降水变化对混生红砂-珍珠生物量及分配的影响
2021-04-01单立山解婷婷
张 鹏, 李 毅, 单立山, 解婷婷
(甘肃农业大学林学院, 甘肃 兰州 730070)
降水作为干旱、半干旱生态系统植物生长和发育主要限制因子和不同时空尺度上各种生物过程的关键驱动因子,其变化对植物的生理生态过程、干物质积累与分配及生态系统结构和功能会产生深远影响[1-3]。因此,在降水格局改变的背景条件下,研究荒漠优势植物生物量以及分配对降水变化的响应机制具有重要意义。
生物量是研究生态系统结构和功能的基础[4]。生物量在各器官间的分配反映了不同的生态策略[5],是植物对环境长期适应的结果[6],对研究个体生长、物种共存、植被恢复具有重要作用[7]。在全球气候变化背景下,降水变化对植物生物量分配模式的影响受到了广泛关注[8-9]。大量研究表明,植物地上生产力与年降水量呈显著正相关[10-12],通常轻度土壤干旱不会导致植物生物量明显减少,而中度或更严重的干旱则使植物光合能力显著降低[13-14]。同时降水变化还可能改变植物营养物质利用,进而改变植物生物量分配[1]。根据最优分配假说,为响应不同环境条件的变异,植物会通过向不同器官分配生物量来获取养分、水分和光使其生长最大化[15-16]。故随着降水量减少,植物会分配更多的生物量到地下,而导致地下生物量/地上生物量增加[9]。然而,有研究指出,由于受土壤异质性或生态系统的物种组成不同等因素的影响,降水量的改变最终对生态系统水平地下生物量/地上生物量可能无显著影响[17]。可见,对降水变化如何影响生物量的分配目前仍存在较大的争议。虽然也有研究指出我国荒漠生态系统生物量在不同器官之间的分配对气候变化并不敏感[18-19],但目前该类研究大多集中在森林和草地生态系统[20],关于灌木特别是荒漠典型混生灌木植物生物量分配格局及其与降水变化关系的研究还较为缺乏[21]。因此,开展不同降水量条件下荒漠混生灌丛生物量及分配特征的研究,对认识全球气候变化背景下荒漠灌丛生态系统的物种多样性维持和碳汇功能具有重要意义。
红砂(Reaumuriasoongorica) 和珍珠(Salsolapasserina)是干旱荒漠和荒漠草原区分布最广的植物种,两者常互为共建种构成混生群落,混生群落适应极端环境的能力要高于单生群落,这种混生群落对维持生态系统稳定和高效利用降水资源有着极为重要的作用[22]。近年来,人工控制试验条件下红砂和珍珠混生后两者的生理变化特征正渐渐受到人们的关注[23-24],就水分胁迫下红砂和珍珠混生后其垂直根系变化也有一些零星报道[25-26]。然而,红砂和珍珠组成的混生群落广泛分布于我国荒漠东部和荒漠草原区,有多样的生境类型,由东向西其气候形成光-温-湿的明显梯度和多种多样的水热组合,在我国形成了沿降水梯度分布格局。