基于ISSR标记的火龙果种质资源遗传多样性分析
2021-02-22武志江黄凤珠黄黎芳陆贵锋梁桂东李祯英刘朝安邓海燕
武志江 黄凤珠 黄黎芳 陆贵锋 梁桂东 李祯英 刘朝安 邓海燕


摘 要:为明确火龙果种质资源的遗传多样性水平和亲缘关系,以69份火龙果核心种质为研究对象,采用ISSR分子标记进行火龙果遗传分析。结果表明:13条ISSR引物共检测到247个位点,其中多态性位点数为232个,多态性条带百分比(PPB)为93.93%,平均多态性信息含量(PIC)为0.9189;平均观测等位基因数(Na)为1.9084,有效等位基因数(Ne)为1.4606,期望杂合度(He)为0.2750,Nei氏基因多样性(H)为0.2616,香农信息指数(I)为0.4190。火龙果种质资源的遗传距离为0.0707~0.6741。基于遗传距离聚类分析结果,火龙果种质资源分为6大类群,第Ⅰ类群为红皮白肉类型,包括25份种质;第Ⅱ类群有19份种质,全部为红肉或紫红肉类型,包括红皮、青皮、橘黄皮类型;第Ⅲ类群包含类型较杂,包括粉肉、双色类型和水晶系列品种;第Ⅳ大类为黄皮白肉类型;第Ⅴ类和第Ⅵ类群均仅有一个种质,单独自聚一类。研究结果为火龙果种质资源的分类、保存与利用、遗传进化等研究及杂交组合选配提供理论指导和技术支持。
关键词:火龙果;ISSR;遗传多样性;亲缘关系;种质资源
中图分类号:S667.9 文献标识码:A
Abstract: In order to clarify the level of genetic diversity and relationship of pitaya germplasm resources, 69 core collections of pitaya were used as the research materials, and ISSR molecular markers were used to analyze the genetic characterizaiton of pitaya. The results showed that a total of 247 loci were detected by 13 ISSR primers, of which 232 loci were polymorphic. The percentage of polymorphic bands (PPB) was 93.93%, and the average polymorphism information content (PIC) was 0.9189. The average observed number of alleles (Na), effective number of alleles (Ne), expected heterozygosity (He), Nei's gene diversity (H) and Shannon information index (I) was 1.9084, 1.4606, 0.2750, 0.2616 and 0.4190, respectively. The genetic distance varied from 0.0707 to 0.6741. Six groups were distinguished by truncating the dendrogram. Grop I contained 25 accessions, which were white pulp genotypes. Group II contained 19 red (purple red) pulp accessions, including red, green, and orange peel genotypes. Group III contained miscellaneous genotypes, including pink pulp, two-color type and crystal series varieties. Both group V and group VI had only one accession and were self-clustered, respectively. The results would provide theoretical guidance and technical support for the classification, conservation and utilization, genetic evolution and hybrid combination selection of pitaya germplasm resources.
