典型喀斯特高原水库浮游植物与环境因子的关系
2021-03-09葛秋诗张萍倪茂飞郭云王志康周慧张志威周业
葛秋诗,张萍,倪茂飞,郭云,王志康,周慧,张志威,周业
贵州民族大学生态环境工程学院,贵州 贵阳 550025
浮游植物是水生生态系统的初级生产者,由于其群落结构对自身生存环境变化较为敏感,也可作为水环境特征的重要指示生物(胡芳等,2014;Lepisto et al.,2004;李然然等,2014)。因此,对水体浮游植物群落特征与环境因子间关系的研究十分必要。影响水体浮游植物密度及群落变化的环境因子具有时空差异,总体而言,水温(tw)、透明度(SD)和营养物质作为自然水体生命活动的重要指标,通常与水生生物过程具有显著的相关关系(邓乐等,2019;邹曦等,2017;邱小琮等,2012)。此外,其他因素如水利调节、化学计量比、溶解氧(DO)和pH等能够显示水生环境情况,因此可能极大地影响浮游植物密度与群落特征(李飞鹏等,2013;葛优等,2017;李磊等,2015)。
水库是一种介于河流和湖泊间的半人工半自然水体,具有城市生活与工业供水等重要功能(林秋奇等,2001)。入库水流在为水库补给水源的同时,也为水库提供营养来源。农业、城市和郊区径流和生活废水排放的营养盐过量将导致浮游植物过量繁殖(杨朝霞等,2019;Howarth et al.,2002),从而对水库水质及水厂后续处理产生恶劣影响。贵州省受喀斯特地貌影响,一方面,该地区受地质条件影响天然湖泊较少,需依靠人工建造的水库以满足城市生产生活用水(晏妮,2006),因而水库水质与居民生活质量息息相关。另一方面,该地区水质具有偏碱性、钙镁含量高的特点(高廷进等,2014),白宁静等(2019)研究认为,钙离子是云贵高原地区湖泊中浮游植物群落构建的重要影响因子,说明喀斯特地区水体具有更加独特的浮游植物群落特征。综上所述,针对该地区水库浮游植物群落特征与其相应的环境因子进行研究,具有十分重要的意义。
本文以典型喀斯特水库——阿哈水库为研究对象,通过对入库支流、库中心和南郊各采样点不同季节(春、夏和秋季)的水体进行分析,探索了喀斯特水库基础水环境指标的量级、浮游植物群落组成、优势种变化及其密度的时空分布,进一步揭示浮游植物与环境因子的耦合关系,为喀斯特高原水库浮游植物研究以及水资源保护提供一定的科学依据。
1 材料与方法
1.1 采样地概况及采样点设置
阿哈水库(106°39′E,26°33′N)位于长江流域乌江水系,属于多入库河流型水库(高廷进等,2014),位于贵阳市南郊,由小车河上游的游鱼河、白岩河、金钟河、烂泥河以及蔡冲沟交汇而成。水库汇水面积190 km2,正常蓄水位1110 m,对应水库容积5.35×107m3,平均水深19 m;多年平均径流量 3.24 m3∙s−1,年均水输送量 1.04×108m3。年均气温为15.3 ℃(万国江等,2001),是贵阳市主要饮用水源地之一。
