龙脊稻作梯田土壤细菌群落结构和功能类群及影响因子分析
2021-05-24王晓彤靳振江周军波程跃扬袁武潘复静刘德深
王晓彤,靳振江,2,3*,周军波,程跃扬,袁武,潘复静,2,3,刘德深,2,3
(1.桂林理工大学环境科学与工程学院,广西桂林 541004;2.桂林理工大学岩溶地区水污染控制与用水安全保障协同创新中心,广西桂林 541004;3.桂林理工大学广西环境污染控制理论与技术重点实验室,广西桂林 541004)
梯田是沿等高线修筑的阶梯状农田,是人为开垦又与独特的自然气候和地理因素密切相关的山地耕作生态系统,其垂直特性是区别于其他农业土地利用方式的显著特征[1]。与普通稻田相比,稻作梯田属于山坡耕地,梯田独特的构造能够增加地表水的下渗量,减缓水流对土壤的冲刷[2],有效防止土壤侵蚀,梯田形式能够有效改善土壤养分状况,改善立地条件。梯田海拔高度的变化会导致土壤养分含量、温度、水分等也随之变化[3-4],并且梯田的光能利用率普遍高于一般农耕地,有助于梯田土壤利用率,发挥梯田效益[5]。我国的稻作梯田主要分布在南方亚热带和热带的丘陵和山地地区。在2018 年第五次全球重要农业文化遗产国际论坛上,包括龙脊稻作梯田在内的4个南方山地稻作梯田被联合国粮农组织列入全球重要农业文化遗产名录[6]。因此,探讨稻作梯田的维持机制有利于农业文化遗产的保护和稻作梯田管理的持续性。
土壤微生物是评价土壤质量和健康状况的重要指标之一,对土壤pH 和养分等环境因子的变化十分敏感[7]。在稻作梯田中,土壤微生物在梯田土壤的生物地球化学循环过程中具有重要的作用[8]。目前,关于梯田土壤微生物方面的研究已有报道,梁勇等[8]的研究发现,元阳梯田稻田土壤微生物数量总体上在低海拔(1 450~1 500 m)稻田较高,土壤自生固氮菌、放线菌、细菌和真菌、纤维分解菌的数量受季节的影响明显。王灿等[9]发现,紫外线辐射能够显著降低水稻孕穗期、抽穗期和成熟期稻田土壤微生物量碳。这些研究表明,土壤微生物易受水稻生长的影响。另外,古梯田水稻土不同年代序列古菌丰度也有报道[10]。而在休闲期,土壤的干扰较少,微生物相对稳定,但对休闲期细菌多样性还鲜有研究;而且,土壤细菌群落结构和功能类群受哪些重要土壤理化因子的影响尚不清楚。因此,选择休闲期的龙脊稻作梯田为研究对象,开展两个方面的研究:一是分析梯田土壤的微生物群落结构和优势功能类群特征;二是明晰梯田土壤中影响优势微生物群落和功能类群的重要环境因子,以期丰富稻作梯田基础理论,为稻作梯田的管理提供科学依据。
1 材料与方法
1.1 研究区概况
