燕山山脉黄檗种群结构与动态特征
2021-04-25王泳腾黄治昊崔国发
王泳腾,黄治昊,王 俊,张 童,崔国发
北京林业大学生态与自然保护学院,北京 100083
种群生态学的核心是种群动态研究[1-2],它涉及物种种群的动态以及这些种群如何与环境相互作用,是一项关于物种数量随时间和空间变化的研究[3-4]。通过研究生物种群结构,可以评估历史干扰,并对种群未来发展趋势进行预测[5-6]。
黄檗(Phellodendronamurense)为芸香科(Rutaceae)黄檗属(Phellodendron)落叶乔木,是古老的孑遗植物。由于黄檗具有抗炎、降血糖血压等重要的药用价值,在20世纪末受到了严重的人为破坏,野生种群数量日益减少,世界自然保护联盟(International Union for Conservation of Nature,IUCN)将其列为易危(Vulnerable,VU)物种[7]。目前我国黄檗研究主要集中在药用机理分析[8-11]、人工栽培繁育[12]、潜在适宜分布区预测[13]等方面。而对某一局部地区黄檗的种群生态学研究相对较少[14-15],对燕山山脉这一地理尺度上黄檗种群的研究还未见报道。为探究黄檗生态学特性,深入分析黄檗的受威胁状况、濒危程度,阐明黄檗野生种群的濒危机制[16-17],为野生黄檗种群保护和利用提供科学理论依据[18],需要对现存黄檗的种群结构和数量特征开展调查与研究。
1 材料和方法
1.1 研究区概况
燕山位于东经115°—119°47′,北纬39°40′—41°20′,东西长约420km,南北200km左右,东猴顶为其最高峰,海拔2392m。燕山植被类型丰富,既有蒙古栎(Quercusmongolica)、槲栎(Quercusaliena)、栓皮栎(Quercusvariabilis)、槲树(Quercusdentata)、白桦(Betulaplatyphylla)、硕桦(Betulacostata)等落叶阔叶树,又有臭冷杉(Abiesnephrolepis)、华北落叶松(Larixprincipis-rupprechtii)等针叶树种。燕山属暖温带大陆性季风气候区,夏季炎热多雨,冬季寒冷干燥。年均温6—10℃,10℃以上持续期195—205d,活动积温2600—3800℃, 降水季节分配不均匀,全年降水的80%集中在夏季6、7、8三个月,年降水量700mm左右,土地类型以棕壤、褐土为主。
