独行菜抗逆相关转录因子LaDREB密码子偏性与进化分析
2021-01-08王玉州王继莲赵惠新
周 茜,陈 芸,卢 函,王玉州,王继莲,赵惠新
(1.喀什大学生命与地理科学学院/叶尔羌绿洲生态与生物资源研究高校重点实验室,新疆喀什 844000;2.新疆师范大学生命科学学院/新疆特殊环境物种保护与调控生物学实验室/干旱区植物逆境生物学实验室,新疆乌鲁木齐 830054;3.和田师范专科学校生地学院,新疆和田 848000)
【研究意义】密码子是核苷酸序列翻译为氨基酸序列的联通途径,是遗传信息准确传递的关键[1−2]。密码子偏性(condon usage bias,CUB)是由于物种长期的进化,导致编码同一氨基酸的密码子(同义密码子)可能有多种,而使用过程不随机,通常根据环境的变化,密码子使用的适应性也发生相应改变而形成。不同的物种,可依据自身特异的进化方式,从而选择性的偏向使用某一特定密码子,导致形成了该物种特有的密码子使用偏性[1]。特定基因和物种对一种或几种同义密码子的使用偏好,在生物界普遍存在,影响特定基因的碱基相关组成和该基因的表达水平等[3]。最优密码子(最优先被使用的密码子),多出现在表达量较高的基因中[4]。评价密码子使用偏好性的参数主要有有效密码子数(effective number of codons,ENc)、同义密码子相对使用度(relative synonymous codon usage,RSCU)、密码子适应指数(codon adaptation index,CAI)和不同位置GC含量等[3]。进行密码子偏性分析能为特定基因功能预测[5]、分子进化水平[6]、表达调控机制[7]、蛋白质结构[8]等提供重要参考。【前人研究进展】DREB转录因子即干旱应答元件结合蛋白质(dehydration responsive element binding protein),是AP2/EREBP转录因子蛋白家族中EREBP亚家族中的一员,能特异性的结合DRE顺式作用元件;激活所调控下游基因的转录表达[9]。不仅在植物体生长发育和形态建成等生理活动的调控中发挥重要作用,而且与植物次生代谢、抗干旱、高盐和低温等生长抗逆反应紧密相关[10−11]。【本研究切入点】目前,已从拟南芥[12]、水稻[13]、大豆[14]、沙漠苔藓[15]和香蕉[16]等植物中分离到DREB类基因。近年密码子使用偏性在水稻[17]、拟南芥[18]、葡萄[19]和小麦[20]等中均有报道,还未见独行菜LaDREB密码子偏性的研究。【拟解决的关键问题】本研究根据本实验室已经克隆到的独行菜LaDREB基因进行了预测,对独行菜LaDREB基因密码子使用模式及其基因进化模式进行了分析,为独行菜LaDREB基因的功能和转基因研究提供理论依据。