本研究在湖库内共设7个采样点,分别为游鱼河(YY)、蔡冲沟(CC)、烂泥河(LN)、南郊水厂取水口(NJ)、水库中心(KZ)、白岩河(BY)、金钟河(JZ),分上、中、下三层取样,其中蔡冲沟、烂泥河处较浅,取上下两层。全年春、夏、秋三季共计取水样 57份,冬季喀斯特地区浮游植物密度较少,且与秋季同属枯水期(罗宜富等,2017),水文条件稳定,所以未将冬季纳入研究范围。采样点的分布见图1。
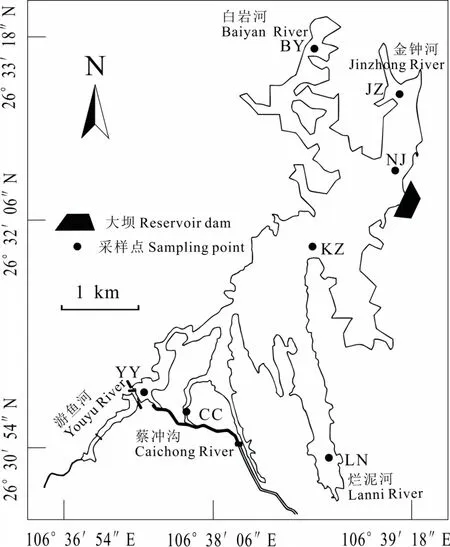
图1 阿哈水库采样点分布Fig.1 Location of sampling sites in Aha Reservoir
1.2 样品采集、处理与分析
1.2.1 野外采样
样品采集工作分别于2019年4、7、10月开展,共设置7个采样点包含水库主体及各支流,保证其地理位置具有空间代表性。3次采样时间均为0:900—11:00,以避免浮游植物随光照变化造成垂直移动现象。利用5 L有机玻璃采水器分3层(表层:0.5 m;中层:3 m;下层:5—8 m)取各点水样,其中取1 L用于测定理化指标,另取500 mL现场加入7 mL左右鲁哥氏液固定,利用25号浮游生物网于表层至0.5 m水深处作“∞”形缓慢循环拖动1—3 min,收集水样并加入福尔马林固定液使其浓度约为4%,作为浮游植物定性样品。用于测定理化指标的水样低温运送回实验室后,立即放入冰箱4 ℃保存并尽快测定,其余浮游植物指标水样常温下保存并及时测定。
1.2.2 样品预处理
浮游植物定量样品在实验室静置24 h,吸去上清液,并浓缩定容至30 mL定量瓶。将样品充分摇匀后,吸取0.1 mL至计数框,利用400倍显微镜对浮游植物进行定量,每份样品观测两次,结果取平均值。
1.2.3 实验室分析
水库tw和DO利用便携式溶解氧仪(JPB-607A,雷磁,上海)进行测定,其中DO饱和度计算公式为:DO饱和度(%)=(溶解氧实测含量/实测条件下溶解氧的饱和含量)×100%,pH 利用便携式 pH 计(PHB-4,雷磁,上海)测定,SD利用塞氏盘现场测定。总磷(TP)采用钼酸铵分光光度法、钙离子(Ca2+)浓度采用EDTA滴定法测定,方法参照《水和废水分析监测方法》(国家环境保护局,1997)。总氮(TN)采用总有机碳分析仪(Vario TOC,Elementar,德国)的总氮(TNb)分析模块测定,氨态氮(NH3-N)利用水杨酸钠法通过全自动间断化学分析仪(Cleverchem380,Dechem-Tech,德国)测定。氮磷比(N/P)为TN与TP的质量比值。浮游植物定性与定量分析依照《水和废水分析监测方法》与《中国淡水藻类》(胡鸿钧等,2006)所述方法进行。
1.3 统计分析
