低温胁迫及恢复对杏树品种雌蕊生长发育的影响1)
2021-01-14于丹刘肖娟毕泉鑫赵阳句娇于海燕王利兵
于丹 刘肖娟 毕泉鑫 赵阳 句娇 于海燕 王利兵
(国家林业和草原局林木培育重点实验室(中国林业科学研究院林业研究所),北京,100091)
杏(PrunusarmeniacaLinn.),蔷薇科(Rosaceae)杏属(Armeniaca),具有耐干旱、耐瘠薄、冬季耐低温、抗盐碱等特性,其分布范围较广,主要集中在华北、华东以及西北等地区,是我国北方地区重要的栽培树种之一[1-2]。杏树具有巨大的开发价值,杏果和杏仁中含有丰富的维生素、矿质元素和胡萝卜素等,具有止咳平喘、预防和治疗癌症等功效[1-3],杏仁油富含油酸(63.39%)和亚油酸(28.79%),不饱和脂肪酸占总脂肪酸含量的90%以上,是一种优质食用油[4-6];杏木可作家具和工艺品;杏壳是制作活性炭的原料。但是,晚霜危害在西北地区普遍存在,而且由于杏树开花时间较早,其花、芽等生殖器官极易受到晚霜的危害,严重影响其生长发育及产量,晚霜已成为限制杏树生产和发展的重要环境因素[3,7]。因此,明确杏树花期的抗寒机理、选育抗冻品种对杏树的发展有重要意义。
杏树花期的抗寒性与细胞膜的损伤程度、抗氧化酶活性以及渗透调节物质等密切相关[8-10]。然而,有关杏树雌蕊对低温胁迫和恢复后的响应的比较研究较少,而且关于雌蕊的可承受的极限温度以及雌蕊的解剖结构的研究也较少。文中通过对抗寒品种‘围选1号’和冷敏感品种‘龙王帽’雌蕊的表型、解剖结构以及生理指标的观测,探究杏树花期对低温胁迫及恢复过程的响应,为杏树花期晚霜响应机理的研究提供一定的理论基础。
1 材料与方法
1.1 试验材料
试验材料来自陕西省榆林市李杏优良品种种质资源圃。随机采取抗寒性不同的2个杏树品种‘围选1号’和‘龙王帽’东、南、西、北方向的1年生枝条各30枝,混合并带回温室水培。试验采用单株小区,重复3次。
1.2 低温处理
将杏树花枝在温室(20 ℃)水培至盛花期,将枝条分批放入LED人工气候箱中,试验温度设置为0、-2、-4、-6、-8 ℃共5个温度。以0.5 h降温10 ℃的速度从20 ℃降至2 ℃,再以3 ℃·h-1的降温速度降至所需温度并在所需温度处维持1 h,并将枝条放在室温下恢复3 h。随机采取盛花期的雌蕊用于相对电导率的测定,并利用Logistic方程计算出杏树雌蕊的低温半致死温度(LT50)。
根据上述计算的‘围选1号’和‘龙王帽’雌蕊的LT50设置杏树枝条处理的温度梯度为-1、-2、-3、-4、-6 ℃,降温速度与上述一致。低温处理完成后,将试验材料分为2批,一批直接在低温环境下采样后分别放入FAA固定液(体积分数38%甲醛+醋酸+体积分数70%酒精)和液氮中,用于解剖结构和生理指标的观测;另一批在室温恢复3 h后再取样并分别放入FAA固定液和液氮中,用于褐化率的统计和生理指标的测定。