这种沿降水梯度分布的混生群落为我们进行人工模拟降水梯度设计提供了很好的参考。为此,本研究根据红砂-珍珠群落自然分布区的降水变化,模拟不同降水量,对单生和混生红砂和珍珠的生物量及分配进行了研究,旨在为分析降水变化下荒漠生态系统中优势物种生物量积累及分配策略以及植物在各器官间的分配过程和贮藏比例、碳素在地上地下的分配格局等提供重要的理论依据。同时有利于进一步解释荒漠植物种间相互作用机制,对指导胁迫环境下的植被恢复具有重要的意义。
1 材料与方法
1.1 苗木的培育
在白银市景泰县山区(103°33′~104°43′ E,36°43′~37°38′ N)戈壁红砂和珍珠混生群落自然分布区内,挖取平均株高10 cm、株幅10 cm的红砂和珍珠幼苗。带回校内试验样地后,种植在高度80 cm、直径为30 cm的PVC管内,缓苗30 d。试验设计两种栽植方式,一种是红砂和珍珠分别单生栽植,另一种是红砂和珍珠二者混生栽植,各36盘。将PVC一分为二,用胶带重新固定后装入黄土、蛭石和珍珠岩1∶1∶1的混合物组成的土壤,放置在稍大的托盘上,以防土壤下沉及其水分流失。有降雨时及时对种植的红砂和珍珠用遮雨布进行遮雨,降雨过后取走,保证植物可以正常生长[32]。
1.2 试验设计
本研究根据红砂-珍珠自然分布区由东向西的分布规律,以中国气象局近58年的数据作为参考,选取6个地点的降水量作为参照[27](其年均降水量及气候类型如表1所示)。
依照6个选取地点的气象数据统计,人工模拟试验年降水量分别设置为300,250,200,150,100和50 mm。因此,在模拟降水量的试验中,分别计算出6—9月的月降水量占比并换算为浇水量,并依据梯度设置浇水频次[28-30]。试验设置6种降水量,重复3组,其年降水量浇水频次依次为:300,250 mm的为每月8次;200,150 mm的为每月6次;100,50 mm的为每月4次,浇水时间从6月1日—9月30日,模拟降水时间为19点,模拟降水量见表2。10月中旬选择晴朗天气进行样品采集[27]。
降水量计算方法:在直径为20 cm的雨量器内,每降水31.4 mm当于自然降水1 mm,即:3.14×(10 cm)2/31.4 cm3=S/A,式中,A代表降水量,S代表降水面积,即在面积为S的地面上降水A mL相当于1 mm降水[27,31-32]。
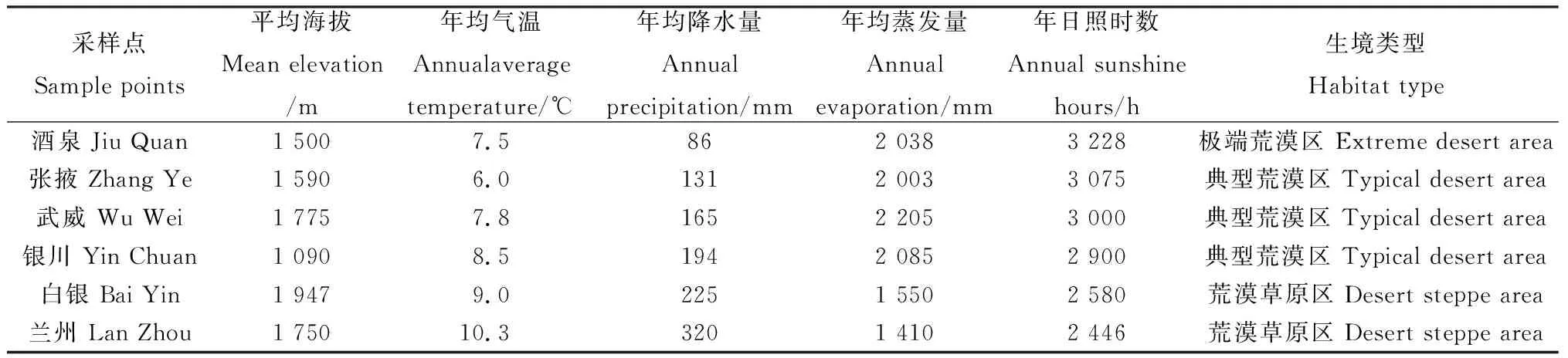
表1 试验设计参照的试验6个选取点的水热条件比较Table 1 Comparison of hydrothermal conditions of six selected test points with referred to experiment design