龙脊稻作梯田距今已有600 多年的历史[11],堪称世界梯田原乡,位于广西壮族自治区龙胜各族自治县龙脊镇平安村龙脊山,距桂林市约80 km。龙脊稻作梯田地处亚热带季风气候区,年平均气温18.1 ℃,极端最高温度39.5 ℃,极端最低温度-4.8 ℃,无霜期314 d,年降雨量1 500~2 400 mm,年均日照时数1 223 h,年均空气相对湿度82%。龙脊稻作梯田分布在海拔850~1 060 m 之间,位于东经109°32′~110°14′、北纬25°35′~26°17′之间[12]。龙脊稻作梯田高海拔、低气温,主要种植粳稻,为单季稻,产量一般为6 000~7 500 kg·hm-2。通常5 月下旬开始灌水插秧,9 月下旬到10 月上旬成熟,收割后秸秆不还田。施肥以无机复合肥为主,插秧前施用无机复合肥作为基肥,用量为1 050~1 500 kg·hm-2,后期追肥视水稻生长状况而定。
1.2 供试土壤与样品采集
2016 年3 月27 日和3 月28 日,在龙脊梯田景区沿梯田海拔(ALS)高度,大致按照10 m高度差进行采样(图1),共19 个样地,参考《土壤农业化学分析方法》[13]进行土壤样品采集。在每个样地中随机选取3个采样点,每个样点采取等量均匀的0~20 cm 耕层土壤[14],3 个样点均匀混合为1 个样本。样本带回实验室后,去除植物根系和石块等杂物,一部分研磨过2 mm 筛,放入-80 ℃冰箱储存,用于后续细菌群落结构分析;另一部分在阴凉处自然风干后研磨,分别过0.85、0.25、0.15 mm 筛后于阴凉处保存,用于测定土壤理化性质。
1.3 土壤理化指标及养分含量的测定
土壤理化性质的分析方法参照《土壤农业化学分析方法》[13]进行,设样点平行组。土壤含水率(SWC)的测定采用烘干法;pH 的测定采用无CO2蒸馏水作浸提剂,按照土水1∶2.5的比例浸提,用精密pH计(型号:IS128C)直接测定;土壤有效磷(AP)采用盐酸-氟化铵法测定;土壤有机碳(SOC)采用浓硫酸-重铬酸钾外加热法测定;碱解氮(AN)采用碱解扩散法测定;土壤总氮(TN)采用浓硫酸消煮-凯氏定氮法测定;土壤总磷(TP)采用碳酸钠熔融法测定。土壤理化性质见表1。
1.4 高通量测序
用混合样进行微生物测序。使用环境样本DNA提取试剂盒(Power Soil,QIAGEN 公司)进行基因组DNA 抽提后,使用0.8%琼脂糖凝胶电泳检测DNA。按指定测序区域,合成带有Barcode 的特异引物对样本的16S rRNA V4 区域进行扩增[15],引物序列为515F(5′-GTG YCA GCM GCC GCG GTA A-3′)和806R(5′-GGA CTA CHV GGG TWT CTA AT-3′)。每个样本进行3 次重复,每个PCR 反应终止于线性扩增期。PCR 结束后,将同一样本的PCR 产物混合后用2%琼脂糖凝胶电泳检测,使用OMEGA 胶回收试剂盒切胶回收PCR 产物,TE 缓冲液洗脱回收目标DNA 片段。测序由成都罗宁生物公司采用PE250 测序方式在Illumina 测序技术的分析平台上完成。双端测序得到的PE reads首先使用FLASH进行拼接,同时对序列质量进行控制,在去除低质量碱基及接头污染序列等操作过程后完成数据过滤,得到可供后续分析的高质量目标序列。