对不同季节测定的8种环境因子(tw、DO、SD、pH、TN、TP、Ca2+和NH3-N)与浮游植物密度(D)结果进行分析。浮游植物与环境因子的季节差异通过单因素方差分析(Analysis of Variance,One-way ANOVA)判断;浮游植物密度与环境因子间关系通过一元线性回归分析(Linear Regression Analysis)反映。
利用 Canoco 5软件对浮游植物与环境因子进行排序分析,浮游植物优势种类的筛选标准应满足:该物种在各点出现的频率>12.5%,且至少在一个点的相对密度>1%(Lopes et al.,2005;Muylaert et al.,2000)。首先,将除pH外的环境因子及浮游植物数据进行lg(x+1)转换,并对浮游植物密度数据进行去趋势对应分析(Detrended Correspondence Analysis,DCA)。DCA结果显示,春、夏、秋的最大长度梯度依次为0.84、0.27、0.62,均小于3,因此采用基于线性的冗余分析(Redundancy Analysis,RDA)分析浮游植物与环境因子间的关系较为合适。
One-way ANOVA由IBM SPSS Statistics 20软件完成,线性回归分析及绘图由Origin 2018软件实现。
2 结果与分析
2.1 水体基础环境因子
如表1所示,tw、DO、pH、SD、TN、NH3-N、和 Ca2+具有显著的季节差异(One-way ANOVA,P<0.01),而各季节TP和N/P不具有显著的统计性差异(P>0.05)。水库tw在夏季最高为 (26.66±1.62) ℃,秋季最低 (17.00±0.84) ℃。DO为4.50—18.8 mg∙L−1,且春季 (13.54±3.28) mg∙L−1显著高于其他季节[夏季(9.45±1.71) mg∙L−1,秋 季(6.05±1.13) mg∙L−1],春季水体 DO 饱和度最高(149%±34%),夏季次之 (118%±23%),秋季呈现缺氧状态,饱和度最低 (63%±12%),这可能与浮游植物的光合作用有关。水体pH整体呈碱性,且在7.10—8.71范围内。由于水体浮游植物活动频繁,导致夏季SD值 (0.91±0.37) m低于春季 (1.08±0.49) m和秋季 (2.50±0.29) m。TN、NH3-N作为浮游植物的营养盐,TN 夏季最高 (3.36±0.88) mg∙L−1,NH3-N春季最高 (1.382±0.451) mg∙L−1,均在秋季最低[TN:(2.09±0.21) mg∙L−1,NH3-N:(0.055±0.042)mg∙L−1]。受喀斯特地貌影响,Ca2+在 61.16—140.92 mg∙L−1之间,其中秋季最高 (92.17±6.04) mg∙L−1,夏季最低 (74.02±11.22) mg∙L−1。

表1 各季节阿哈水库的环境因子Table 1 Environmental indicators of Aha Reservoir in different seasons