1.3 测定方法
1.3.1 相对电导率的测定
称取0.1 g新鲜雌蕊放入装有10 mL去离子水的试管中,将试管放入真空干燥器中,反复抽气4次,每次20 min,测定其初始电导,然后将其置入沸水中水浴30 min,冷却至室温后测其最终电导。计算相对电导率。相对电导率=(初始电导-去离子水电导)/(最终电导-去离子水电导)。
将杏树雌蕊的相对电导率拟合成Logistic方程:
Y=K/(1+ae-bx)。
式中:Y为相对电导率;K为极限相对电导率;a,b均为方程参数;x为温度。
求Logistic方程的二阶导数,并令其为0,可得到方程的拐点温度LT50。
LT50=(lna)/b。
1.3.2 雌蕊表型及褐化率的观测
组织褐变是植物受到冻害最直接的形态表现[11-12]。低温胁迫后,在室温下静置3 h后,随机选取100个雌蕊,观察其褐变程度并计算其褐化率,重复3次。
1.3.3 解剖结构的观察
采用常规石蜡切片法[13]。将FAA固定液中的雌蕊依次进行一系列浓度的酒精脱水,二甲苯透明,浸蜡,包埋,连续切片,粘片,番红-固绿染色,封片。切片厚度为10 μm。
1.3.4 生理指标的测定
丙二醛(MDA)质量摩尔浓度、过氧化氢酶(CAT)和过氧化物酶(POD)活性测定分别采用巴比妥酸法、过氧化氢法和愈创木酚法[14-15]。MDA质量摩尔浓度,CAT和POD活性的测定所用的试剂盒均由中国北京索莱宝科技有限公司提供。
1.4 数据分析
利用SPSS 21.00对数据进行单因素方差分析以及Duncan多重比较分析。
2 结果与分析
2.1 低温胁迫下杏树品种雌蕊相对电导率的变化以及LT50的确定
2个杏树品种雌蕊相对电导率的变化如表1所示。随着温度的降低‘龙王帽’和‘围选1号’的相对电导率均呈现不断升高的趋势。‘龙王帽’和‘围选1号’相对电导率的增长速率有所差异。‘龙王帽’相对电导率在0~-2、-2~-4、-4~-6、-6~-8 ℃的增长分别为1100%、117%、10%和33%,其中,在-4~-8 ℃时相对电导率有轻微升高直至趋于平缓;‘围选1号’的相对电导率在0~-2、-2~-4、-4~-6、-6~-8 ℃的增长分别为992%、63%、52%和12%,其中,相对电导率在-6~-8 ℃趋于平缓。另外,在相同胁迫条件下,‘围选1号’的相对电导率始终低于‘龙王帽’。

表1 低温胁迫下杏树相对电导率的变化