1 材料与方法
1.1 序列数据来源
独行菜LaDREB克隆于本实验室,序列全长639 bp,ORF长度633 bp,编码210个氨基酸,在NCBI中注册号为MT939484。
玫瑰花、欧洲油菜、毛果杨、荠、草莓、芥菜和菥蓂等26种植物的DREB基因序列从GenBank中获得(表1)。
大肠杆菌、酵母菌、拟南芥和烟草等模式生物的基因组密码子偏好性数据来源于Codon Usage Database。

表1 DREB基因编码区序列Tab.1 Sources of coding sequence of DREB genes
1.2 密码子使用偏性分析
密码子使用频率采用CodonW软件和EMBOSS在线程序中的CHIPS(codon heterozygosity in a protein coding sequence)、CUSP(create a codon usage table)进行分析[21−22]。对独行菜LaDREB基因及其他物种DREB基因的CDS(coding sequence)序列的ENc有效密码子数,密码子第1、2和3位上的GC含量(GC1、GC2和GC3)、密码子各个碱基在第3位上的含量(A3s、G3s、T3s和C3s)、密码子适应指数CAI和同义密码子相对使用度RSCU等密码子偏性的相关参数,进行运算分析。采用CUSP分析基因密码子使用频率。使用Excel和SPSS等软件,将密码子相关参数进行统计,进行ENc绘图、中性绘图和PR2分析(PR2−bias plot analysis)。
1.3 同义密码子使用偏性分析
密码子使用的偏性程度,用RSCU值评价,RSCU值=某一密码子使用频率/其无偏性使用时预期频率。当RSCU值等于1,RSCU值大于l和RSCU值小于l时,分别表明该密码子没有使用偏好,使用频率较高和使用频率较低[23]。基因碱基组成偏性由GC3s的值进行评价[24]。密码子使用频率,能够体现物种间使用密码子的选择差异。比值在0.5~2.0,表示两物种密码子使用偏性较小。比值≤0.5或≥2.0时,表明两物种密码子偏性有较大差异[25]。基因有效密码子数ENc的值,在20~61,偏性越强,值越接近20,值越大,偏性相对较弱且稀有密码子增多[26]。
1.4 DREB基因的聚类分析
使用SPSS19.0进行密码子使用选择性相关参数等进行分析,并针对不同物种DREB基因RSCU的值,把密码子使用选择偏性聚类。将每个基因作为一个对象,用欧氏平方距离表示,采用离差平方和法[27]表示基因间的距离。使用MEGA 7.0邻接法构建27个物种DREB基因的CDS序列进化树。