表2 降水量占比及浇水量换算Table 2 Precipitation and conversion of irrigation volume
1.3 地上生物量的测定及指标计算
植物样带回试验室,把根、茎、叶分开后分别在105℃下放置30 min杀青,利用80℃烘箱烘至恒重后用天平称重,再进行指标计算。相关指标计算如下:
①叶生物量比重=叶生物量/总生物量;
②根生物量比重=根生物量/总生物量;
③茎生物量比重=茎生物量/总生物量;
④叶根比=叶生物量/根系生物量;
⑤根冠比=根生物量/地上部分生物量;
⑥源汇比=叶生物量/非叶生物量。
注:非叶生物量=总生物量—叶生物量。
1.4 数据处理与分析
数据采用SPSS 22.0进行统计分析,采用双因素方差分析处理降水量和生长方式对植物生物量的影响;对同一生长方式不同降水量的植物生物量利用方差分析进行差异显著性检验;利用独立样本t检验对同一降水量不同生长方式的植物生物量进行差异显著性检验。采用Origin 2018作图。
2 结果与分析
2.1 不同降水量条件下单生与混生红砂和珍珠生物量变化特征
2.1.1总生物量 两种荒漠植物总生物量受到生长方式显著影响,红砂达到显著水平(P<0.05)、珍珠达到极显著水平(P<0.01)(表3)。由图1可知,与单生相比,除50 mm降水处理外,各降水量条件下混生红砂总生物量均小于单生红砂,平均降低了32.01%。各降水量条件下混生珍珠总生物量远小于单生珍珠,平均减少70.51%,在50 mm和300 mm年降水量处理下混生珍珠总生物量极显著低于单生珍珠(P<0.01)(图1)。对同一生长方式下不同物种总生物量进行比较,发现单生珍珠总生物量远大于单生红砂,混生珍珠总生物量总体小于混生红砂。随降水量增加,两种荒漠植物单生与混生总生物量并未呈线性增加,红砂在降水量200 mm处理下其生物量达到最大,50 mm和200 mm年降水量处理下单生红砂总生物量差异显著(P<0.05)(图1)。
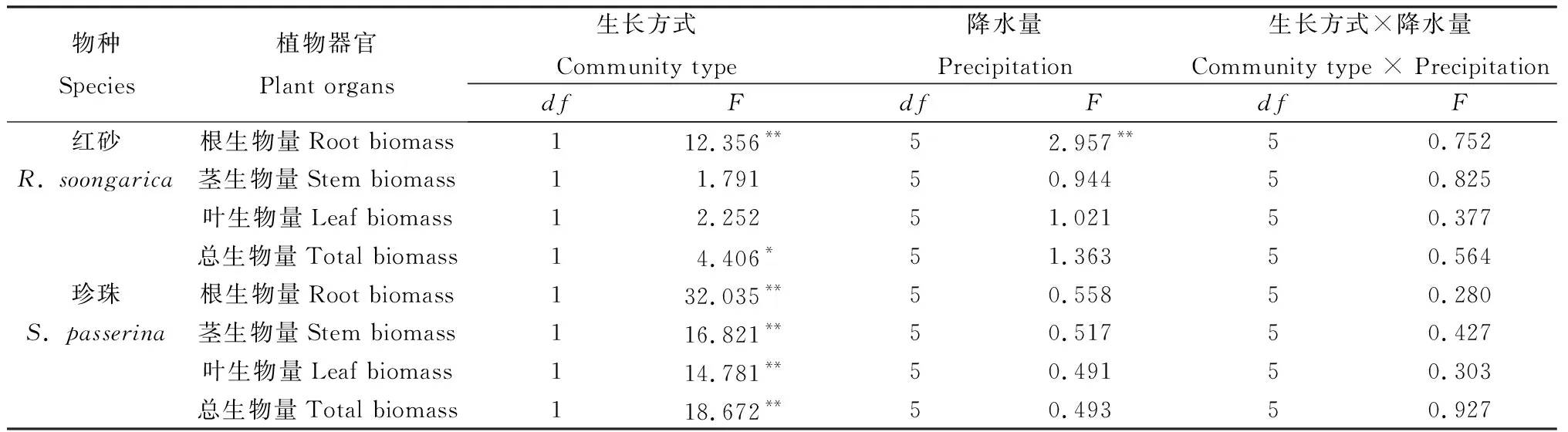
表3 不同降水量下单生与混生红砂和珍珠总生物量和根、茎、叶生物量双因素方差分析(F值)Table 3 Two-way ANOVA (F-values) of effects of precipitation and community type on R. soongarica and S. passerine in monocultural and mixed conditions in total biomass,root biomass,stem biomass and leaf biomass