在97%的相似水平上使用UPARSE 算法(默认)进行OTUs(可操作分类单元)的聚类。挑选出OTUs 的代表性序列。使用Uchime 去除嵌合体。使用SILVA 等数据库(默认)或Greengene进行物种分类信息的划分,同时去除注释为叶绿体、线粒体以及非细菌或古菌界的OTUs。α-多样性可以反映细菌群落的丰度和多样性,通过Chao 指数和Ace 指数来分析样品中细菌群落的丰富度,Shannon指数和Simpson指数来分析样品中细菌群落的多样性,Observed 和Coverage指数来表示测序深度。
图1 采样点示意图Figure 1 Schematic diagram of sampling points
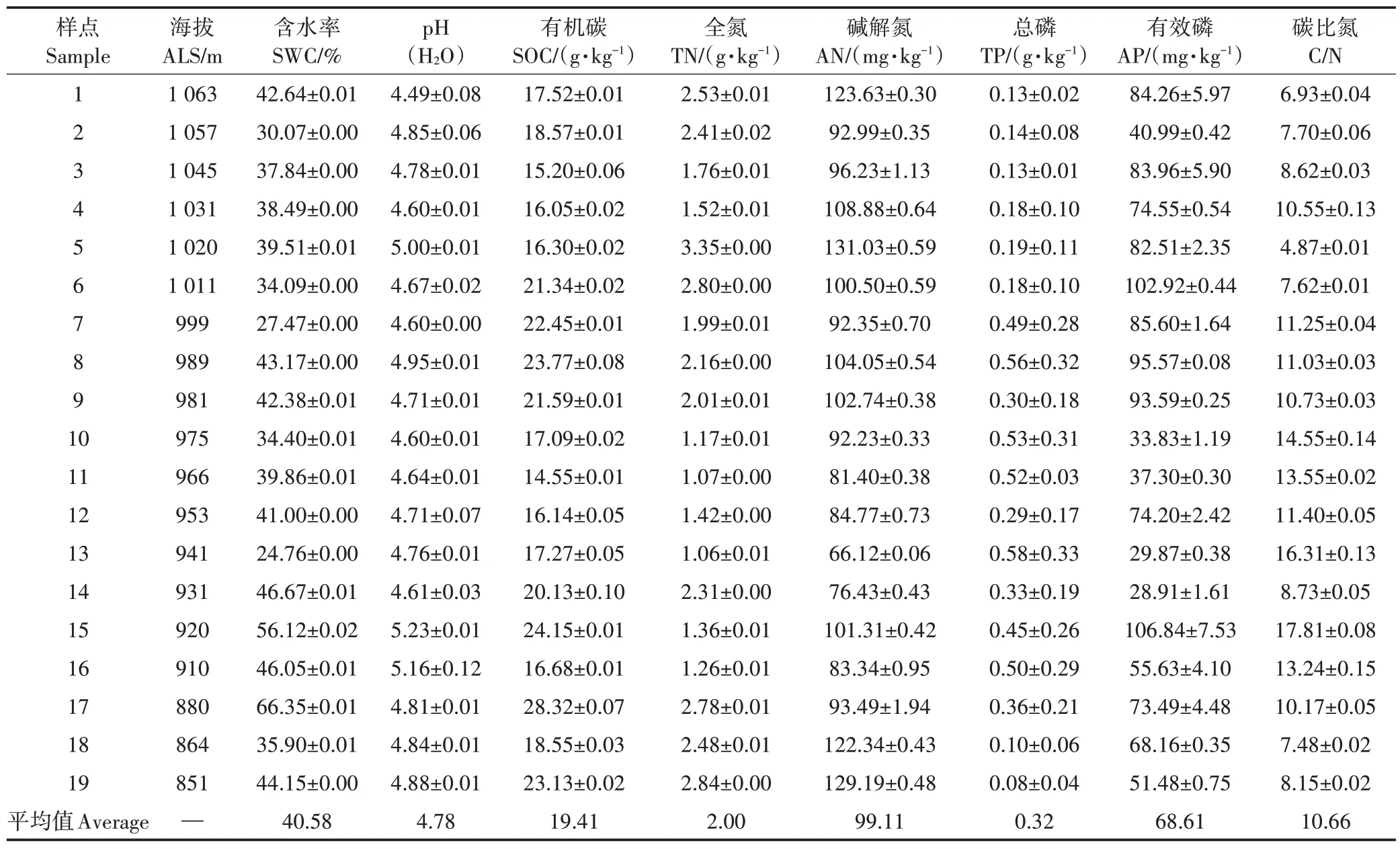
表1 龙脊稻作梯田土壤的理化性质Table 1 Physico-chemical properties of paddy soils in Longji rice terraces(0~20 cm)
1.5 数据分析与统计
采用EXCEL 2010 进行数据处理,使用FAPROTAX在线软件进行细菌功能预测,基于SILVA数据库构建细菌域全谱系的基因功能图谱,进行细菌功能分类预测。用SPSS 25.0 进行Pearson 相关性分析,使用OmicShare Tools 进行主成分分析(PCoA)。用Origin 2017 绘制优势细菌门和属的群落组成图。将数量占总OTUs 数0.3%以上的OTUs 界定为优势OTUs[16],利用Rstudio 和TBtools 绘制优势OTUs 和细菌功能类群与环境因子的相关性热图。
2 结果与分析
2.1 龙脊稻作梯田稻田土壤细菌的多样性分析
本研究中,19 个采样点共得到3 991 个细菌OTUs,它们分属于细菌界的48 门、112 纲、239 目、454科和792 属。从图2 龙脊稻作梯田稻田土壤细菌的α-多样性可知,反映细菌群落丰富度的Chao1指数和Ace 指数分别为1 183~1 984和1 295~2 173;反映细菌群落多样性的Shannon 指数和Simpson 指数分别为4.52~5.16 和0.946~0.976 之间;表示测序深度的Observed指数和Coverage指数分别为819~1 271和0.932~0.961。从表2 土壤环境因子与土壤微生物α-多样性之间的相关性可知,Simpson 指数与ALS 呈显著负相关(P<0.05),而与土壤pH 呈显著正相关(P<0.05),说明土壤细菌多样性受到ALS和土壤pH的影响。
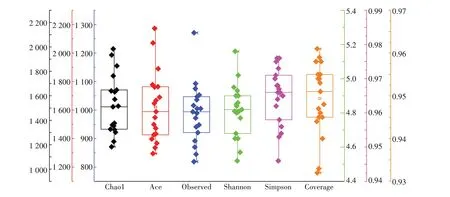
图2 龙脊稻作梯田土壤微生物α-多样性指数Figure 2 Soil microbial α-diversity index in Longji rice terraces