2 结果与分析
2.1 独行菜LaDREB基因密码子使用偏性分析
2.1.1 独行菜LaDREB基因的GC、ENc和CAI分析 利用CHIPS和CodonW程序,计算得到独行菜LaDREB基因的EN值为50.49c,GC含量为0.456,GC3s值为0.337。ENc值>35,接近61,表明在独行菜LaDREB基因编码时,对密码子选择偏性较弱,表达水平可能偏低(表2)。独行菜LaDREB基因编码区GC含量为45.6%,与AT含量(54.4%)基本相当。独行菜LaDREB基因中密码子的GC、GC1、GC2和GC3值分别为0.456、0.521、0.460和0.384,表明独行菜LaDREB基因密码子第3位碱基偏好使用以AT结尾。独行菜LaDREB基因编码区的密码子适应指数CAI值为0.262,远小于1,进一步表明独行菜LaDREB基因对密码子偏性弱。
2.1.2 独行菜LaDREB基因的RSCU分析 采用EMBOSS对独行菜LaDREB基因的RSCU值进行计算,使用CodonW程序获得独行菜LaDREB基因密码子的使用频率。如表2,在独行菜LaDREB基因中,密码子RSCU值大于l.5的个数为11,RSCU值大于l的个数为24。在使用偏性较强的密码子中,GCU的RSCU值为2.61,GCC的值为0.70,GCG的值为0.00,GCA的值为0.70,表明编码独行菜LaDREB的丙氨酸残基的4种密码子中主要使用GCU,对该密码子有较强的选择偏性。

表2 独行菜LaDREB基因密码子的使用偏性分析Tab.2 Analysis of codon bias of LaDREB gene in Lepidium apetalum
密码子RSCU值>2的有GCU、CGU、AUU和CCA,这4种密码子也具有较高的使用偏性。CUA、AUC、CAC、CCU等10个密码子的RSCU值=0,表明独行菜LaDREB基因不使用这10个密码子。AUG、ACU、ACA、UGG和CGA密码子的RSCU值为1,说明这5个密码子无使用偏性。其余22种密码子的RSCU值<1,使用偏好小于预期值。然而GCU、CGU、AUU和CCA在各自的同义密码子中被优先使用,导致ENc值的偏离61。
2.2 不同物种间DREB基因密码子选用偏好性分析