Keywords: pitaya; ISSR; genetic diversity; genetic relationship; genetic resources
DOI: 10.3969/j.issn.1000-2561.2021.01.007
火龍果(pitaya, pitahaya)是世界上一种独特的保健型热带水果,属双子叶植物纲石竹目仙人掌科(Cactaceae)量天尺属(Hylocereus)或蛇鞭柱属(Selenicereus)多年生攀缘植物,兼具热带沙漠和雨林植物特性,起源于热带中南美洲地区,后传入越南、泰国等东南亚国家和中国台湾[1]。火龙果集水果、花卉、蔬菜、保健、医药于一体[2],具有多批次开花结果习性,自然季花果期长达半年,其产能潜力和经济效益高,市场容量大。近年来,我国火龙果产业规模不断壮大,据2019年统计数据,全国火龙果种植面积已发展到5万余公顷,主要集中在南方地区,包括广西、广东、海南、贵州、云南、福建、台湾等热带、南亚热带地区。目前火龙果生产上的栽培品种大多从中国台湾、越南引进,以红皮白肉和红皮红肉2种类型为主,品种结构单一且种质资源遗传基础狭窄[3-4],因此需要加强对火龙果种质资源的收集保护、挖掘利用及良种选育等工作。然而,火龙果在种质资源引进和发展过程中存在品种名称混乱、来源记载不详等问题,同时现有种质资源的遗传背景不清,亲缘关系不明,挖掘利用力度不够,严重阻碍了火龙果新品种的选育进度。因此,为了加强火龙果种质资源的管理、提高研究和利用效率,有必要针对火龙果种质资源开展遗传多样性分析,有利于更有效地挖掘和利用优异种质资源,提高育种效率。
ISSR(Inter Simple Sequence Repeat)又称简单重复序列间扩增,Zietkiewicz等[5]开发的一种基于微卫星序列的分子标记,具有操作简单、成本低、多态性丰富、稳定性较高等特点,已广泛用于遗传多样性和居群遗传机构、遗传指纹图谱或分子身份证构建等研究[6]。目前在火龙果种质资源的遗传分析上已有不少文献报道,而不同的研究者其保存的种质资源存在差异,所采用的分子标记类型也不一,其中包括RAPD[7]、AFLP[8]、ISSR[9]、SSR[10]等。本课题组早在2012年就已开始收集火龙果种质资源,目前已收集保存的火龙果种质资源达200余份,并于2016年7月被认定为“农业农村部南宁火龙果种质资源圃”,成为广西首批农业农村部认定授牌的果树种质资源圃,是我国火龙果种质资源保存和研究的重要基地。
本研究选取农业农村部南宁火龙果种质资源圃中69份核心种质作为研究对象,采用ISSR分子标记技术对其进行遗传多样性评估,以期阐明种质资源之间的亲缘关系,更全面地了解火龙果种质资源的遗传背景,为火龙果种质资源的分类、保存与利用、遗传进化等研究及杂交组合的亲本选配提供理论指导和技术支持。
1 材料与方法
1.1 材料
供试材料均来源于农业农村部南宁火龙果种质资源圃,基于前期表型性状鉴定,从圃中篩选出69份核心种质(表1)作为研究对象,于2019年4月采取健康嫩茎的茎尖作为供试组织材料,采后于–76 ℃冰箱中保存备用。
1.2 方法
1.2.1 基因组DNA的提取 采用改良CTAB法,每个火龙果嫩茎组织样品称取0.12~0.2 g,放置2 mL EP管中,剪碎,加入1 mL CTAB,组织研磨仪研磨;65 ℃水浴60 min,其间振荡混匀2~3次;12 000 r/min离心5 min,吸取上清转入新的2 mL离心管中,加入300 μL水饱和酚,300 μL氯仿剧烈震荡混匀,室温放置5~10 min;12 000 r/min离心10 min,吸取上清,重复步骤3操作1次;上清液用等体积氯仿抽提1次;12 000 r/min离心10 min,吸取上清,加入1/10 3 mol/L醋酸钠和0.8倍体积的异丙醇沉淀DNA;12 000 r/min离心10 min,沉淀用70%乙醇洗2遍,晾干,用200~500 μL TE溶解DNA。