空间上,春、夏季的营养盐浓度(TN、TP、NH3-N)以游鱼河最低,金钟河最高,而Ca2+浓度则与之相反,为金钟河低、游鱼河高。秋季的金钟河和南郊取水口TN、TP略高于其他位点,游鱼河的NH3-N虽然高于其他位点的2—2.8倍,但仅为0.181 mg∙L−1,Ca2+浓度各点差距较小,浓度最大的位点仍为游鱼河(101.08 mg∙L−1),即使是浓度最低的白岩河也达到 81.84 mg∙L−1。总体而言,游鱼河输入营养盐的量最少,金钟河最多,其他各点对水库的贡献介于两者之间,且由于金钟河与南郊取水口距离最近,对水厂水质的影响不容忽视。
2.2 浮游植物群落组成及优势种变化
通过对定性样品的镜检,共鉴出8门80属149种浮游植物,绿藻67种,为数量最多,硅藻42种次之,蓝藻24种,甲藻5种,金藻4种,隐藻、裸藻各3种,黄藻1种。其中,不同时期的蓝藻密度均占比最大,为64.7%—99.1%,其次是绿藻、硅藻、隐藻,密度占比分别为0.2%—22.8%、0.2%—11.0%、0—10.9%。金藻仅在春季具有数量优势,最大密度占比为4.8%,甲藻、裸藻、黄藻数量较少,密度占比小于0.8%。
根据浮游植物出现频率>12.5%和相对密度>1%的原则,筛选了22种浮游植物优势种见表2。伪鱼腥藻Pseudoanabaenasp.、水华束丝藻Aphanizomenonflos-aquae、曲壳藻Achnanthessp.为三季优势种,三季各点出现频率均为 100%;粗壮细鞘丝藻Leptolyngbyavalderiana为春、夏季优势种,春、夏各点出现频率分别为89%、100%;尖尾蓝隐藻Chroomonasacuta、卵形隐藻Cryptomonas ovata和微球衣藻Chlamydomonasmicrosphaera为春、秋季优势种,尖尾蓝隐藻和卵形隐藻在春、秋各点出现频率均为 100%,微球衣藻在春、秋各点出现频率分别为 84%、95%;狭形纤维藻Ankistrodesmusangustus为夏、秋季优势种,出现频率均为100%;华美色金藻Chromulinaelegans、湖北小环藻Cyclotellahubeiana、尖针杆藻Synedra acus、库津新月藻Closteriumkuetzingii为春季优势种,出现频率依次为89%、84%、37%、100%;两头针杆藻Synedraamphicephala为夏季优势种,出现频率为 100%;铜绿微囊藻Microcystis aeruginosa、点形平裂藻Merismopediapunctata、旋折平裂藻Merismopediaconvoluta、小空星藻Coelastrummicroporum、柱孢藻Cylindrospermumsp.、小球藻Chlorellavulgaris、啮蚀隐藻Cryptomonaserosa、卷曲纤维藻Ankistrodesmus convolutus、颤丝藻Ulotrixoscillatoria为秋季优势种,出现频率依次为32%、68%、79%、79%、95%、95%、100%、100%、100%。