2.2.1 不同物种CAI和ENc分析 不同物种间DREB基因密码子具有不同的使用选择偏性。如表3,27个物种DREB基因密码子使用偏性存在差异。不同单双子叶植物DREB基因的CAI值范围为0.14~0.261,差异较小,GC值与ENc却有较大差异。27个物种DREB基因的ENc平均值为49.571,均大于35,表明27个物种基因的密码子使用偏性和表达水平有一定差异,且选择使用偏性和表达水平都较低,这与CAI指数验证结果一致。高山离子芥的ENc值=61,表明其密码子使用选择上无偏性。双子叶植物中欧洲白桦、猕猴桃、毛果杨、草莓、胡杨、小叶杨、河北杨、杧果、茄、番木瓜等的DREB基因与独行菜LaDREB基因的密码子使用偏性相关参数相似,ENc值的平均值为49.17,值的范围为37.46~61.00。表明双子叶植物DREB基因在翻译过程中,选择密码子编码氨基酸时偏性不强;但单子叶植物DREB基因的ENc值在53.56~55.50,平均值54.53,高于双子叶植物的ENc值,暗示这些植物DREB基因密码子使用偏性较双子叶弱,其表达量可能较双子叶植物更低。
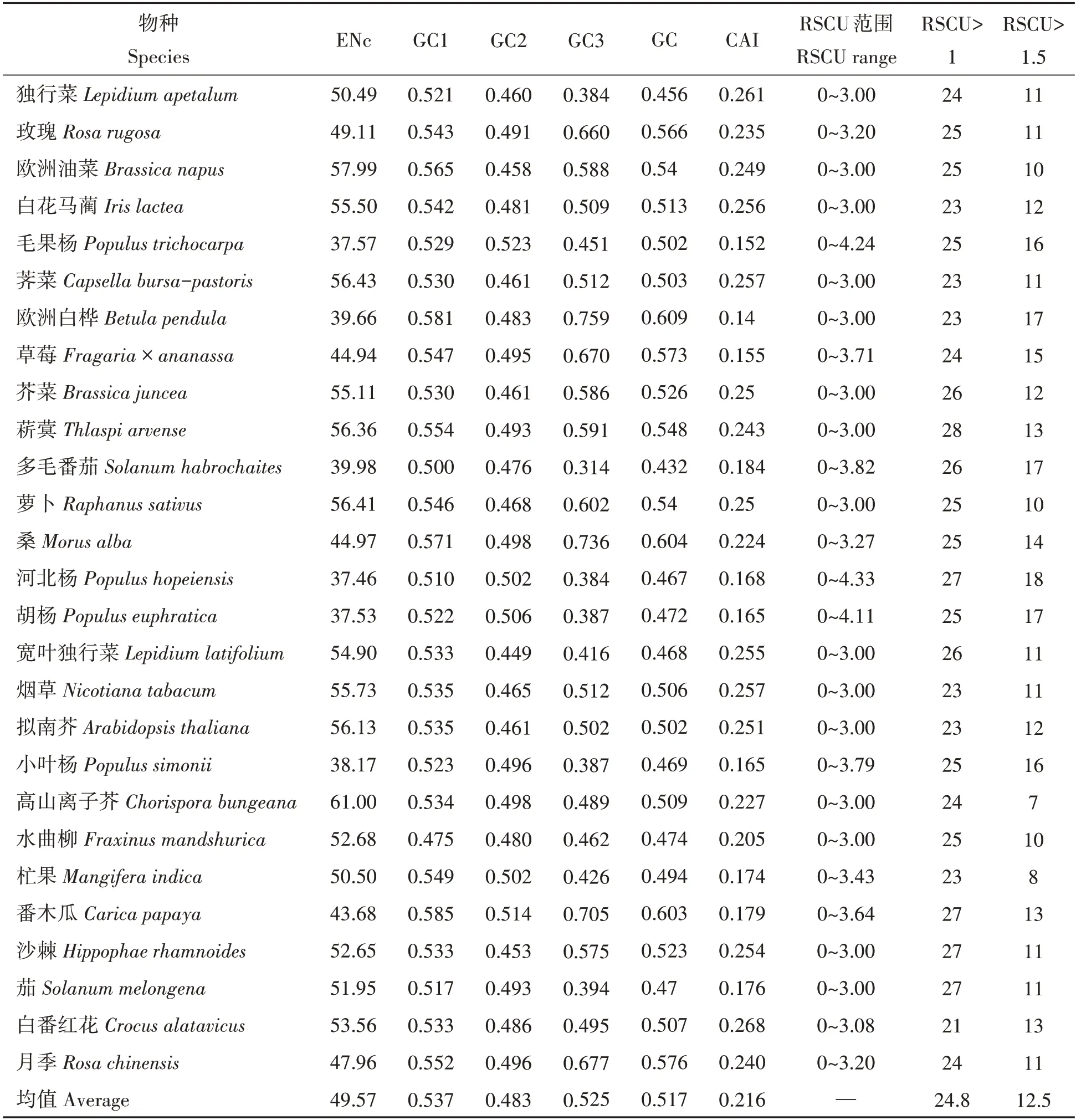
表3 不同物种DREB基因密码子使用偏性相关参数Tab.3 Preference related parameters of Analysis of DREB gene condons among different species
2.2.2 不同物种RSCU和GC含量分析 利用RSCU值分析,27个物种中RSCU值>1的密码子,数目最多的是菥蓂,为28个;最少的是白番红花,为21个。RSCU值>1.5的密码子,数目最多的是河北杨,为18个;最少的是高山离子芥,为7个(表3)。
27个物种GC3值为0.314~0.759,均值为0.525;GC含量值为0.430~0.608,均值为0.515。GC含量分析如表3,双子叶植物的GC、GC1、GC2和GC3含量的均值为:51.55%、53.68%、48.32%和52.66%,GC含量为52.11%。说明双子叶中DREB基因对GC的使用偏性更强,密码子第一位碱基偏好使用GC,第二位偏好使用AT,第三位偏好使用GC。单子叶植物的GC、GC1、GC2和GC3含量的均值为:50.78%、53.74%、48.37%和50.23%,说明单子叶植物中的DREB基因也对GC有较强使用偏性。不同物种DREB基因对GC和AT结尾的密码子偏性不同,其中,10个双子叶植物(独行菜、野生多毛番茄、河北杨、宽叶独行菜、茄、小叶杨、胡杨、水曲柳、杧果和拟南芥)GC含量小于0.5,密码子使用偏向于AT;其余单双子叶植物均对GC有一定使用偏性,欧洲白桦对GC使用偏性最强。独行菜LaDREB基因与双子叶植物相比有关参数在一定程度上相似,说明该基因在密码子使用特性上与双子叶植物密码子一致。
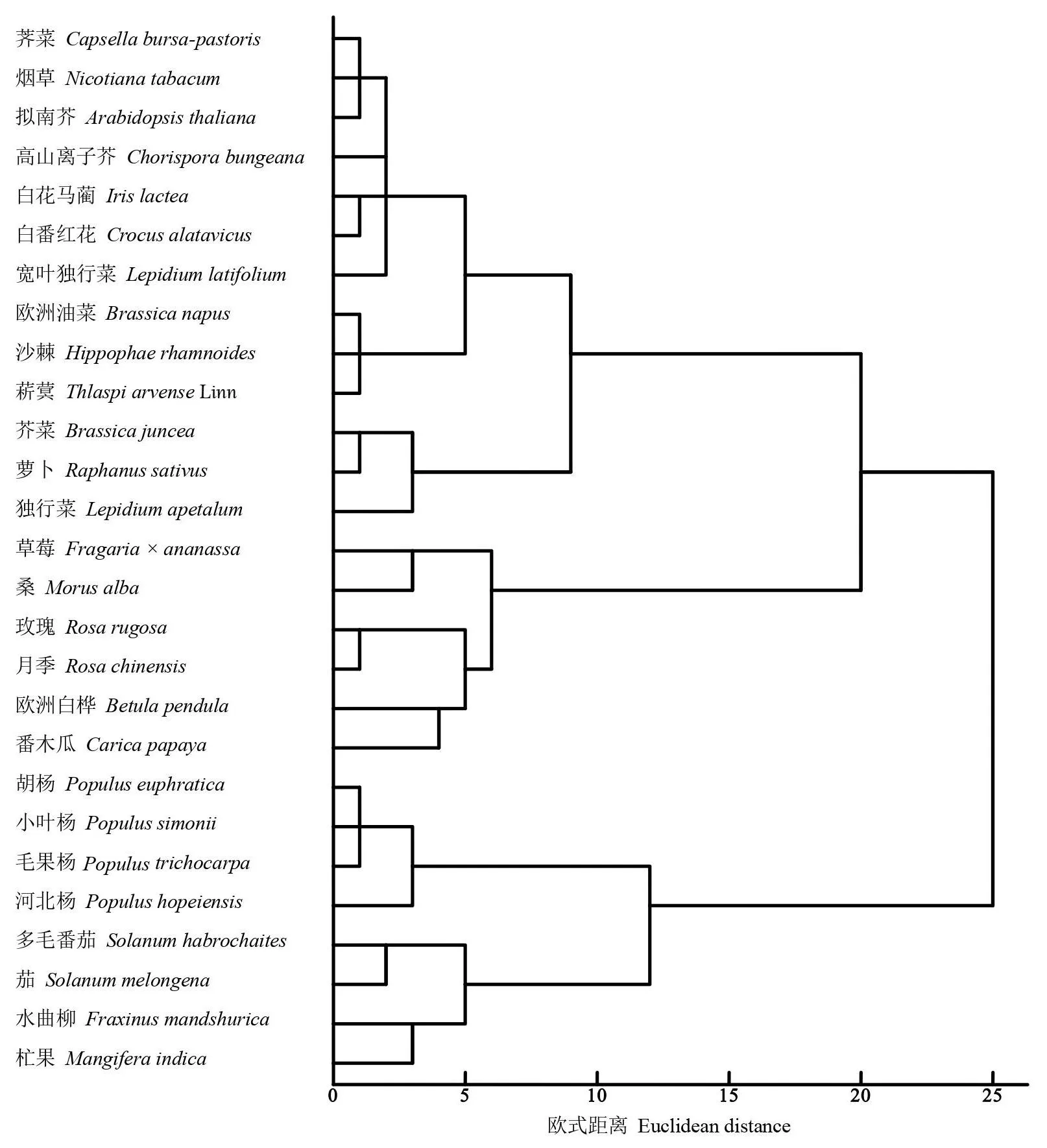