1.2.2 ISSR-PCR扩增 PCR扩增所用的100条ISSR引物来源于哥伦比亚大学(University of British Columbia, UBC)公布的第9套ISSR引物(801-900),由生工生物工程(上海)股份有限公司合成,用于多态性引物筛选。ISSR-PCR反应体系为20 μL,包含2×Taq PCR Master Mix 10 μL、10 μmol/L引物1 μL、模板DNA 10 ng。PCR反应程序为:94 ℃预变性5 min;94 ℃变性30 s,(Tm–5)℃(根据引物而定)退火45 s,72 ℃延伸90 s,共37个循环,循环结束后4 ℃保存。PCR扩增产物用2.0%琼脂糖凝胶电泳检测,以5 V/cm的电压电泳40 min。电泳后用JS-1075x型凝胶成像系统观察结果,拍照保存。
1.3 数据分析
根据凝胶中条带迁移的位置和有无进行统计,将清晰度好的条带赋值“1”,无条带赋值“0”,构建原始数据矩阵,统计每条引物扩增条带数和多态性条带数,计算多态性条带百分比(PPB)。采用GenAIEx 6.5和POPGENE 1.32进行数据统计分析,计算观测等位基因数(Na),有效等位基因数(Ne),期望杂合度(He),香农信息指数(I)和Nei氏基因多样性(H)等遗传多样性参数和。多态性信息含量(PIC)的计算参考Milbourne等[11]的方法进行,计算公式如下。各参数间的相关性分析通过IBM SPSS Statistics 22.0软件进行计算。将0/1代码序列转化为A/C碱基序列,利用MEGA 10.1.6软件计算遗传距离(genetic distance),采用UPGMA(Unwired pair-Group method with Arithmetical Averages)方法进行聚类分析,构建系统发育树。Bootstrap的初始值设为1000以检验发育树分支的可信度。
其中,Pij是第i个标记的第j个等位基因的频率。
2 结果与分析
2.1 ISSR引物的多态性分析
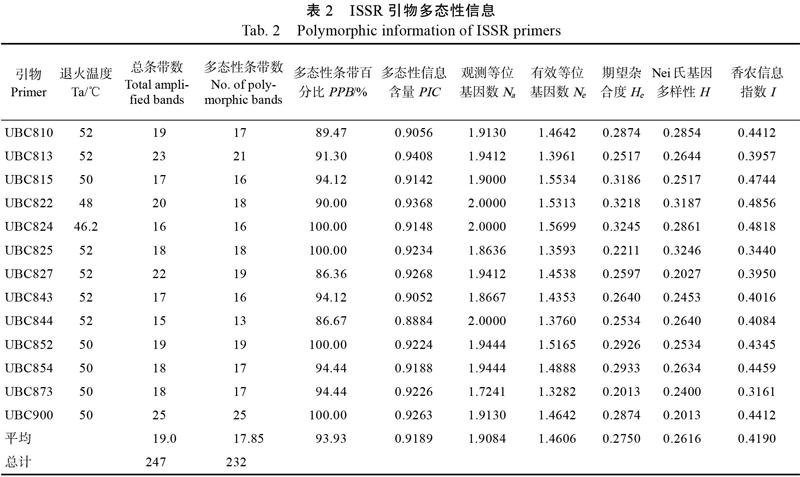
从100条ISSR通用引物中筛选到多态性丰富、重复性好并且能扩增条带清晰的引物13条(表2),分别对69份火龙果种质进行多态性检测,扩增出多个位点且条带清晰,不同种质间有很多特异性条带(图1是引物UBC810的PCR扩增产物电泳图)。13条引物共检测到247个位点,其中多态性位点数为232个,多态性条带百分比(PPB)为93.93%。引物的扩增位点数为15~25,平均每个引物扩增19个位点,其中扩增位点最多的是UBC900,最少的是UBC844。引物的多态性信息含量(PIC)为0.8884~0.9408,平均PIC为0.9189,其中UBC813最大,UBC844最小(表2)。
2.2 种群遗传多样性分析
69份火龙果种质资源的平均观测等位基因数(Na)为1.9084,有效等位基因数(Ne)为1.4606,期望杂合度(He)为0.2750,Nei氏基因多样性(H)为0.2616,香农信息指数(I)为0.4190(表2),这表明火龙果种质资源具有较丰富的遗传多样性。另外,为确定PPB和PIC是否适合用作评估火龙果种群遗传多样性的参数,对各参数间的相关性进行显著性分析,结果显示,PPB和PIC与Na、Ne、He、H、I的相关性均不显著(表3),因此PPB和PIC不适合评估火龙果种群遗传多样性。