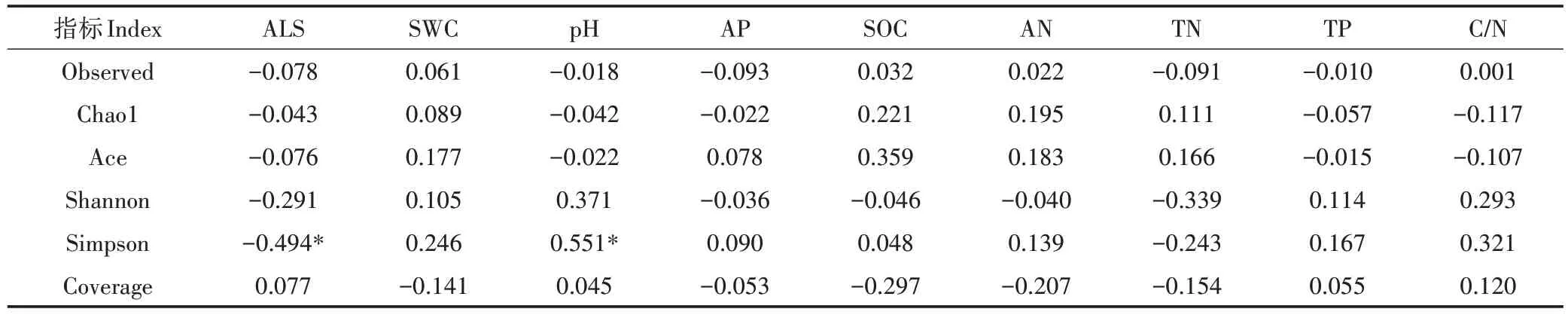
表2 土壤微生物α-多样性与环境因子之间的相关性(r)Table 2 Correlation between soil microbial α-diversity and environmental factors(r)
基于Bray-Curtis 距离算法对不同样点土壤细菌群落组成进行主成分分析(PCoA),结果如图3 所示,轴1和轴2的差异贡献值共为37.91%。从图3可以看出,样点18 和19、样点16 和17、样点13 和14、样点8和9、样点3 和4、样点6 和7 与样点1 和2 两两之间在海拔高度上比较接近,表明细菌β-多样性样点在海拔高度上的分布以小集群分布为主。
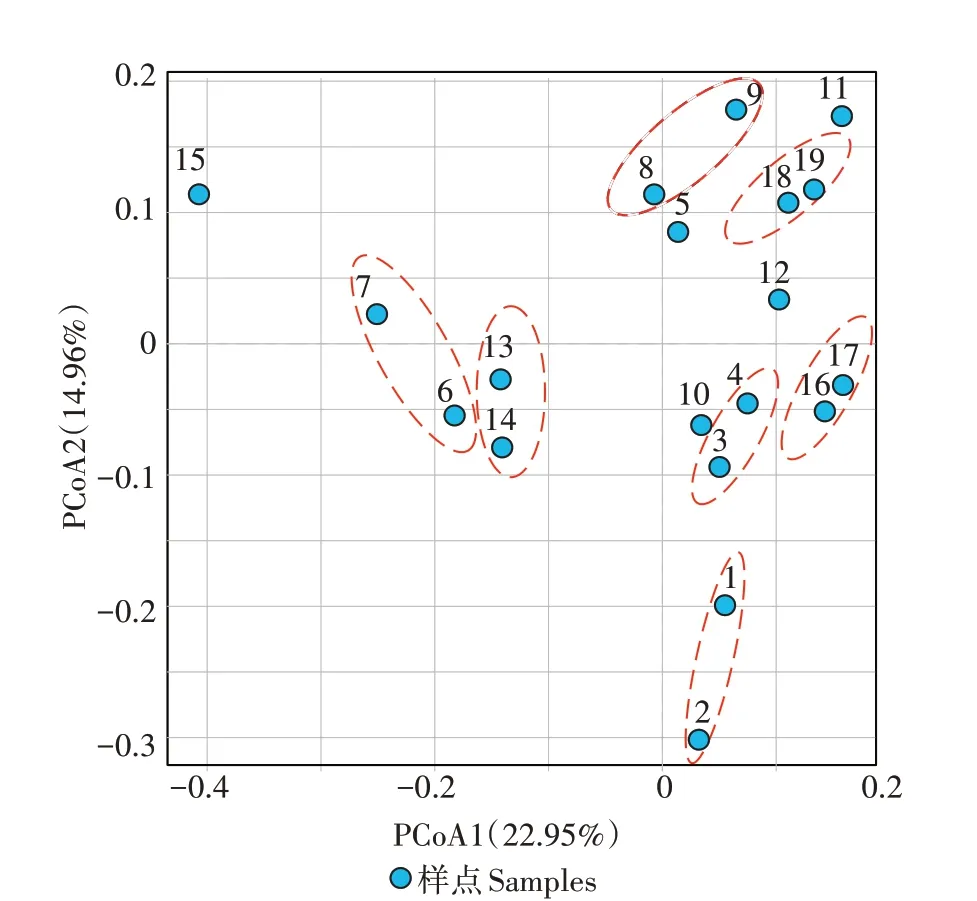
图3 龙脊稻作梯田细菌β-多样性基于Bray-Curtis距离算法的主成分分析Figure 3 Principal component analysis of bacterial β-diversity in Longji rice terraces based on Bray-Curtis distance algorithm
2.2 龙脊稻作梯田土壤细菌群落结构分析
本研究把筛选到门和属水平的OTUs 序列丰度占总OTUs 丰度0.3%以上定义为优势门和优势属,Others 表示未分类的和OTUs 序列丰度占总OTUs 丰度0.3%以下的门和属。土壤细菌在门和属水平的组成及相对丰度如图4所示。
从图4a 可以看出,在门水平上,龙脊稻作梯田共有变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)、WD272 门、芽单胞菌门(Gemmatimonadetes)、拟杆菌门(Bacteroidetes)、浮霉菌门(Planctomycetes)、硝化螺旋菌门(Nitrospirae)和疣微菌门(Verrucomicrobia)等11 个优势门,它们的丰度分别为49.22%、29.18%、5.32%、4.18%、3.76%、2.98%、1.61%、1.06%、0.71%、0.66%和0.40%,其OTUs 丰度合计占细菌总OTUs的99.08%。从图4b可以看出,在属水平上,龙脊稻作梯田共有鞘氨醇单胞菌属(Sphingomonas)、罗思河小杆菌属(Rhodanobacter)、酸杆菌属(Acidibacter)、不动杆菌属(Acinetobacter)和端瘤杆菌属(Telmatobacter)、青枯菌属(Ralstonia)、嗜酸栖热菌属(Acidothermus)、慢生根瘤菌属(Bradyrhizobium)、假双头斧形菌属(Pseudolabrys)、土壤芽孢杆菌属(Solibacillus)、芽孢八叠球菌属(Sporosarcina)、嗜冷芽孢杆菌属(Psychrobacillus)、链球菌属(Streptococcus)、陶厄氏菌属(Thauera)、Rhizomicrobium属和假单胞菌属(Pseudomonas)等16个优势属,OTUs丰度合计占细菌总OTUs 的33.98%。