1.2 调查方法
由于黄檗分布稀少且在野外多呈现散生状态,在大量文献调查的基础上,于2019年7—8月,选择了燕山范围内8个国家级自然保护区中黄檗分布较集中且自然林木保存完好的地点作为调查区域,对保护区内几乎所有的黄檗都进行了详细样方调查(图1、表1),设置样地共计33块,每个样地面积为20m×20m,对样地中的乔木进行每木检尺,记录植物的名称、胸径、树高、冠幅、数量等等,同时记录样地的海拔、经纬度、坡度、坡向、坡位、郁闭度等生境条件。

表1 黄檗调查样地的基本概况

图1 自然保护区位置分布图
1.3 数据分析
1.3.1年龄结构划分
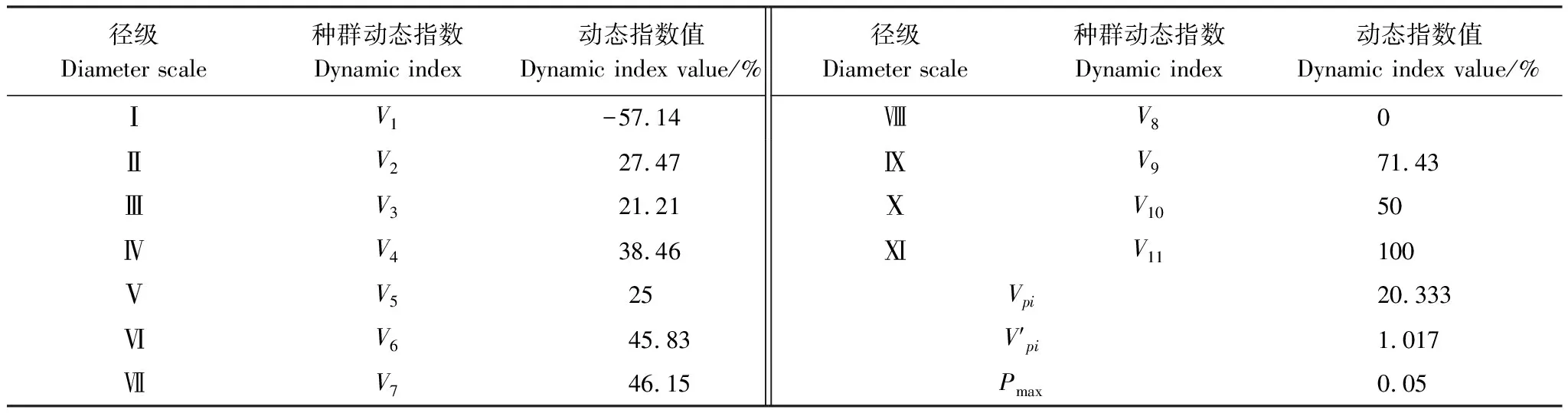
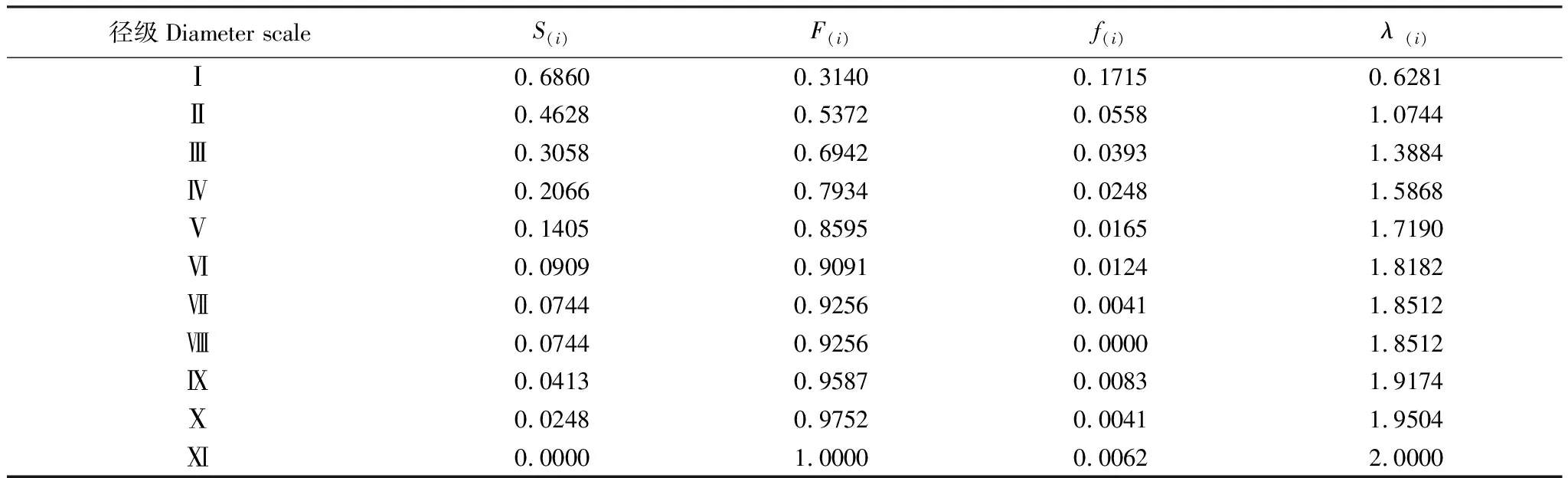
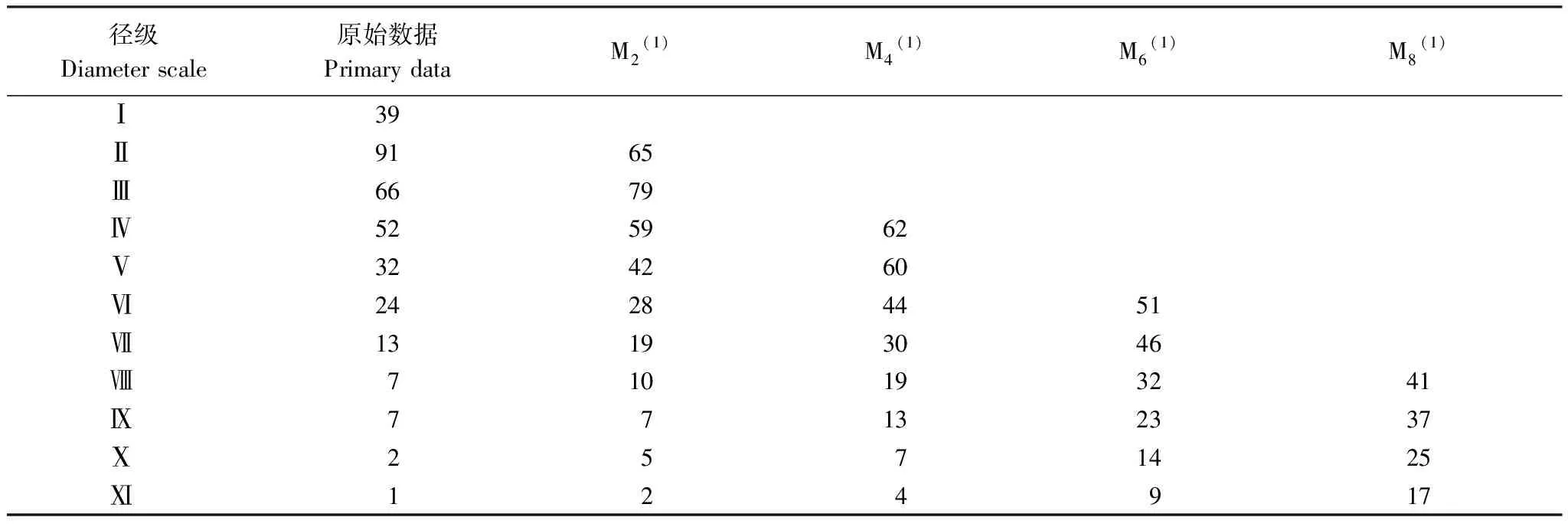
黄檗作为濒危物种,种群数量有限。选择径级结构代替年龄结构,参考相关林木径级划分方法[15][19],根据黄檗的生长规律和生活史特征,将其划分为11个径级:第Ⅰ径级为幼苗(DBH≤4cm);第Ⅱ径级(4cm 1.3.2种群动态数量化方法 参考陈晓德[20]的量化方法,利用动态指数进行分析,具体公式如下: 1.3.3种群静态生命表及存活曲线 参考姜在民、赵阳[21-22]的方法,编制黄檗种群静态生命表,由于静态生命表是根据某一特定时刻种群所有个体的径级编制而成[23],且此次调查中幼苗个体数量较少,会出现死亡率为负的情况,因此对Ax采用方程拟合并做匀滑处理,得到修正后的数据ax,拟合时以径级为自变量,存活数量为因变量,得到的拟合方程为y=182.06e-0.103x,R2=0.8746生命表主要包括以下参数,计算公式分别为: lx=(ax/a0)×1000; dx=lx-lx+1; qx=(dx/lx)×100%; Lx=(lx+lx+1)/2; Tx=Lx+Lx+1+Lx+2…+Lx+n; ex=Tx/lx; Kx=lnlx-lnlx+1; Sx=lx+1/lx; 式中,lx为第x径级的标准化存活个体数;dx为从x到x+1径级间的标准化死亡数;qx为第x到x+1 径级间隔期间死亡率;Lx为第x到x+1径级间隔期间仍存活的个体数;Tx为大于等于x径级的存活个体总数;ex为进入第x径级个体的生命期望或平均期望寿命;Kx为消失率;Sx为存活率。 