2.3 种质间遗传距离分析
69份火龙果种质资源之间的遗传距离为0.0707~0.6741,呈正态分布,平均遗传距离为0.3605,其中遗传距离在0.24~0.48之间的居多,所占频率为86.02%(图2);遗传距离大于0.64的频率仅为0.09%(图2)。这表明大部分的种质资源遗传距离分布较为集中且距离较小,亲缘关系较近。另外,遺传距离小于0.08的种质可能来源于同一亲本或为同一品种的繁殖材料。
2.4 聚类分析
采用UPGMA方法进一步对69份火龙果进行聚类分析(图3)。由图3可知,将69份种质资源分为6大类群,第Ⅰ大类为红皮白肉类型,包括25份种质,属于量天尺属;第Ⅱ大类有19份种质,全部为红肉或紫红肉类型,包括红皮、青皮、橘黄皮类型,属于量天尺属;第Ⅲ大类包含类型较杂,进一步聚类可分为粉肉、双色类型和水晶系列品种,水晶系列又可聚类分为白水晶和红水晶两类,均属于量天尺属,这一类群处于红肉和白肉类型之间,可能为二者杂交后代;第Ⅳ大类为黄皮白肉类型,包括燕窝果(60)、黄龙果(21)、上海黄龙(36)等,属于蛇鞭柱属或量天尺属;第Ⅴ类仅有一个种质黑龙(2),单独聚为一类,属于蛇鞭柱属的一个种。第Ⅵ类也仅有一个种质糖龙(63),此种质来自美国,其生物学特性和果实性状均比较特别,可能属于量天尺属。
3 讨论
本研究采用ISSR分子标记对69份国内外收集的核心火龙果种质资源的遗传多样性及亲缘关系进行评估分析。本研究从100条哥伦比亚大学设计的ISSR通用引物中筛选出多态性丰富、扩增条带清晰的引物13条,其中UBC810、UBC825和UBC873是前人在火龙果遗传分析上应用最多的ISSR引物[9, 12-14],而UBC822、UBC843、UBC852和UBC854是本研究新筛选出来的ISSR引物。另外,本研究筛选出的ISSR引物平均每个扩增19个位点,与其他火龙果研究数据相比扩增的位点相对较多,这可能与火龙果种质资源不同或退火温度存在差异有关。依据PIC值大小与基因多态性高低的相关性[15],本研究所用的ISSR引物的PIC值均高于0.5,属于高度多态性位点,这与Tao等[9]基于表型和ISSR评估火龙果遗传多样性的结果一致。本研究中69份火龙果种质资源的平均观测等位基因数(Na)为1.9084,有效等位基因数(Ne)为1.4606,期望杂合度(He)为0.2750,Nei氏基因多样性(H)为0.2616,香农信息指数(I)为0.4190,此结果与具有较高遗传多样性的野生桃儿七的ISSR参数大小相近[16],这表明火龙果种质资源具有较丰富的遗传多样性。另外,为确定PPB和PIC是否适合用作评估火龙果种群遗传多样性的参数,对各参数间的相关性进行了显著性分析,结果显示PPB和PIC与Na、Ne、He、H、I的相关性均不显著,因此PPB和PIC不适合评估火龙果种质资源的遗传多样性,本结果与Pan等[10]有关火龙果SSR的文献报道不尽相同,他们发现PIC可用于评估其遗传多样性,而PPB不能,这一差异可能与所采用的分子标记类型不同有关。
基于ISSR聚类分析,本研究将火龙果种质资源分为6大类群,基本符合植物分类学结果。第Ⅰ类群(红皮白肉类型,25份)、第Ⅱ类群(红皮红肉或紫红肉类型,19份)和第Ⅳ类群(黄皮类型,3份)的聚类结果与其他研究者们[8-10, 12-14, 17-18]基于分子标记对火龙果的研究结果完全一致,基于果肉颜色分为白肉和红肉类型,基于果皮颜色分为红皮和黄皮类型。