利用Logistic方程对‘龙王帽’和‘围选1号’的相对电导率进行拟合。由表2可知,‘龙王帽’和‘围选1号’的拟合度R2分别为0.999和0.984,均达到显著水平,表明拟合结果可靠、精确度高。‘龙王帽’和‘围选1号’的LT50分别为-2.30 ℃和-3.44 ℃,表明相较于‘龙王帽’,‘围选1号’雌蕊的抗寒性更强。

表2 杏树雌蕊的Logistic方程以及低温半致死温度的确定
2.2 低温胁迫和恢复处理对杏树雌蕊表型及解剖结构的影响
在冻害下,植物组织解冻后会释放一些酚类化合物,导致受损组织变成褐色[11],因此,杏树雌蕊在室温恢复3 h后对其褐化率进行了观测。随温度的降低‘龙王帽’和‘围选1号’的褐变程度加剧,褐化率也呈逐渐上升的趋势(图1,表3)。在-1 ℃时,‘龙王帽’和‘围选1号’雌蕊均未发生明显的变化,表明雌蕊未受到冻害。在-2~-4 ℃时,‘龙王帽’的褐化率始终高于‘围选1号’,其中,-3 ℃时,‘龙王帽’和‘围选1号’的褐化率急剧升高,分别是-2 ℃时的1.94倍和3.50倍。在-6 ℃时,‘龙王帽’和‘围选1号’雌蕊的褐化率均达到100%。

图1 低温胁迫下杏树雌蕊的表型变化

表3 低温胁迫下杏树雌蕊的褐化率
采用常规石蜡切片法,对‘龙王帽’和‘围选1号’的雌蕊的连续切片进行观察(图2)。在低温胁迫下,‘龙王帽’雌蕊在20、-1 ℃和-2 ℃均未发现明显变化,在-3 ℃时,‘龙王帽’雌蕊的子房壁出现小裂隙或空腔,-4 ℃和-6 ℃时,其子房壁和胚珠均出现较大裂隙和空腔;‘围选1号’雌蕊在20、-1、-2 ℃和-3 ℃均未发现明显变化,在-4 ℃和-6 ℃时子房壁和胚珠均出现裂隙和空腔。在恢复过程中,‘龙王帽’在-3 ℃的子房壁有小空腔,在-4 ℃和-6 ℃的子房壁、胚珠均出现较大裂隙;‘围选1号’的子房壁和胚珠在-4 ℃出现轻微裂隙,在-6 ℃时子房壁和胚珠均出现较大的裂隙和空腔。另外,在-4 ℃时,低温胁迫和恢复过程中‘围选1号’的裂隙总是比‘龙王帽’小。
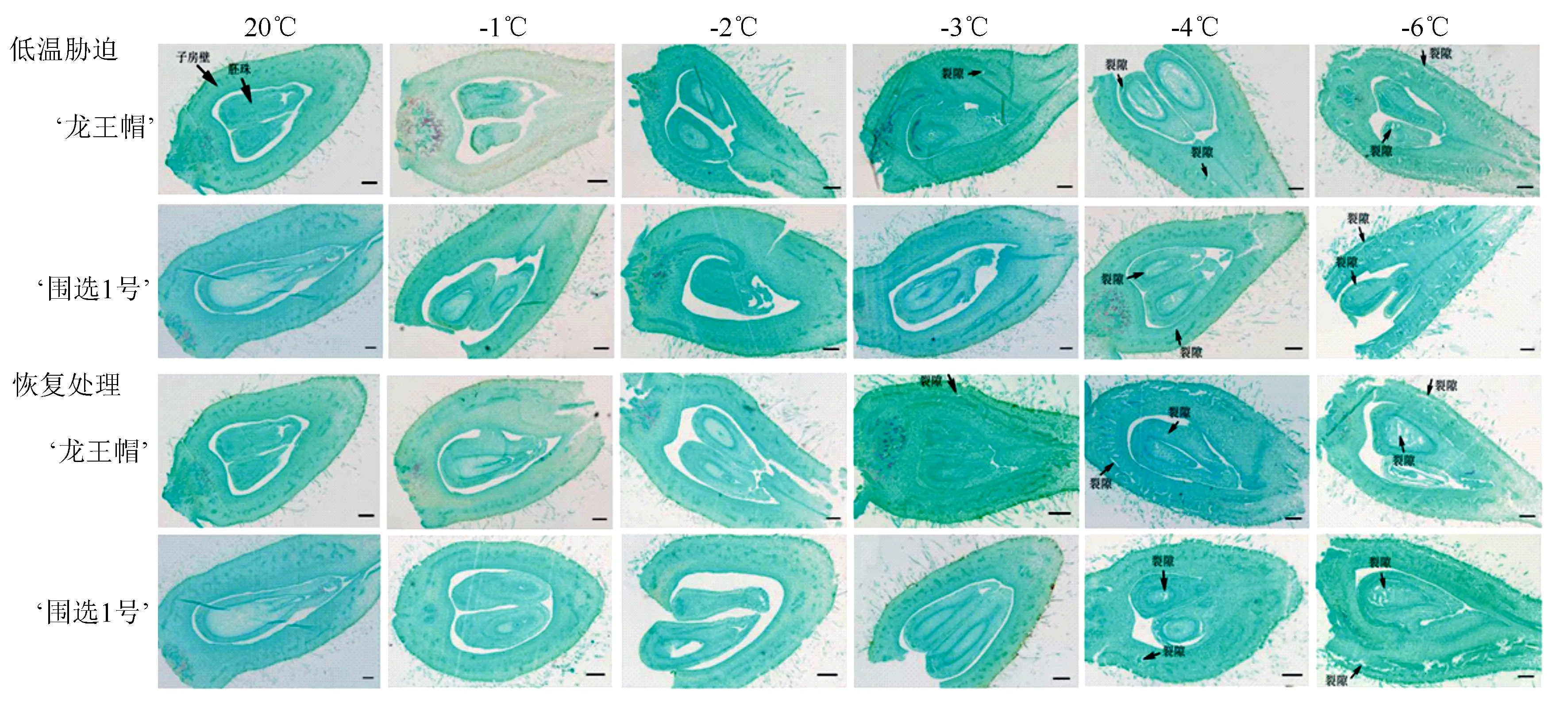
图2 低温胁迫和恢复处理对杏树雌蕊解剖结构的影响
2.3 低温胁迫和恢复处理对2个杏树品种雌蕊生理指标的影响
2.3.1 对杏花雌蕊MDA质量摩尔浓度的影响
低温胁迫和恢复处理过程中的MDA质量摩尔浓度变化如表4所示。‘龙王帽’和‘围选1号’雌蕊中的MDA质量摩尔浓度均随温度的降低呈上升的趋势,其中,-6 ℃时‘龙王帽’和‘围选1号’的MDA质量摩尔浓度与对照组(20 ℃)的差异极显著。同一低温胁迫下‘龙王帽’的MDA质量摩尔浓度始终高于‘围选1号’,而且随着温度的降低两者MDA质量摩尔浓度的差距越来越大。‘龙王帽’MDA质量摩尔浓度在-2~-6 ℃的增长幅度较大,而‘围选1号’MDA质量摩尔浓度在-3~-6 ℃的增长幅度较大。