表2 阿哈水库主要浮游植物优势种名录Table 2 Main phytoplankton species in Aha Reservoir
春季,华美色金藻、湖北小环藻及尖针杆藻在游鱼河、蔡冲沟两处的密度高于其他位点,而南郊取水口、库中心、白岩河、烂泥河及金钟河处的浮游植物以粗壮细鞘丝藻、伪鱼腥藻、水华束丝藻、曲壳藻、尖尾蓝隐藻、卵形隐藻微球衣藻和库津新月藻为主;夏季蔡冲沟处为粗壮细鞘丝藻密度最大,游鱼河、白岩河以两头针杆藻、曲壳藻为主,烂泥河、南郊取水口、库中心、金钟河以狭形纤维藻、伪鱼腥藻、水华束丝藻、曲壳藻为主要种类;秋季各采样点优势浮游植物种类最多,不同采样点的各类优势种密度差别不大,可能是秋季水库整体水质良好且混合均匀的缘故。
2.3 浮游植物密度时空分布
如图2所示,浮游植物密度在时间尺度上存在显著差异(One-way ANOVA,n=57,P<0.01),春、夏、秋季的浮游植物密度依次为2.55—21.10×105、3.13—43.92×105、6.79—17.45×104cells∙L−1,呈夏季>春季>秋季的趋势。春季浮游植物密度的最大点为南郊取水口(2.11×106cells·L−1),白岩河次之( 1.44×106cells∙L−1),游鱼河最低( 2.55×105cells∙L−1);夏季浮游植物密度的最大点为库中心(4.39×106cells∙L−1),南郊取水口次之(3.38×106cells∙L−1),蔡冲沟最低(3.13×105cells∙L−1);秋季浮游植物密度的最大点为烂泥河(1.74×105cells∙L−1),蔡冲沟次之(1.05×105cells∙L−1),南郊取水口最低(6.79×104cells∙L−1)。由此表明,春、夏两季水库中心区域易发生藻华。
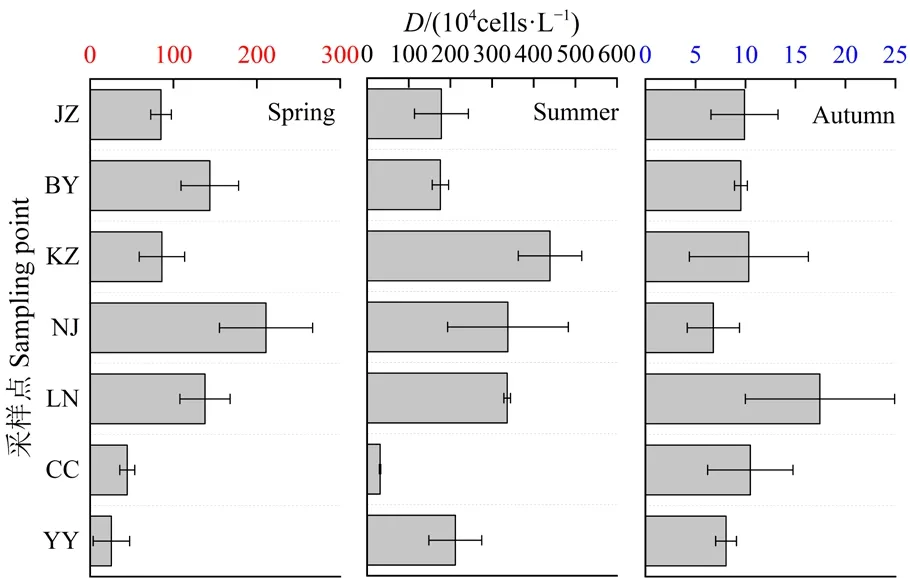
图2 浮游植物密度(D)的时空分布Fig.2 Temporal and spatial distribution of phytoplankton density (D)
2.4 浮游植物密度与环境因子的线性回归分析
不同季节浮游植物密度与环境因子间的线性回归分析如图3所示。春季浮游植物密度与众多环境因子具有显著相关关系,其与Ca2+浓度(r2=0.41,P<0.01)、N/P(r2=0.36,P<0.01)、pH(r2=0.26,P<0.05)以及 DO(r2=0.25,P<0.05)呈显著的正相关,而与 SD(r2=0.38,P<0.01)呈显著的负相关。夏季浮游植物密度仅与SD(r2=0.22,P<0.05)相关性较强,秋季浮游植物密度与所测环境因子均无显著相关关系(P>0.05)。