图1 不同降水量条件下单生与混生红砂和珍珠总生物量变化特征Fig.1 Total biomass of R. soongorica and S. passerine in monocultural and mixed conditions under different precipitation注:不同大写字母表示在相同水分处理下,不同生长方式下各指标之间差异显著(P <0.05),不同小写字母表示相同生长方式下,不同降水处理下各指标之间差异显著(P<0.05),下同Note:Different uppercase letters indicate significant difference between different interspecific relationship at same precipitation at the 0.05 level,different lowercase letters indicate significant difference between different precipitation at same interspecific relationship at the 0.05 level,the same as below
2.1.2根、茎、叶生物量 生长方式对红砂根生物量、珍珠根、茎、叶生物量有极显著影响(P<0.01)(表3)。与单生相比,混生红砂除在50 mm年降水量处理下叶、茎生物量和300 mm年降水量茎生物量略有增加外,其余各降水量条件下叶、茎、根生物量均小于单生,分别平均减少27.75%,32.12%和38.40%,在200 mm年降水量处理下混生红砂根生物量显著小于单生红砂(P<0.05)(图2)。就珍珠而言,各降水量条件下混生红砂叶、茎和根生物量均小于单生珍珠,且比混生红砂减少得更显著,分别平均减少了66.68%,73.78%和71.17%,其中在50 mm和300 mm年降水量处理下,单、混生珍珠根、茎、叶生物量的差异极显著(P<0.01)(图2)。
红砂根系生物量受到降水量的极显著影响(表3)。随降水量的增加,单生与混生红砂根、茎、叶生物量总体呈现出先增大后减小的趋势;在降水量为200 mm处理下,红砂根、茎、叶生物量达到最大值。单因素方差分析表明,单生红砂在50 mm与150 mm,200 mm年降水量处理下,混生红砂在50 mm与300 mm和100 mm与300 mm年降水量处理下根生物量差异均显著(P<0.05)(图2)。随降水量增加,混生珍珠根、茎、叶的生物量均呈现先增大后减小,在150或200 mm降水量条件下,其生物量达到最大值。单因素方差分析表明,混生珍珠在50 mm和200 mm,200 mm和250 mm,200 mm和300 mm年降水量处理下茎生物量差异均显著(P<0.05)(图2)。