在第Ⅱ类群中有2个主栽商业化品种‘软枝大红(55)和‘金都1号(52)聚在一起,表明这2个品种的亲缘关系较近,且形态学鉴定其植物学特性和开花结果习性均非常接近,推断可能来源于同一亲本或同一品种的不同繁殖材料。另外,红肉类型中的‘青皮(62)和‘橘黄皮(61)红肉火龙果与其他红皮红肉火龙果遗传距离较远,且单独聚为一小类,这一结果与基于表型性状差异进行植物学分类地位相一致,尤其‘青龙的花色为红色,花瓣细长,与其他火龙果的花外形非常不同。据Le Bellec等[19]报道‘青龙(62)的拉丁学名为Hylocereus stenopterus,而据果实性状的差异‘橘龙(61)应归类为量天尺属(Hylocereus)的另外一个种。
第Ⅳ类群的黄皮火龙果均为四倍体类型(已鉴定,数据未发表),其表型兼具蛇鞭柱属和量天尺属的特征,已有文献报道其为蛇鞭柱属和量天尺属自然杂交形成的异源四倍体[20],目前大部分火龙果专家将其归类为量天尺属,但以色列火龙果专家则认为根据其表型更应该将其归类为蛇鞭柱属。
第Ⅲ类群包含的类型较杂,这一类群处于红肉和白肉类型之间,其中粉肉品种(34、37、28)和双色品种(23、41、31、40)可能为红肉和白肉的属内种间杂交选育种或芽变品种。另外水晶系列的品种也聚为此类群,其中‘白水晶和‘红水晶品种各自聚为一类,说明这2个品种亲缘关系既较近又存在一定的遗传距离。从植物学形态来看,水晶类种质在茎蔓、花和果实上与普通火龙果存在明显差异,其花果带少量刺、花细长,而成熟茎蔓近圆柱型,兼具量天尺属和蛇鞭柱属的特征;从聚类分析结果看,整体上处于蛇鞭柱属和量天尺属之间,而且单独成类,与红白肉种间杂种的聚类结果颇为相似,推断其为二倍体量天尺属和二倍体蛇鞭柱属的属间杂交后代,此结论进一步支持了黄凤珠等[21]的观点。
第Ⅴ类和第Ⅵ类群均仅有一个种质,单独自聚一类,分别为‘黑龙(2)和‘美国糖龙(63)。‘黑龙为二倍体火龙果(已鉴定,数据未发表),与黄皮蛇鞭柱属的遗传距离较小,亲缘关系较近。蛇鞭柱属火龙果原产于中南美洲带地区的肉质茎灌木,与量天尺属形态特征差异非常明显,蛇鞭柱属的枝条一般呈近圆柱形或多棱柱形,比量天尺属的枝条更为纤长,果皮长有大量的刺和刚毛[20]。因此,综合考虑植物形态和聚类分析结果,‘黑龙应属于蛇鞭柱属。‘美国糖龙为四倍体火龙果(已鉴定,数据未发表),是新引进的一个国外稀有品种,其植物学形态特征与蛇鞭柱属很接近,果皮呈暗红色、无刺,果肉呈红色,与其他红肉火龙果外形差异明显,其他生物学特性有待进一步观察。
据相关文献报道,火龙果量天尺属包括17个种,蛇鞭柱属有20个种[18, 22]。国内引进的大部分火龙果以商业化主栽品种为主,因此资源圃从国内收集保存的火龙果种类非常少,本研究中85%以上的种质资源的遗传距离在0.24~0.48之间,这表明大部分的种质资源遗传距离分布较为集中且距离较小,亲缘关系相对较近,遗传基础比较狭窄。这与火龙果原生种的获取途径有限以及国外对本国种质资源的保护限制有关。因此,对于国内火龙果育种者来说,国外原生种资源的引进和自主知识产权品种的选育是未来火龙果育种工作的重中之重。
参考文献
田新民, 李洪立, 何 云, 等. 火龙果研究现状[J]. 北方园艺, 2015, 39(18): 188-193.
Ortiz-Hernández Y D, Carrillo-Salazar J A. Pitahaya (Hylocereus spp.): A short review[J]. Comunicata Scientiae, 2012, 3(4): 220-237.