其中样点5、11、16、17、18和19 的最优势属和次优势属分别为罗思河小杆菌属和鞘氨醇单胞菌属,丰度分别为8.72%~21.53%和7.13%~10.27%;其余13 个样点的最优势属和次优势属分别为鞘氨醇单胞菌属和罗思河小杆菌属,丰度分别为8.70%~20.87%和2.34%~11.57%。
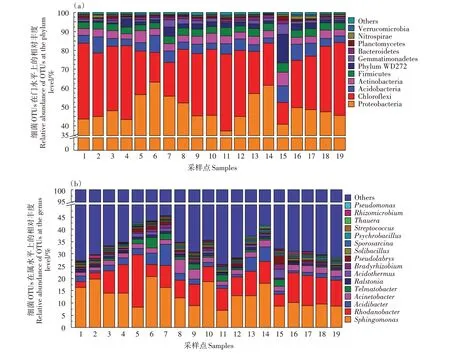
图4 龙脊稻作梯田细菌门(a)和属水平(b)的群落组成Figure 4 Bacterial community composition at the level of phylum(a)and genus(b)in Longji rice terraces
2.3 龙脊稻作梯田细菌群落与环境因子的相关分析
细菌优势OTUs与环境因子的相关性分析结果如图5 所示,pH 与OTU36(Rhodospirillales,红螺菌目)等4 个优势OTUs 呈显著的正相关关系,与OTU1(Sphingomonas,鞘氨醇单胞菌属)和OTU2(Chloroflexi,绿弯菌门)呈显著负相关关系。ALS 与OTU23(Ktedonobacteria,纤线杆菌纲)等3 个优势OTUs 呈显著的正相关关系,与OTU496(Psychrobacillus,嗜冷芽孢杆菌属)和OTU89(红螺菌目)呈显著负相关。
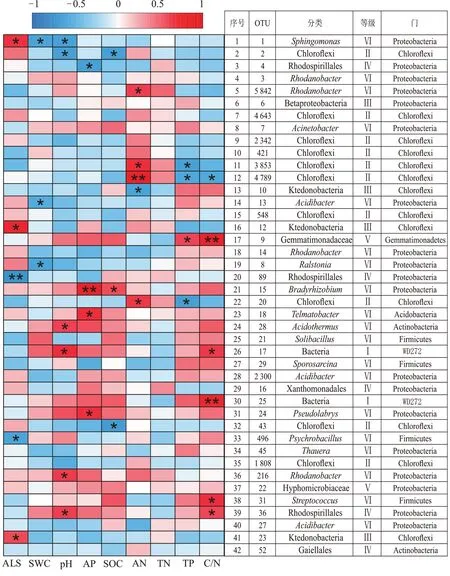
图5 龙脊稻作梯田细菌优势OTUs与环境因子的相关性Figure 5 Correlation between bacterial OTUs for advantage and environmental factors of Longji rice terraces
AN 与OTU20(绿弯菌门)等4 个优势OTUs 呈显著正相关,与OTU10(纤线杆菌纲)呈显著负相关;C/N 与OTU9(Gemmatimonadaceae,芽单胞菌科)等5个优势OTUs 呈显著正相关,与OTU4789(绿弯菌门)呈显著负相关。另外,AP 与3 个优势OTUs 呈显著正相关,SOC 和TP 均与1 个优势OTUs 呈显著正相关。以上结果表明,影响龙脊稻作梯田土壤细菌群落丰度的主要环境因子为pH、C/N、AN和ALS。
2.4 龙脊稻作梯田土壤细菌功能的预测