在低温胁迫下,-6 ℃时‘龙王帽’和‘围选1号’的MDA质量摩尔浓度分别为43.77、29.42 nmol·g-1,分别是对照组(20 ℃)的2.28倍和1.43倍。在恢复处理下,-6 ℃时‘龙王帽’和‘围选1号’MDA质量摩尔浓度分别为38.75、26.16 nmol·g-1,分别是对照组(20 ℃)的1.90倍和1.16倍。在低温胁迫和恢复处理下,‘龙王帽’MDA质量摩尔浓度在-3~-6 ℃时极显著高于‘围选1号’。另外,相较于低温胁迫,‘龙王帽’和‘围选1号’恢复处理过程中的MDA质量摩尔浓度均有所降低(表4)。
2.3.2 对杏花雌蕊抗氧化酶活性的影响
不同低温胁迫下,杏树雌蕊抗氧化酶活性(POD和CAT)变化如表4所示。‘龙王帽’和‘围选1号’雌蕊的POD和CAT活性随温度的降低呈‘上升-下降’的趋势,二者POD和CAT活性均在-3 ℃达到最大值,而且在同一温度下‘龙王帽’的POD和CAT活性均低于‘围选1号’的;低温胁迫时的POD和CAT活性总是低于恢复过程中的酶活性。
POD是植物应对胁迫时的重要抗氧化酶[16]。以低温胁迫过程为例,-1~-6 ℃时,‘龙王帽’和‘围选1号’的POD活性与对照组差异极显著。其中,在-3 ℃时,‘龙王帽’和‘围选1号’雌蕊的POD活性达到最大值,分别为491.27、606.18 U·g-1,分别是对照组的1.73倍和2.89倍;此外,在-3~-4 ℃时,‘围选1号’的POD活性极显著高于‘龙王帽’;在-6 ℃时,‘围选1号’的POD活性显著高于‘龙王帽’(表4)。
CAT对H2O2的清除有重要作用[17]。以低温胁迫过程为例,在-1~-6 ℃,‘龙王帽’和‘围选1号’的CAT活性极显著高于对照组。其中,-3 ℃时,‘龙王帽’和‘围选1号’雌蕊的CAT活性分别为441.21、480.72 U·g-1,分别是对照组的2.43倍和2.27倍;此外,在-3 ℃时,‘龙王帽’和‘围选1号’的CAT活性并无显著差异,但在-4~-6 ℃时,‘龙王帽’和‘围选1号’的CAT活性差异极显著(表4)。

表4 低温胁迫和恢复过程中杏树雌蕊生理指标变化
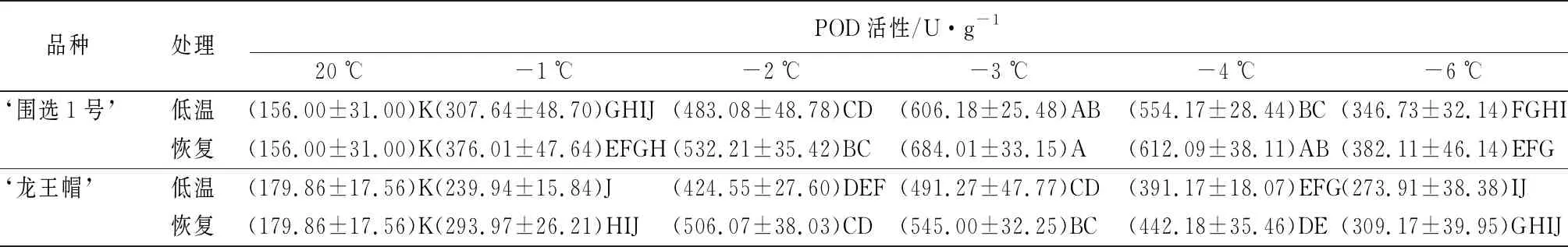
品种处理POD活性/U·g-120℃-1℃-2℃-3℃-4℃-6℃‘围选1号’低温(156.00±31.00)K(307.64±48.70)GHIJ(483.08±48.78)CD(606.18±25.48)AB(554.17±28.44)BC(346.73±32.14)FGHI恢复(156.00±31.00)K(376.01±47.64)EFGH(532.21±35.42)BC(684.01±33.15)A(612.09±