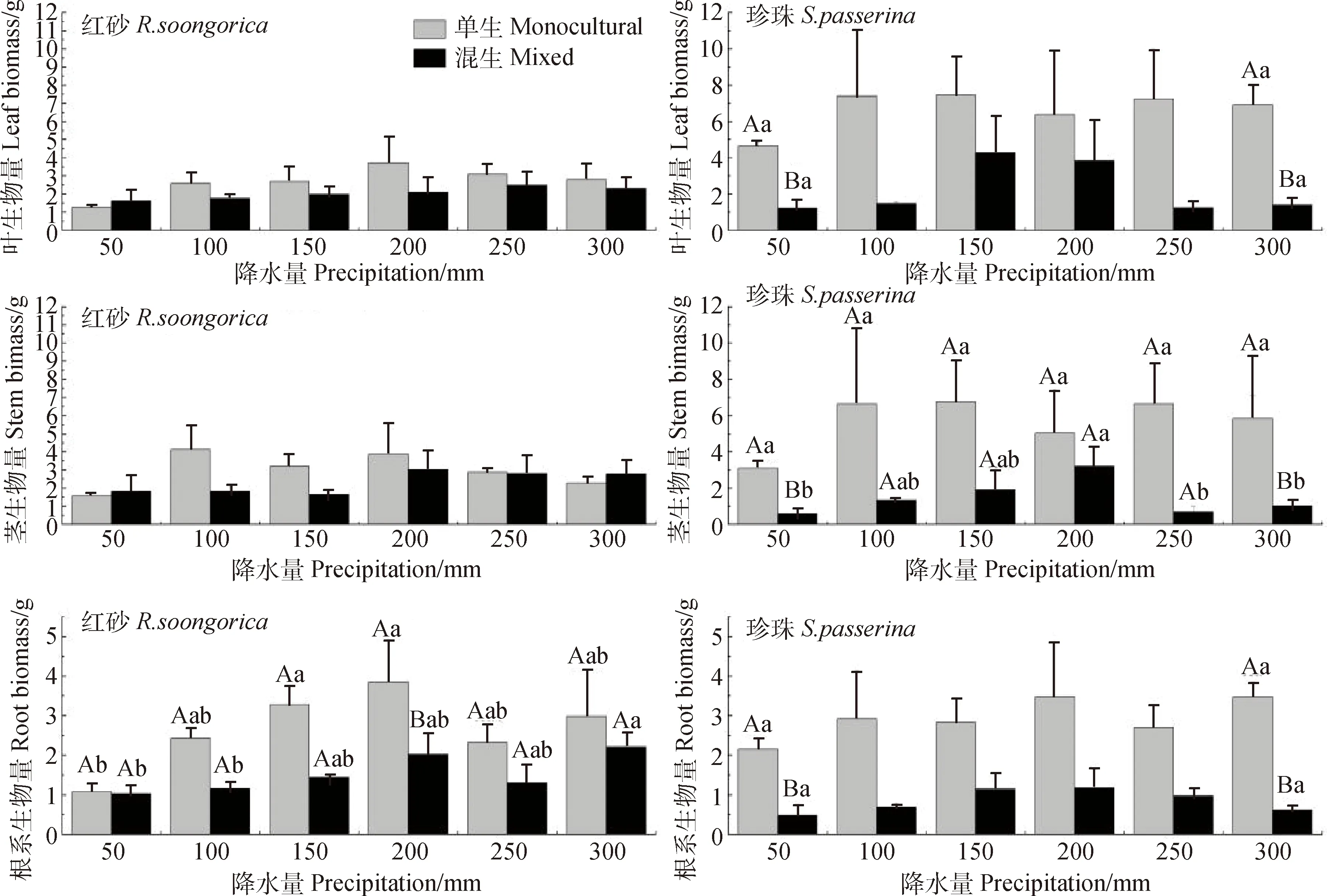
图2 不同降水量下单生和混生红砂与珍珠的根、茎、叶各构件生物量变化特征Fig.2 Component biomass of R. soongorica and S. passerine in monocultural and mixed conditions in leaf,stem and root under different precipitation
2.2 不同降水量条件下单生与混生红砂和珍珠生物量分配特征
2.2.1根、茎、叶生物量比 如表4所示,混生对红砂叶、茎生物量比以及降水对红砂根、茎、叶生物量比均无显著影响,但混生显著改变了红砂根生物量比重(P<0.05)。与单生相比,各降水量条件下混生红砂根系生物量比均减少,平均降低了15.43% (P<0.05);然而,混生红砂叶、茎生物量比较单生要高,分别增加了9.29%,5.51%,但差异不显著。就珍珠而言,混生对其根、茎、叶生物量比以及降水对根、茎生物量比无显著影响,但降水对珍珠叶生物量比有显著影响(P<0.05),且在降水量150 mm处理耦合混生条件下珍珠叶生物量比重达到最大值,为57.27%。生长方式和降水交互作用也显著改变了珍珠茎和根生物量比重,尤其在混生耦合200 mm降水量处理下珍珠茎生物量比重达到最大(图3b),为47.48%;在混生耦合250 mm降水量处理下珍珠根生物量比重达到最大(图3b),为36.08%。
对红砂和珍珠根、茎、叶生物量比重进行了分析,发现红砂茎生物量比重最大,为36.75%,即红砂总物量主要分配在茎上,其生物量分配为:茎>叶>根;而珍珠则是叶生物量比较大,为45.41%,即珍珠总生物量主要分配在叶上,相较红砂增加了35.78%;珍珠根生物量比重则远小于红砂,比红砂减少了28.43%,其生物量分配为:叶>茎>根。

图3 不同降水量下单生和混生红砂与珍珠的根、茎、叶生物量比Fig.3 Component biomass and allocation proportion of R. soongorica and S. passerine in monocultural and mixed conditions in leaf,stem and root
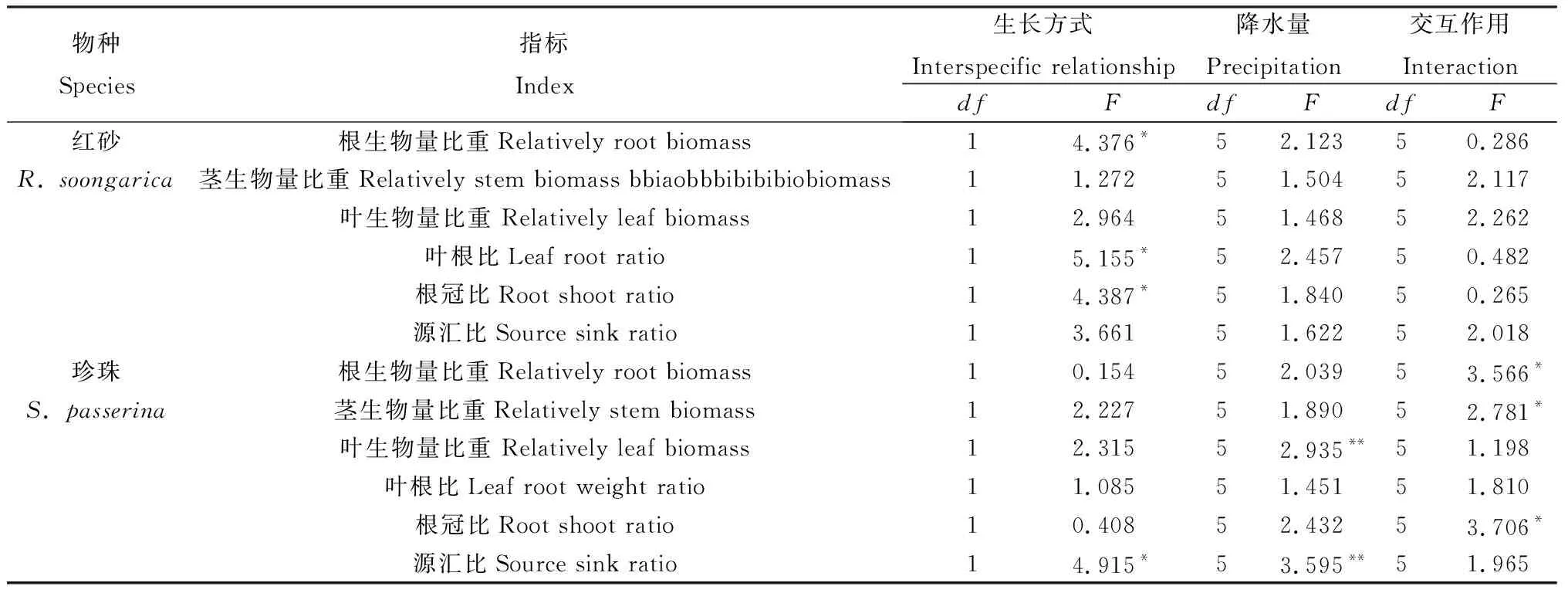
