著较高。
分布在叶片上表皮的花青素能够阻挡强光[30],Gould等[31]发现含花青素的叶片在受到光损伤后能够快速恢复;Neill和Gould的工作证实,花青素可以耗散过剩激发能减少活性氧的产生[32]。Hughes等[33]研究证实,红叶树种的叶黄素循环过程较弱,更多的依赖花青素的光保护作用。
研究中,含有花青素的棉花玉米黄质的合成速率显著低于无花青素的棉花,可能是紫黄质转化为玉米黄质的过程受到了影响,跨类囊体膜质子梯度(ΔpH)也显著低于无花青素的棉花,也就是说质子梯度驱动的叶黄素循环的热耗散在含有花青素的棉花中较弱,可能含有花青素的棉花主要依赖花青素的光破坏防御功能保护其光合机构免受光抑制。随花青素含量的增加,玉米黄质的合成速率减弱,花青素越多,叶黄素循环的热耗散越弱。相反对于无花青素的棉花来说,主要依赖叶黄素循环的热耗散维持PSII的稳定。
绿叶棉花不含花青素,主要依赖环式电子途径建立的ΔpH驱动叶黄素循环的热耗散过程保护光合机构免受光损伤;而含有花青素的棉花主要依赖环式电子建立的质子梯度驱动ATP合成酶合成ATP,合成D1蛋白修复PSII,减少氧自由基的伤害,另一方面花青素本身具有一定的光破坏防御功能替代了叶黄素循环的过程耗散过剩激发能,保护光合机构。
4 结 论
随花青素含量的增加,净光合速率从34.25减弱至31.43,而吸光系数由0.83增加至0.87,因此,将面临较多的过剩激发能。关关荧信号的快速上升的斜率也逐渐增强,并且After AL条件下515信号快速下降的初始斜率也显著升高,光-暗反应条件下515信号叶黄素循环的热耗散能力逐渐减弱;无花青素的棉花515信号叶黄素循环的热耗散显著较含花青素的棉花强。含花青素的棉花可以通过花青素的耗散过程维持光合机构的稳定,而无花青素的棉花主要通过依赖叶黄素循环的热耗散耗散过剩光能保护光合机构免受损伤。