董美超, 岳建强, 李进学, 等. 火龙果育种研究进展[J]. 热带农业科学, 2013, 33(5): 56-59.
申世辉, 马玉华, 蔡永强. 火龙果研究进展[J]. 中国热带农业, 2015(1): 48-52.
Zietkiewicz E, Rafalski A, Labuda D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification[J]. Genomics, 1994, 20(2): 176-183.
朱岩芳, 祝水金, 李永平, 等. ISSR分子标记技术在植物种质资源研究中的应用[J]. 种子, 2010, 29(2): 55-59.
Junqueira K P, Faleiro F G, Junqveira N T V, et al. Genetic diversity of native pitaya native from brazilian savannas with basis on RAPD markers[J]. Revista Brasileira de Fruticultura, 2010, 32(3): 819-824.
Pagliaccia D, Vidalakis G, Douhan G W, et al. Genetic characterization of pitahaya accessions based on amplified fragment length polymorphism analysis[J]. HortScience, 2015, 50(3): 332-336.
Tao J, Qiao G, Wen X P, et al. Characterization of genetic relationship of dragon fruit accessions (Hylocereus spp.) by morphological traits and ISSR markers[J]. Scientia Horticulturae, 2014, 170: 82-88.
Pan L M, Fu J X, Zhang R, et al. Genetic diversity among germplasms of Pitaya based on SSR markers[J]. Scientia Horticulturae, 2017, 225: 171-176.
Milbourne D, Meyer R, Bradshaw J E, et al. Comparison of PCR-based marker systems for the analysis of genetic relationships in cultivated potato[J]. Molecular Breeding, 1997, 3(2): 127-136.
袁亞芳, 陈明贤, 陈清西, 等. 福建地区火龙果种质资源调查及ISSR分析[J]. 中国农学通报, 2013, 29(34): 216-220.
王 彬, 郑 伟, 宋 莎, 等. 紫红龙火龙果及其大果型芽变系的ISSR分析[J]. 西南农业学报, 2013, 26(4): 1614-1617.
张冰雪, 范付华, 乔 光, 等. 贵州地方火龙果芽变种质DNA指纹图谱及遗传多样性的ISSR分析[J]. 果树学报, 2013, 30(4): 573-577.
Cho Y G, Ishii T, Temnykh S, et al. Diversity of microsatellites derived from genomic libraries and GenBank sequences in rice (Oryza sativa L.)[J]. Theoretical and Applied Genetics, 2000, 100(5): 713-722.
沈 奇, 张 盾, 臧春鑫, 等. 基于ISSR分子标记的野生桃儿七遗传多样性研究[J]. 植物遗传资源学报, 2019, 20(1): 129-136.
杨仕美, 乔 光, 毛永亚, 等. 基于火龙果转录组测序的SSR标记开发及种质亲缘关系分析[J]. 分子植物育种, 2018, 16(24): 8096-8110.
Telzur N, Abbo S, Bar-Zvi D, et al. Clone identification and genetic relationship among vine cacti from the genera Hylocereus and Selenicereus based on RAPD analysis[J]. Scientia Horticulturae, 2003, 100(1): 279-289.
Le Bellec F, Vaillant F, Imbert E. Pitahaya (Hylocereus spp.): a new fruit crop, a market with a future[J]. Fruits, 2006, 61(4): 237-250.
Tel-Zur N, Abbo S, Bar-Zvi D, et al. Genetic relationships among Hylocereus and Selenicereus vine cacti (Cactaceae): evidence from hybridization and cytological studies[J]. Annals of Botany, 2004, 94(4): 527-534.
黄凤珠, 陆贵锋, 黄黎芳, 等. 火龙果种质资源收集保存与初步评价[J]. 西南农业学报, 2016, 29(4): 920-924.
Ortiz-Hernández Y D, Livera-Mu?oz M, Carrillo-Salazar J A, et al. Agronomical, physiological, and cultural contributions of pitahaya (Hylocereus spp.) in Mexico[J]. Israel Journal of Plant Sciences, 2013, 60: 359-370.
责任编辑:黄东杰