图6 是龙脊稻作梯田土壤细菌功能菌与土壤理化因子的相关性热图,共获得51 个功能类群,相对丰度占总OTUs 相对丰度的70.36%。相关性分析结果显示,C/N 与发酵类、碳氢化合物降解类、芳族烃降解类和脂肪族非甲烷烃降解类等4 个功能类群丰度呈显著正相关,与好氧氨氧化菌功能类群丰度呈显著负相关;pH 与暗氢氧化类和碳氢化合物降解类、捕食者或外来寄生虫类、芳族烃降解类和脂肪族非甲烷烃降解类等5 个功能类群的丰度呈显著正相关;ALS 与3类硫呼吸类功能细菌呈显著正相关,与胞内寄生类细菌呈显著负相关;AN与反硝化作用类、一氧化二氮反硝化作用类、硝酸盐反硝化类和亚硝酸盐反硝化类等4 个功能类群丰度呈显著负相关(P<0.05)。另外,TN与2个功能类细菌呈显著正相关,含水率、AP和TP分别与1 个功能类细菌呈显著正相关。以上结果表明,影响龙脊稻作梯田土壤细菌群落功能类群的主要环境因子为pH、C/N、ALS和AN。
3 讨论
3.1 龙脊稻作梯田细菌群落结构与土壤因子的关系
土壤微生物的群落组成及多样性是反映土壤微生物群落生态功能的重要指标[16-18]。在本研究中,Simpson 指数与ALS 和pH 分别呈显著负相关和显著正相关,说明海拔的降低和pH 的提高均会增强土壤微生物的物种多样性和均匀度,从而增加土壤生态系统的稳定性。土壤微生物和土壤环境因子都是维持土壤生态系统稳定的重要组成部分[19],微生物的丰富度和数量的增大都会使整个土壤生态系统的稳定性得到明显提升,也使其多样性十分明显[20]。张媚等[21]研究表明,山核桃园土壤pH值降低,土壤微生物的多样性也随之降低,土壤微生物优势菌群的群落结构改变,山核桃树体抗虫抗病能力减弱。相似的结果也出现在茶园中,当茶园土壤pH 低于6.0 时,微生物的生长活性随之减弱[22];反之,施用石灰将土壤pH 从4.1提高至6.88,土壤中细菌和放线菌数量提高近百倍[23],从而改善茶树长势。由此说明土壤微生物多样性与土壤生态系统稳定性之间息息相关。土壤pH会直接影响植物生长和生理代谢,从而影响其对土壤养分的吸收利用,在土壤能量流动和物质循环过程中又反过来影响土壤微生物的多样性[21]。侯建伟等[24]研究发现,随着土壤pH 的升高,细菌群落Simpson 指数也随之增加,说明pH 是决定细菌Simpson 多样性指数的重要因素[25]。在本研究中,受地质背景影响,龙脊稻作梯田土壤pH值呈酸性,梯田开垦时间早,耕作时间长,土壤受人为干扰强烈[8],不同样地之间肥料施用的种类、用量和时间差异较大,影响了细菌群落多样性。从这个意义上说,提高土壤pH 可以提高土壤微生物多样性和均匀度,从而增强龙脊稻作梯田土壤生态系统的稳定性。有研究表明,麦作耕地环境pH 值从4.0 升至8.3 时,相应的细菌拷贝数也随之增加了约3 倍,且细菌群落多样性也增加了1 倍[26-27]。茶园常用石灰调节酸性土壤pH,pH 每升高1~2 个单位,土壤细菌数量会增加10 倍以上[28]。细菌群落生长和pH 值正相关,并且这种现象不会因土壤类型不同而变化[29]。
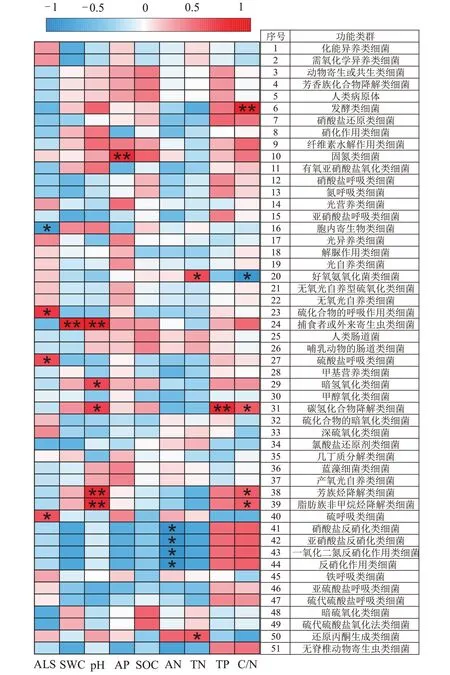