表4 不同降水量下单生与混生红砂和珍珠生物量分配指标的双因素方差分析(F值)Table 4 Two-way ANOVA(F-values) of effects of precipitation and interspecific relationship on R.soongorica and S.passerine in monocultural and mixed conditions in index of biomass allocation
2.2.2叶根比、根冠比和源汇比 由表4可知,降水量以及降水量与生长方式的交互作用对红砂叶根比、根冠比以及源汇比均无显著影响,但生长方式对红砂叶根比、根冠比有显著影响(P<0.05)。由图4a,c可知,在相同的降水量下,混生红砂叶根比都大于单生红砂,平均增加了33.19%,但各降水量混生红砂根冠比均小于单生红砂,平均减少了20.89%。就珍珠而言,源汇比受到了生长方式、降水量显著影响(P<0.05)。由图4f可知,混生珍珠源汇比基本都大于单生珍珠,平均增长量24.74%。珍珠根冠比也受到生长方式和降水量交互作用的显著影响,在混生耦合250 mm降水量处理下珍珠根冠比达到最大(图4 d),为0.58。
对同一生长方式下不同物种根冠比、叶根比和源汇比进行了比较发现,除250 mm处理下混生红砂和混生珍珠外,同一生长方式下单生、混生珍珠根冠比均小于单生、混生珍珠,分别平均减少了43.64%,43.26%。然而,同一生长方式下单生、混生珍珠叶根比、源汇比却大于单生、混生红砂,分别平均增长了108.93%,80.36%,69.04%,81.54%。

图4 不同降水量下单生、混生红砂珍珠的叶根重比、根冠比和源汇重比变化特征Fig.4 Leaf root weight ratio,root shoot ratio and sink weight ratio of R. soongarica and S. passerine in monocultural and mixed conditions
3 讨论
3.1 混生对红砂和珍珠生物量及分配的影响
研究表明,植物在生物或非生物胁迫下会衍生出一种平衡生长模式,即通过对生物量的调节分配来维持它的最优生长[33]。本研究发现,两种荒漠植物总生物量均受到生长方式显著影响,混生显著降低红砂和珍珠总生物量。然而,物种不同其总生物量减少程度及导致其总生物量减少的器官有所差异,混生红砂总生物量显著减少,降低幅度相对较少,其总生物量的减少是由根系生物量减少所致;混生珍珠总生物量极显著减少,降低的幅度较大,且在降水量较少和水分条件较好情况下效应更明显,其总生物量减少是由根、茎、叶生物量共同所致。说明两种荒漠植物混生后,其种间竞争作用强烈,其生物量均受到了损失。然而,物种不同其受到的竞争胁迫所有不同,红砂因竞争作用强,只有根系生物量受到了损失,而珍珠各器官生物量均受到了较大损失,在不适应水分条件下效应更明显。