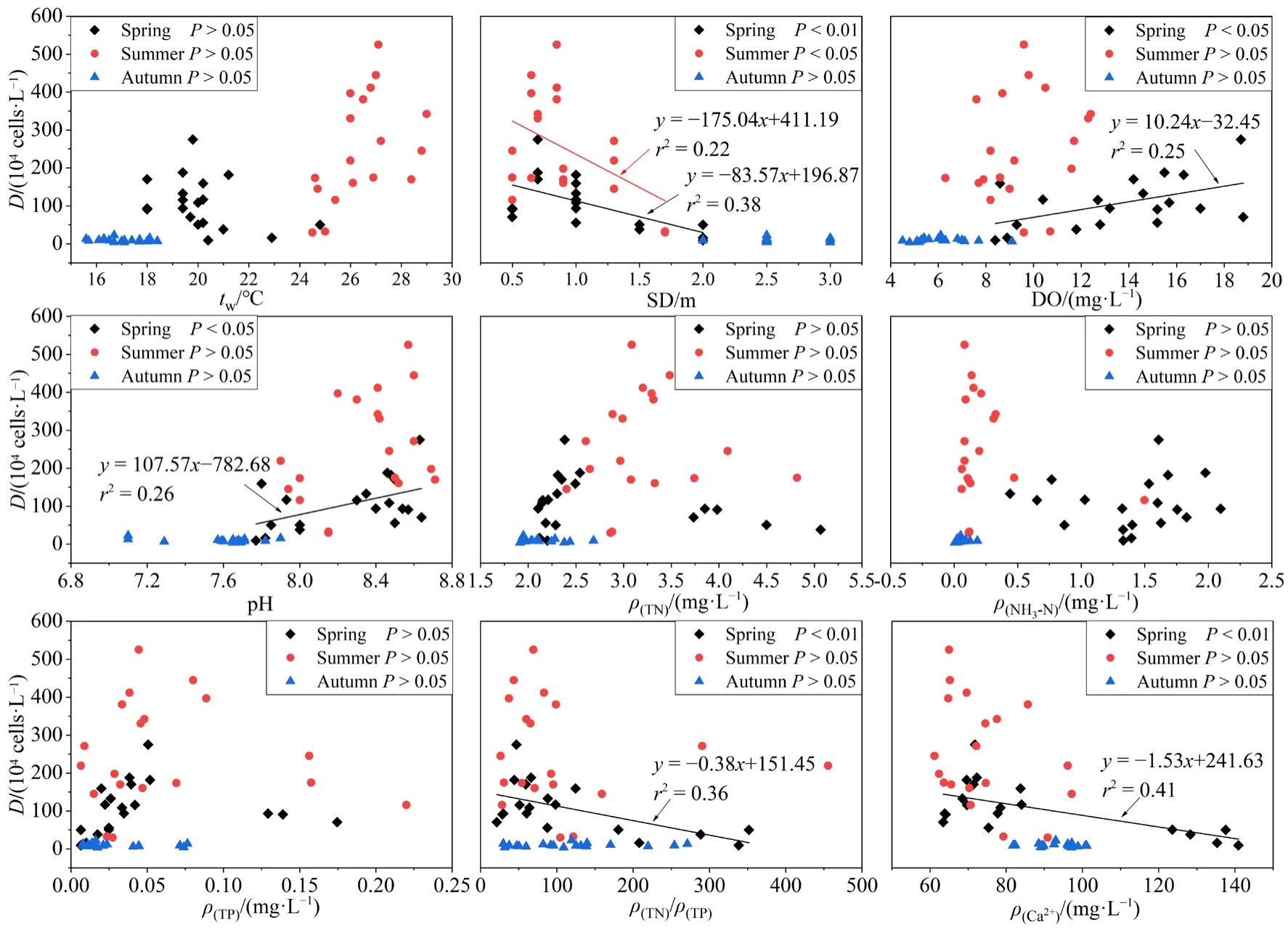
图3 浮游植物密度(D)与环境因子之间的关系Fig.3 Relationship between phytoplankton density (D) and environmental factors
2.5 浮游植物与环境因子的RDA分析
RDA分析结果见表3及图4,除秋季无环境因子入选外,春季轴Ⅰ和轴Ⅱ的特征值分别为 0.488和 0.1855,夏季轴Ⅰ和轴Ⅱ的特征值分别为0.445和0.1388,入选环境因子共解释了春季76%、夏季68%的物种变化信息。前两轴累计解释了春季67.35%、夏季 58.38%的物种变化信息和春季88.7%、夏季 85.9%的物种-环境关系信息;春、夏季的两个物种排序轴近似垂直,相关系数分别为−0.0451、−0.0149,环境因子轴与物种排序轴之间的相关系数春季为0.9672、0.8536,夏季为0.9527、0.8388,春夏季两个环境排序轴的相关系数均为0,两季节的Monte Carlo置换检验所有排序轴均达到显著水平(P<0.05),说明排序结果可信(Ter Braak et al.,1988),排序轴与环境因子间的线性结合程度较好地反映了浮游植物与环境之间的关系。

表3 浮游植物与环境因子的RDA统计结果Table 3 Redundancy analysis results of phytoplankton and environmental factors
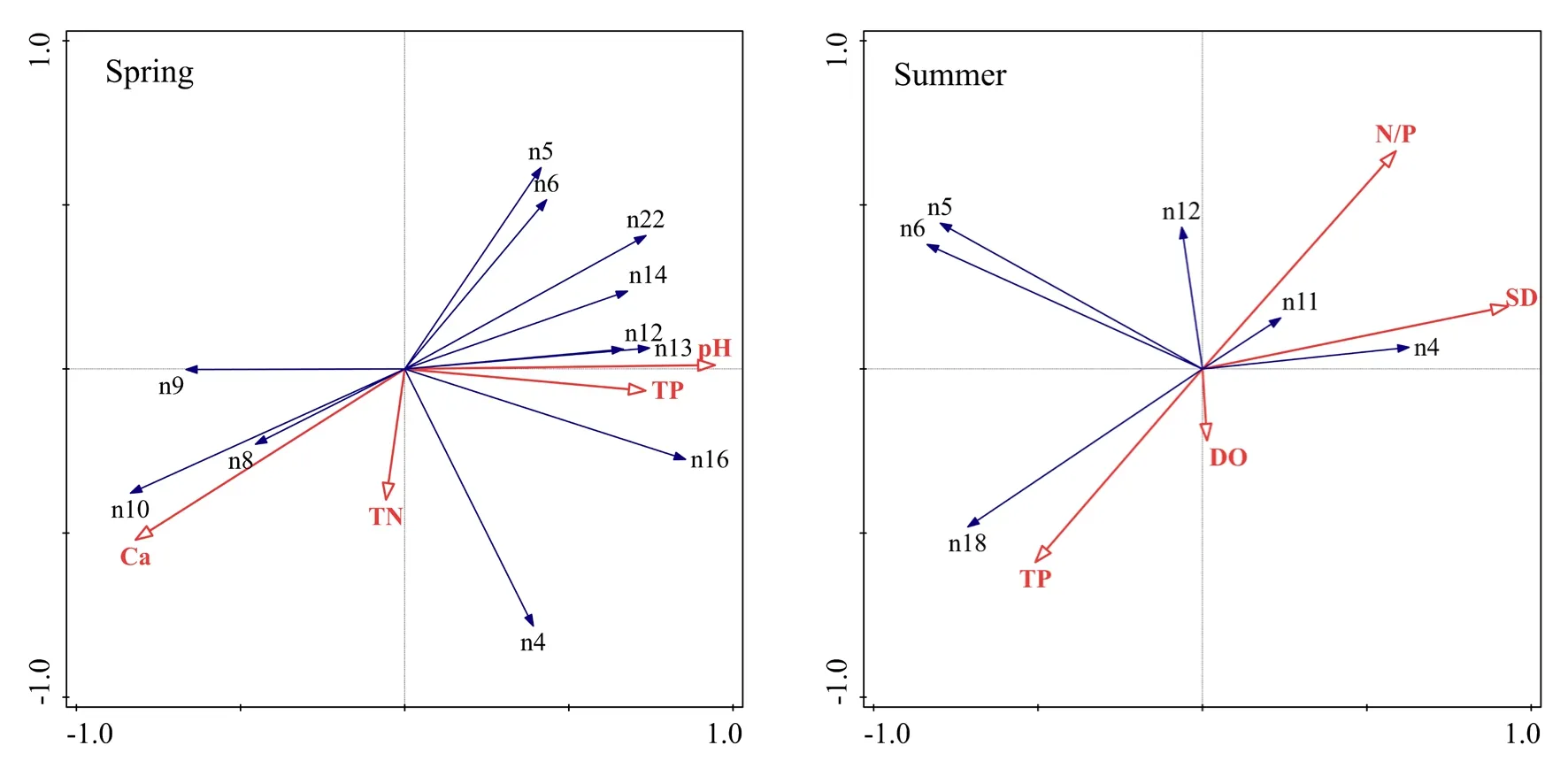
图4 不同季节浮游植物-环境因子关系的RDA分析Fig.4 RDA of phytoplankton environmental factors in different seasons
利用向前引入法(Forward Selection)对环境因子进行筛选,将Monte Carlo置换检验结果为P<0.05的环境因子作为该季节的主要影响因子参与 RDA分析,春季的主要影响因子为pH、Ca2+浓度、TP、TN,夏季为SD、DO、N/P、TP,秋季浮游植物优势种类组成较复杂,未有环境因子入选(P>0.05)。春季影响浮游植物分布的环境因子中,pH、Ca2+浓度、TP为第Ⅰ轴的主要因子,TN为第Ⅱ轴的主要因子。多数优势种与pH、TP呈正相关,与Ca2+浓度呈负相关;其余优势种,如华美色金藻(n8)、湖北小环藻(n9)、尖针杆藻(n10)与 Ca2+浓度呈正相关,与 pH、TP呈负相关;粗壮细鞘丝藻(n4)、华美色金藻(n8)、尖针杆藻(n10)与TN呈正相关,伪鱼腥藻(n5)、水华束丝藻(n6)、卵形隐藻(n14)、库津新月藻(n22)与TN呈负相关。夏季影响浮游植物分布的环境因子中,SD为第Ⅰ轴的主要因子,TP、N/P、DO为第Ⅱ轴的主要因子。