图6 龙脊稻作梯田土壤理化因子与细菌功能类群相关热图Figure 6 Heatmap of soil physicochemical factors with bacterial functional group in Longji rice terraces
在门水平上,变形菌门为最优势菌门。在三江平原湿地[30]、松花江下游沿江湿地[31]、秦岭中段北坡[32]和青藏高原[33]不同生境的土壤中,变形菌门也是细菌群落的优势菌门,表明该门的细菌有较宽的生态位,对土壤环境有高度适应性。理鹏等[34]研究表明,变形菌门细菌的代谢与碳氮循环密切相关,高丰度的变形菌门有利于土壤肥力的保持。在本研究中,β-变形菌、δ-变形菌和α-变形菌均属于变形菌门中的优势纲,β-变形菌可降解烃类芳香族类化合物,参与氮循环。δ-变形菌在化能自养的代谢过程中通过还原硫酸盐升高生境的pH[35]。α-变形菌包含与植物进行共生作用的细菌(如根瘤菌属)[36],表明这个门的细菌群落在水稻吸收利用土壤养分的过程中发挥着重要的作用。图5 相关性分析表明,土壤微生物群落组成和丰度受pH、ALS、AN 和C/N 这些驱动因子的敏感调节。有研究显示,土壤pH 是影响土壤新陈代谢和细菌群落结构和多样性的重要因子[37-39],能显著影响稻田中细菌的多样性和细菌类群的分布[40]。本研究中,pH 与细菌Simpson 指数,门和属水平上的绿弯菌门、Rhizomicrobium属和假双头斧形菌属的丰度以及OTU36(变形菌门)、OTU28(放线菌门)等4 个OTUs 的丰度呈显著正相关(P<0.05),表明pH 显著影响了龙脊稻作梯田土壤细菌群落组成和丰度。龙脊梯田地处温暖湿润的亚热带地区,水分长期在岩石裂隙中移动,对碱土金属有较强的淋滤作用,促使土壤酸化,使pH 降低;同时,淋滤作用促进土壤黏化,改变土壤物理性质,使土壤蓄水保水能力增强,经过长期耕作,形成水稻土[41],为这些细菌创造了适宜生存的稳定条件。另外,海拔高度变化对优势物种具有一定的影响[42],本研究中,ALS 与优势属鞘氨醇单胞菌属、优势OTUs中OTU1(鞘氨醇单胞菌属)等3 个OTUs 的丰度呈显著正相关,说明海拔高度变化影响了龙脊稻作梯田的土壤细菌群落结构;AN 与OTU20(绿弯菌门)等4 个优势OTUs 的丰度呈显著正相关。在本研究中,绿弯菌门为次优势菌门,这个门的细菌在广西的石灰土[16]和太湖地区的黄泥土[43]中均为优势菌门,是进行光合作用的自养微生物,具有固碳作用,因此可使土壤氮转化消耗减少,促进碱解氮含量提升[44]。土壤C/N 是土壤质量的重要反映,在本研究中,C/N 与芽单胞菌门、厚壁菌门、浮霉菌门、链球菌属的细菌和OTU25等5 个OTUs 的丰度呈极显著正相关。研究表明,芽单胞菌主要进行碳氮的固定[45],能够将各种糖分子转化为碳化合物[46],维持土壤中碳含量水平的稳定,同时土壤养分中的碳氮含量也显著影响芽单胞菌的丰度。表明稻作梯田较高的土壤有机碳水平和碳氮养分的相对平衡,为土壤细菌提供了充足的碳氮养分供应,促进微生物的生长,提高土壤微生物多样性[47-48]。