此外,依据Helt与Loucks的数学模型描述黄檗种群存活曲线,即运用指数方程式Nx=N0e-bx和幂函数Nx=N0x-b分别描述Deevey-Ⅱ型和Deevey-Ⅲ型存活曲线[24]。 式中,Nx、N0分别代x径级内存活数和种群形成初期的个体数,Nx、N0为经匀滑后的个体存活数,b为死亡率。 1.3.4生存分析曲线 +为了更好地分析黄檗种群动态,本文引入4个生存函数,分别是种群生存函数S(i)、累计死亡率函数F(i)、死亡密度函数f(i)和危险率函数λ(i)[25-26],计算公式为: S(i)=S1×S2×S3×…×Si F(i)=1-S(i) f(i)=(S(i-1)-S(i))/hi λ(i)=2(1-S(i))/[hi(1+S(i))] 式中,Si为存活率;hi为径级宽度。 根据上述4个生存函数的估算值,绘制生存率曲线、累计死亡率曲线、死亡密度曲线和危险率曲线。 1.3.5种群数量动态的时间序列预测 采用一次移动平均法[27]预测黄檗种群年龄结构,其计算公式为: 式中,t为径级;n为需要预测的时间;Xk为当前k径级的个体数量;Mt为t径级黄檗在n年之后的种群存活数。本文t分别取2、4、6、8径级,对种群数量进行预测。 本次调查到黄檗共计334株,其中第Ⅱ、Ⅲ、Ⅳ径级个体数量较多,分别占总数量的27.25%,19.76%、15.57%,说明小树数量充足;第Ⅰ径级个体数量占总数量的11.34%,可能是由于21世纪初期,人类砍伐、盗伐现象严重,黄檗种群通过根蘖萌生,由此产生一定数量的幼苗,但由于更新能力不足,导致数量不断减少;第Ⅹ、Ⅺ径级数量也较少,仅占总数量的0.87%,说明黄檗种群老龄和幼龄个体数量匮乏;第Ⅱ—Ⅷ径级的种群数量随径级的增大而减小。黄檗种群径级结构总体上呈倒“J”型(图2),为增长型种群。 图2 黄檗种群径级结构 表2 黄檗种群动态变化指数 从表3和图4可知,黄檗种群存活数量(lx)整体情况为随着径级增大,个体数量逐渐下降,而死亡率(qx)和消失率(Kx)在第Ⅵ径级后波动浮动剧烈(图3),说明种群易受外界环境干扰,种群状态不稳定。