38.11)AB(382.11±46.14)EFG‘龙王帽’低温(179.86±17.56)K(239.94±15.84)J(424.55±27.60)DEF(491.27±47.77)CD(391.17±18.07)EFG(273.91±38.38)IJ恢复(179.86±17.56)K(293.97±26.21)HIJ(506.07±38.03)CD(545.00±32.25)BC(442.18±35.46)DE(309.17±39.95)GHIJ

品种处理CAT活性/U·g-120℃-1℃-2℃-3℃-4℃-6℃‘围选1号’低温(147.10±14.14)MN(240.34±30.03)KL(370.67±44.67)EFGH(480.72±46.10)ABC(451.92±30.17)BCD (290.93±33.61)IJK恢复(147.10±14.14)MN(285.20±27.42)IJK(432.18±33.11)CDEF(551.68±40.26)A(511.71±25.82)AB(322.58±22.01)HIJ‘龙王帽’低温(128.50±28.07)N(205.31±32.23)LM(359.75±38.22)FGHI(441.21±35.22)BCDE(345.73±24.10)GHI(204.54±31.78)LM恢复(128.50±28.07)N(250.34±29.30)JKL(418.13±24.48)CDEFG(491.01±28.23)ABC(391.87±30.75)DEFGH(233.50±38.01)KL
3 结论与讨论
本试验以2个抗寒性不同的杏树品种‘龙王帽’和‘围选1号’的雌蕊[7,18]为试验材料,利用LED人工气候箱模拟自然降温过程,重点对低温胁迫和恢复处理过程中杏花雌蕊的表型、解剖结构和生理指标进行了观测。研究发现,随着温度的降低,‘龙王帽’和‘围选1号’的褐变程度加剧且褐化率增加,但-2~-4 ℃时,‘围选1号’的褐化率始终低于‘龙王帽’。该结果与王萍等[7]的研究结果一致。在-6 ℃时,‘围选1号’和‘龙王帽’雌蕊的褐化率均达到100%,表明此时2个杏树品种的雌蕊均受到严重的冻害。
冻害会引起细胞间隙的水结冰,导致原生质脱水,破坏蛋白质结构;随后,原生质体结构也会因冻害而结冰,造成不可逆损伤[3]。本研究中,‘龙王帽’和‘围选1号’雌蕊因冻害产生了一些空腔或裂隙,推测这些空腔或裂隙可能是因细胞失水或死亡导致的;另外,‘龙王帽’在-3 ℃发现裂隙,而‘围选1号’在-4 ℃发现裂隙,相对于‘围选1号’,‘龙王帽’裂隙出现的温度较高,且裂隙较多或较大,这表明相同条件下‘龙王帽’的雌蕊细胞被冻害破坏较严重。彭伟秀等[19]在研究低温胁迫对杏树子房结构的影响中发现,在-3 ℃时,‘优一’和‘一窝蜂’的子房均出现一些裂隙或空腔,这与本研究结果相似。