粗壮细鞘丝藻(n4)、两头针杆藻(n11)与SD呈正相关,伪鱼腥藻(n5)、水华束丝藻(n6)、狭形纤维藻(n18)与SD呈负相关;粗壮细鞘丝藻(n4)、两头针杆藻(n11)、曲壳藻(n12)与N/P呈正相关,与TP呈负相关;狭形纤维藻(n18)与 TP呈正相关,与 N/P呈负相关;狭形纤维藻(n18)与DO呈正相关,伪鱼腥藻(n5)、水华束丝藻(n6)、两头针杆藻(n11)、曲壳藻(n12)与DO呈负相关。
总体而言,影响春季浮游植物群落结构的主要环境因子是pH、Ca2+浓度和TP,而夏季的主要环境因子为SD、TP和N/P。
3 讨论
3.1 阿哈水库浮游植物群落特征
采样期间阿哈水库浮游植物密度在 3.95—525.35×104cells∙L−1之间(图 2),春、夏季均为藻华暴发的高风险时期,浮游植物密度可达 106cells∙L−1数量级,采样期间浮游植物数量以蓝藻、硅藻、绿藻、隐藻居多,属于富营养型湖泊(庞清江等,2003)。另外,金藻也是春季的优势种之一,在游鱼河处密度最大,占总密度的1.88%,远高于其他点位(<0.7%),金藻常生长于温度较低、有机质含量少的清洁水体(胡鸿钧等,2006),表明游鱼河区域水体营养程度相对较低。与 2012—2013年对阿哈水库浮游植物群落的调查结果(罗宜富等,2017)相比,甲藻与微囊藻数量均明显减少,伪鱼腥藻、水华束丝藻等丝状蓝藻成为水库最大优势种。
3.2 浮游植物密度与环境因子间的关系
根据线性回归可知(图3),影响浮游植物密度的环境因子,春季为 SD、DO、pH、Ca2+浓度和N/P,夏季为 SD。SD由水体泥沙颗粒物与浮游植物含量共同决定,SD随着浮游植物密度的增长而降低(P<0.05),说明SD很大程度上是由浮游植物密度决定的,因此将SD看作浮游植物的被动影响因子。春季DO出现过饱和状态且随浮游植物密度的增加而升高(P<0.05),说明浮游植物光合作用释放氧气是水体DO偏高的重要因素之一,同时,浮游植物活动也将引起pH升高(Ouyang et al.,2018),再次验证了春季浮游植物与 SD、DO、pH之间存在密切联系。阿哈水库Ca2+浓度偏高除了受喀斯特地貌的影响外,还有部分人为因素,游鱼河、蔡冲沟、白岩河是煤矿废水流入阿哈水库的主要通道,酸性pH及铁、锰等元素也会对水库生态系统造成影响,需常在三处投放大量石灰进行治理(白薇扬等,2007),使得春季游鱼河、蔡冲沟 Ca2+浓度高达140 mg∙L−1左右,这势必会对浮游植物生长造成影响。一方面,Ca2+可与磷形成共沉淀,从而减少水体磷含量(刘冠男等,2011),间接抑制浮游植物生长;另一方面,蓝藻大量繁殖时会增强对Ca2+的吸收利用(史绵红等,2009),使其浓度减小,因此春季浮游植物密度随 Ca2+浓度增加而显著减少(P<0.01)。