生命期望(ex)在第Ⅶ径级达到峰值,说明此时黄檗正值生命力旺盛的壮年时期,此后进入老龄时期,生理活动减弱,生命期望值迅速降低,这种趋势符合生物学特征。 图3 黄檗种群死亡率qx和消失率Kx曲线 表3 黄檗种群静态生命表 以径级(x)为自变量,以标准化存活数(lx)为因变量,绘制黄檗种群存活曲线(图4),并拟合回归得到黄檗种群相应存活曲线模型:Nx=1309.2e-0.353x,R2=0.987,P<0.001。 图4 黄檗种群标准化存活曲线 Nx=1749.3x-1.514(R2=0.915,P<0.001) 结果表明,指数函数模型的R2最大,表明黄檗种群的存活曲线更趋近于各径级死亡率相似的Deevey-Ⅱ型。 黄檗种群累计死亡率F(i)随径级增大而上升,生存率S(i)随径级增大而下降,两者趋势相反。第Ⅰ—Ⅵ径级之间,两者升降变化剧烈,说明黄檗幼龄种群存活率低,自然更新受阻。在第Ⅵ径级之后,两条曲线的变化趋于缓和(图5)。 图5 黄檗种群生存率S(i)和累积死亡率F(i)曲线 黄檗种群危险率函数λ(i)整体呈单调递增,死亡密度函数f(i)在第Ⅰ—Ⅱ径级间急剧下降(图6),在第Ⅷ径级出现最小值(0),这说明黄檗种群开始进入生理衰老期,种群开始衰退。4个生存函数曲线表明,黄檗种群动态表现为前期减少、中期稳定、后期衰退。 表4 黄檗种群的生存函数估算值 图6 黄檗种群死亡密度f(i)和危险率λ(i)曲线 以黄檗种群各径级株数为原始数据,按照一次移动平均法预测各径级在未来2、4、6、8个径级时间后的个体数(表6、图7)。经历2个径级时间后,第Ⅱ径级的数量减少,表明黄檗幼苗数量少,自我更新能力较差;其他径级在2、4、6、8个径级之后的数量均有不同幅度的增加,表明黄檗种群的增长趋势主要依靠中年和老年树木,在外界环境保持良好的前提下,黄檗种群会呈现增长趋势。 表6 黄檗种群数量动态时间序列预测 图7 黄檗种群数量动态的时间序列预测 本研究黄檗种群的存活曲线为Deevey-Ⅱ型,这与张强[28]对辽宁省黄檗种群特征研究中黄檗存活曲线类型一致,但与易雪梅等[8]对长白山地区黄檗种群存活曲线为Deevey-Ⅲ不一致,推测这与长白山地区红松、白桦等针阔叶树的数量居多,且树高冠密,较强地抑制了黄檗幼苗的生长有关。