当植物受到冻害时,细胞膜透性变大,导致电解质外渗,而且植物的相对电导率与植物的抗寒性呈负相关[20-21]。随着温度的降低,‘龙王帽’和‘围选1号’的相对电导率均不断升高,这表明细胞膜的损伤程度随温度的降低而加剧。‘龙王帽’和‘围选1号’雌蕊的相对电导率分别在-2 ℃时增长速率最大,分别是对照组(20 ℃)的11.00倍和9.92倍,说明‘龙王帽’和‘围选1号’的膜系统分别在0~-2 ℃受到严重的低温伤害,但‘围选1号’细胞膜透性受到的伤害较小。-4~-8 ℃时,‘龙王帽’和‘围选1号’增长速率逐渐降低,表明杏树雌蕊细胞膜透性逐渐变大,电解质外渗严重。‘龙王帽’和‘围选1号’雌蕊分别在-4~-8 ℃和-6~-8 ℃时相对电导率变化趋于平缓且均为82%以上,表明此时‘龙王帽’和‘围选1号’的细胞膜透性基本已被完全破坏,细胞死亡。LT50可反映植物可承受的极限低温,LT50越低其抗寒性越强[22]。‘围选1号’的LT50低于‘龙王帽’,表明‘围选1号’可承受的极限温度较低,抗寒性更强。低温胁迫下,植物体内会产生大量的活性氧和自由基,导致膜脂过氧化并积累大量MDA,所以MDA质量摩尔浓度反映了细胞膜损伤的程度[23-25]。本研究中,‘龙王帽’和‘围选1号’雌蕊的MDA质量摩尔浓度呈逐渐上升的趋势,-6 ℃时,MDA质量摩尔浓度极显著高于对照组(20 ℃)。‘龙王帽’雌蕊的MDA质量摩尔浓度始终高于‘围选1号’,表明‘围选1号’细胞膜损伤的程度较小;‘龙王帽’和‘围选1号’在恢复处理过程中的MDA质量摩尔浓度相较于低温胁迫过程中均有所降低,这表明恢复室温后其膜系统的损伤也有所缓解。
POD和CAT是植物体内清除活性氧和自由基的重要抗氧化酶[26-28]。随着胁迫的加剧,‘龙王帽’和‘围选1号’雌蕊的POD和CAT活性呈‘上升-下降’的趋势。这与孟庆瑞等[29]的研究一致。其在-3 ℃达到最大值,这表明‘龙王帽’和‘围选1号’的抗氧化能力在-3 ℃时最强;在-3 ℃之后,二者的酶活性均有不同程度的降低。相较于低温胁迫,恢复处理时POD和CAT活性总是较高,这表明在温度恢复后,植物会产生更多的抗氧化酶尽快清除体内的自由基和活性氧;而在-6 ℃时,低温胁迫和恢复处理中的酶活性相差较小,这表明杏树雌蕊的抗氧化酶系统已遭受到严重的破坏。此外,同一温度下‘龙王帽’的POD和CAT活性均低于‘围选1号’,表明‘围选1号’具有较强的抗氧化能力。总之,相较于‘龙王帽’,‘围选1号’具有较强的抗氧化能力且恢复能力较强,这为解析杏树雌蕊的霜冻响应机理提供一定的基础。