这与王政权和张彦东[34]对水曲柳(Fraxinusmandshurica)和落叶松(Larixgmelinii)混栽导致落叶松生物量低于纯栽的结果一致。表明植物混生后,因种间竞争,竞争能力差的植物(比如珍珠)不易从种间关系中获益,其生物量受到较大损失。同时,本研究还发现,单生珍珠总生物量远大于单生红砂,混生珍珠总生物量却小于混生红砂,可能是由于珍珠作为C4植物,具有较高水分、养分利用效率和较高的光合作用,在没有受到邻体竞争时,本身的特性促进了植物快速生长、提高了生产力[35],因而生物量较大,而与红砂混生后,因红砂地上部分竞争能力强于珍珠[36],其生长受到抑制,其生物量受到了较大损失。表明在中国西北红砂-珍珠组成混生群落中红砂竞争力强而珍珠竞争力弱,这也是红砂在中国西北干旱和半干旱区分布的生态幅广,而珍珠分布的生态幅相对窄些的主要原因。
生物量分配指生物量在不同植物器官或功能单位中的分配方式,可指示植物之间竞争能力的强弱[37]。根据最优分配假说,植物会分配更多的生物量到能获取受限资源的器官中,以维持最大的生长速率[14,38]。本研究发现,生长方式对红砂根生物量比、叶根比和根冠比均有显著影响(P<0.05)。与单生相比,混生红砂叶根比、源汇比有不同程度增加,而根生物量比和根冠比有所下降,表明混生显著降低了红砂根系生物量,其根分配的生物量减少,而地上分配的生物量增加。这也说明红砂和珍珠混生后,地下根系竞争强烈,两种荒漠植物根生长均有所抑制(图2),且对红砂影响更显著,即红砂生物量向根系分配更小,从而导致了叶根比增加、根冠比降低[20,39]。对于珍珠而言,生长方式只对源汇比有显著影响(P<0.05),混生显著提高了珍珠源汇比。表明红砂-珍珠混生后,因地上遮阴对珍珠光合作用产生了影响,且珍珠为C4植物,有较高的光合作用,混生后光照成为其限制因子。根据最优分配假说,植物会分配更多的生物量到能获取受限资源的器官中,以维持最大的生长速率[7],因此,珍珠将叶分配更多的生物量来减少光照的限制,其源汇比增加。混生条件下红砂-珍珠生物量分配所表现出来的差异,说明不同植物在遭受非生物胁迫压力时采取了不同的应答机制,也说明植物对非生物胁迫的响应比我们所认知的复杂得多[40]。本研究还发现,两种荒漠植物生物量在不同器官分配存在差异,红砂主要分配在茎上,而珍珠则主要把生物量分配在叶上,这可能与两种荒漠植物生物学特性有关。红砂为复苏植物,在西北干旱生境条件下其叶片会大量脱落,为了适应干旱环境,红砂较高比例的生物量分配到茎,一方面减少了因西北干旱胁迫而在植物木质部产生气穴现象,另一方面,可以存储大量的水分,有助于缓解因茎部水分含量减少引起的水分渗透[8]。而珍珠为C4植物,较高比例叶的生物量分配维持了较高的光和作用[26]。