营养盐也是促进浮游植物生长的重要因素,根据罗宜富等(2017)的研究结果,阿哈水库属于磷限制水体,低N/P可能是内源或外源排放的磷浓度增加造成的,亦可能是蓝藻水华导致氮浓度下降、磷浓度升高的结果(许海等,2011),从而使春季浮游植物密度随 N/P增加呈现下降趋势(P<0.05)。秋季浮游植物密度较低且各点位浮游植物组成无明显差异,主要是整体tw、营养盐浓度偏低导致。此外,采样期间水库大坝开闸放水,水流速率较大,也妨碍了浮游植物的正常生长,使其密度降低。
3.3 浮游植物群落结构与环境因子间的关系
根据RDA可知(图4),影响春季浮游植物群落结构的主要环境因子是TP、pH和Ca2+浓度,而夏季的主要环境因子为TP、N/P和SD。高TP浓度的水体中,蓝藻、绿藻、隐藻更易成为优势种,金藻、硅藻更倾向生存于低TP浓度水体。高浓度Ca2+可抑制沉积物中的磷向水中释放(刘冠男等,2011),间接抑制了蓝藻、绿藻、隐藻的生长,促进了金藻和硅藻的繁殖。研究表明,大多数蓝藻的最适pH在7—9之间,绿藻为7—10(许海等,2009),本研究结果显示,蓝藻、绿藻、隐藻更适于pH较高的水体生长,同时,浮游植物的光合作用也会导致pH进一步升高,从而巩固其生长优势。伪鱼腥藻、水华束丝藻(蓝藻)以及狭形纤维藻(绿藻)数量随N/P的增加而减小,而两头针杆藻和曲壳藻(硅藻)与 N/P呈正相关,这是因为蓝藻的生长环境需要高磷低氮,绿藻也是在高磷水体中更具优势,而硅藻则倾向于高氮低磷的水体(石彭灵,2015;吴雅丽等,2013)。蓝藻可通过伪空胞的组装与破裂、碳水化合物的合成与消耗来调节其在水体中的垂直位置(Wallace et al.,2000;Pinilla,2006),因此SD降低使得蓝藻上浮至水体表层,对中下层其他浮游植物生长造成影响,进而使蓝藻成为夏季密度最大的优势种。秋季水体优势浮游植物组成最丰富,含有蓝藻、绿藻、硅藻、隐藻共 16种浮游植物优势种。蓝藻的大量繁殖会分泌化感物质抑制其他藻类的生长(杨斌等,2007),其最适生长温度为 25—30 ℃(Yamaguchi et al.,2000),秋季水温均值仅为 17 ℃,在一定程度上抑制了蓝藻的生长,为其他藻类提供生存机会。
4 总结
本研究通过对阿哈水库3个季节的监测表明,春、夏季均为藻华暴发的高风险时期,浮游植物密度在 3.95—525.35×104cells∙L−1之间,采样期间浮游植物数量以蓝藻、硅藻、绿藻、隐藻居多,丝状蓝藻(如伪鱼腥藻、水华束丝藻)为水库主要优势种。各季节浮游植物与环境因子之间存在相互作用,春季与SD、DO、pH、Ca2+浓度和N/P关系密切,夏季仅与SD相关,秋季受水温、营养盐浓度以及水流速率影响较大。其中,pH、Ca2+和TP是影响春季浮游植物群落结构的主要环境因子,而夏季的主要环境因子为SD、TP和N/P,秋季受低温影响较大。春季Ca2+浓度的变化对浮游植物密度及群落结构均有显著影响。