3.2 龙脊稻作梯田细菌功能类群与土壤因子的关系
土壤细菌是土壤微生物中群落关系与功能最复杂多样的生物,土壤环境微小变化可使土壤细菌群落结构和功能发生显著变化,同时这种变化能够反作用于环境[37]。反硝化过程会将NO-3还原成N2,不仅会造成氮损失,还会产生具有温室效应的中间产物N2O[49]。在本研究中,AN 与反硝化功能类群的丰度呈显著负相关,C/N 与反硝化功能类群的丰度呈正相关,说明提高氮素的可利用性可以降低由反硝化造成的氮损失[50],可能是由于细菌将获得的氮素快速合成为新陈代谢所需要的酶,提高自身的生存竞争力。属水平和优势OTUs中的罗思河小杆菌和慢生根瘤菌都可参与反硝化过程,前者能在低pH 时进行反硝化作用,后者在稻田中对N2O 释放有贡献[51],因此可通过适当提高土壤pH 来抑制反硝化功能类群的丰度,减少温室气体的产生。另外,ALS 与硫呼吸类细菌、硫酸盐呼吸类细菌和硫化合物呼吸类细菌功能类群丰度呈显著正相关,这三类功能菌在OTUs 水平上包括了属于硫酸盐还原菌类的脱硫弧菌属、脱硫杆菌属、互营杆菌属和硫还原菌属等。由于微生物在驱动硫化合物的氧化还原过程中也与其他元素(碳、氮和磷等)循环耦合[52],因此海拔可能和土壤碳氮磷等养分共同影响土壤细菌硫循环功能类群的丰度。
烃类物质在土壤中的积累,会改变土壤有机质的组成和结构[53],而农药污染是烃类物质进入土壤的重要来源[54]。在本研究中,环境因子C/N 和pH 均与烃类降解类功能菌显著正相关,表明土壤中C/N 和pH升高能够提高烃类降解功能菌的丰度。微生物降解烃类物质效率高、对环境友好,是修复烃类污染的最佳选择[53]。本研究中,烃类降解功能类群中的优势属鞘氨醇单胞菌属,对芳香族化合物具有极强的降解能力[55-56]。罗思河小杆菌属对难降解的氯化芳族化合物(如林丹)和各种芳族化合物也具有潜在降解能力[57-58]。假单胞菌属对农药、石油烃类都具有良好的降解效果[53]。因此提高土壤有机碳的含量,可以提高C/N,为细菌提供更多的可利用碳源,提高烃类降解功能类群丰度,从而降低农药的残留。龙脊梯田土壤pH 偏酸性,较低的pH 不利于烃类降解功能菌类群丰度的提高,因此,通过改良土壤等方法提高土壤pH可以提高烃类降解功能菌类群的丰度和多样性,加快农药降解速率。
4 结论
(1)龙脊稻作梯田土壤细菌群落的优势门为变形菌门和绿弯菌门,优势属为鞘氨醇单胞菌属和罗思河小杆菌属,土壤pH、碳氮比、海拔和碱解氮是显著影响土壤细菌群落结构的环境因子。
(2)龙脊稻作梯田土壤细菌的功能类群有51 个,受环境因子影响的主要功能类群为反硝化类、硫呼吸类和烃降解类功能类群,主要的环境因子为pH、碳氮比、海拔和碱解氮。