3.2 降水对红砂和珍珠生物量及分配的影响
不同降水梯度上植物生物量及在各器官的分配存在较大差异[41]。一般认为,适度的增加降水会提高植物的光合能力,增加植被的净初级生产力,干旱则会抑制植物生长,改变干物质分配比例,降低干物质积累[1,42]。本研究发现,单生与混生红砂总生物量、根、茎和叶生物量均随着降水量的增加呈现出先增加后减少的变化趋势,在200 mm降水条件下,其总生物量和各器官生物量达到最大值,这与Chen等[43]对古尔班通古特沙漠降水增加限制了Nepetamicrantha生长的结论一致。表明红砂对降水量响应存在阈值效应,适当增加降水量有利于其生长,而较好水分条件其生长受到抑制。其主要原因是红砂为主根型植物,即主根明显侧根相对较少[26],这种主根型植物比须根植物从空间和时间上不均匀的土壤中捕获移动离子更少,降水增加加快了氮素损失[43],进而在氮素本身匮乏的西北干旱区降水增加抑制了其生长和生物量的累积。然而,在森林和草地生态系统中,降水变化与生物量的累积呈显著的正相关[10-11,44],表明植物对降水增加的响应具有种间效应。对于红砂而言,年降水量超过200 mm以后,其生长将受到抑制,且在降水增加并存在种间竞争存在时(如珍珠),其抑制效应更明显。
本研究还发现,降水量对红砂根生物量产生了极显著影响(P<0.01),而对茎和叶生物影响不显著,表明红砂不同器官对降水变化的响应敏感程度不同,降水变化红砂幼苗根系最先感知,采取“积极”调整策略来适应变水环境[45],这与荒漠植物梭梭(Haloxylonammodendron)、多枝柽柳(Tamarixramosissima)降水变化的响应一致[46]。混生条件下,珍珠根、茎和叶生物量及总生物量也均随降水量增加呈现出先增加后减少的变化趋势,表明在种间竞争和水分(增加或减少)双重胁迫下,其生长均受到抑制,各器官生物量均损失。方差分析表明,珍珠只有叶生物量比和源汇比受降水量的显著影响,这表明珍珠叶对降水变化较敏感,这也符合珍珠的生物学特性,水分变化则更多进行叶生物量的调整来适应变水环境。
4 结论
本研究发现,生长方式对两种荒漠植物生物量有显著影响。与单生相比,混生红砂与珍珠总生物量及各器官生物量均减少,混生红砂总生物量减少较少,其减少量由根系生物量决定,而混生珍珠总生物量减少较大,由根、茎、叶生物量共同决定。生长方式对红砂-珍珠生物量的分配过程的影响表现在两种混生植物将生物量更多的分配到地上部分。生物不同其生物量在不同器官分配也存在较大差异,红砂主要将生物量分配给茎,而珍珠将生物量主要分配给叶,这由两种荒漠植物生物学特性决定。降水量对红砂的根生物量有极显著影响、对珍珠叶生物量比有显著影响,表明红砂幼苗根系对降水变化的反应较为敏感、而珍珠叶片对降水变化最敏感。