种群死亡率和消失率曲线变化基本一致,都呈现先减少后增加的趋势。黄檗种群前4个径级的累积死亡率增长趋势显著高于后7个径级,这可能是由于黄檗幼龄和低龄个体在生长发育过程中对光照、水分等资源的竞争力不足,说明此时是造成黄檗种群生长波动的重要时期。植物种群的发展不仅涉及到种内种间竞争,种群与环境之间的相互适应过程对其发展也有着重要影响[29]。生存期望表现为先下降后上升再下降的趋势,推测黄檗种群在中龄时对环境的适应和生存能力最强,而进入衰老期,个体对资源不能进行充分利用,数量减少,种群衰退。这与其种群前期减少、中期稳定、后期衰退的结构特征相一致。 黄檗具有一定的萌蘖能力,种子繁殖也是其种群更新的方式之一。燕山地区黄檗种群低龄数量丰富,年龄结构相对完整,短期内不会出现毁灭性的衰退[30]。但是,在本次调查的某些保护区内,仍存在黄檗的盗伐(剥皮)现象,将砍伐后的黄檗树干用来制作枕头治疗颈椎。目前我国采取就地保护、迁地保护、回归自然等措施开展珍稀濒危野生植物保护,鉴于黄檗当前现状,建议重点保护和改善现存黄檗周围环境种群的生境条件,一定程度上限制人类进入黄檗生境区域,从而降低人为破坏;其次,黄檗为阳性树种,对光照要求较高,因此可进行适度的人工修枝,对一些竞争力较强的树种,如胡桃楸(Juglansmandshurica)、大果榆(Ulmusmacrocarpa)进行修枝或者择伐,改善林内光照[31];此外,黄檗幼苗数量不够乐观,应及时清除幼苗周围的杂草、灌木等,为幼苗生长提供充足的空间和资源,提高幼苗的存活率,使更小的植株能够进入林冠层,提高中、成年树的比例。 本文从燕山这一大区域来对黄檗进行了整体研究和分析,但由于每个保护区存在生境异质性,不同保护区内黄檗所处的群落也未尽相同,而海拔、坡度、坡向等也是影响黄檗群落动态的主要因素,下一步将开展影响黄檗种群生长的微生境因子研究,探讨其濒危机制和致危因素,为黄檗种群的快速恢复和保育提供重要的理论依据。
2 结果与分析
2.1 黄檗种群分布现状及结构特征
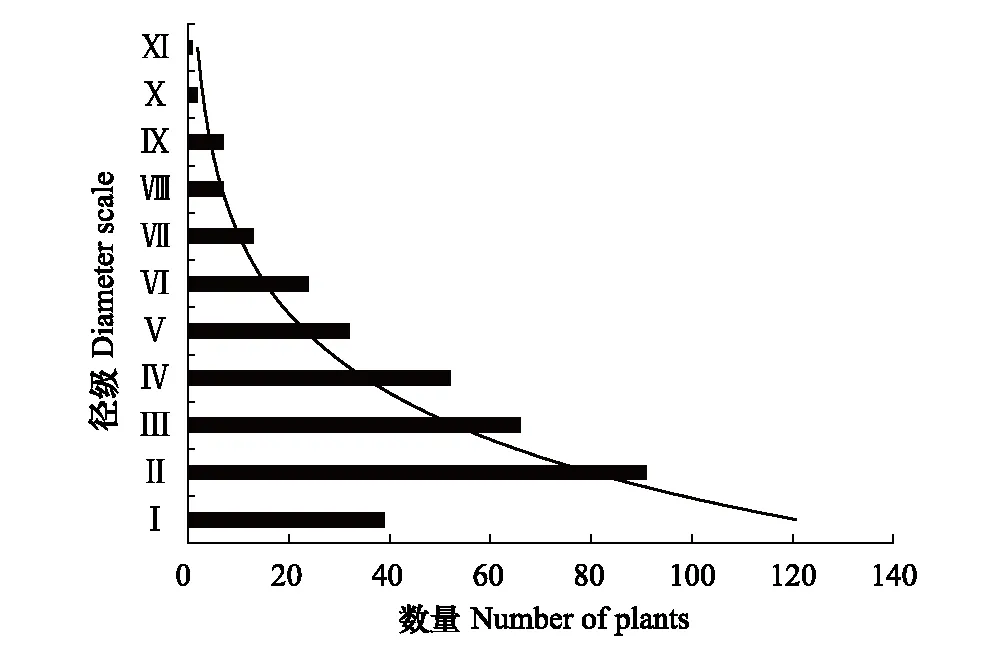

2.2 黄檗种群静态生命表


2.3 黄檗种群存活曲线

2.4 黄檗种群生存变化规律


2.5 黄檗种群时间序列预测

3 结论和讨论