图1 DREB基因密码子使用偏性聚类分析Fig.1 Cluster analysis dendrogram of DREB genes based on codon bias
2.2.3 不同物种DREB基因CDS序列及RSCU值的系统聚类 对27个物种DREB基因的密码子偏性分析聚类。如图1,27个物种聚为两大类,多毛番茄、水曲柳、茄、杧果和4种杨柳科植物属于I类,其中4种杨柳科杨属植物归为一小类,2种茄科植物归为一小支。剩余的19种属于II类,在II类中,烟草、沙棘、2种单子叶植物和9种十字花科植物聚为一小类,其中单子叶植物白花马蔺和白番红花单独归为一小支。同属于独行菜属的宽叶独行菜和独行菜DREB基因没有在进化树中聚为一小类。整体上看,分类物种较近的DREB基因密码子使用频率也相近。但也存在分类相近而DREB基因密码子使用频率并不一定相近,如烟草和另外两种茄科植物的密码子偏性却相对较远。可见不同物种之间的DREB基因密码子,使用偏性和物种进化两者有时并完全相关。一些物种在进化过程中偏性使用某个独特的密码子,而DREB基因存在着丰富的密码子偏好性。

图2 DREB基因CDS序列系统聚类分析Fig.2 Cluster analysis of CDS sequences of DREB genes
对27个不同物种DREB基因CDS序列构建系统发育树(图2),发现27个物种基本按照十字花科植物和其他植物进行分类,单子叶植物和4种杨柳科杨属植物就仍旧各归为一小类,与密码子偏性聚类分析结果相似。而亲缘关系较近的十字花科独行菜属的独行菜与宽叶独行菜在CDS序列构建的进化树上归为了一小类。整体上DREB基因CDS序列的聚类分析结果与密码子偏性在不同植物中存在一致性,也与传统的物种分类基本一致,在一定程度上反映了DREB基因的一些特殊进化规律。但CDS序列的聚类分析结果也有一些与传统的物种分类规律不完全吻合,如茄科烟草的DREB基因的CDS序列与十字花科拟南芥同源性高于与茄科的茄及多毛番茄的同源性。
2.2.4 不同DREB基因的中性分析 采用SPSS19.0对GC、GC1、GC2、GC3和ENc,采用两两Pearson关联性分析。发现GC与GC1、CG3呈极显著正相关性(P<0.01),说明不同物种DREB基因第1和3位上的碱基成分有相似,见表4。不同物种DREB基因的中性分析见图3。发现GC3的范围为0.314~0.759,GC12的范围为0.478~0.549,波动较小,多数DREB基因分布在回归线左右。GC3和GC12的极显著相关系数是0.595,线性回归系数是0.080 2,对比Pearson结果相一致。表明不同物种DREB基因之间碱基组成差异不大,突变压力是影响密码子偏性的使用选择的关键[28−31]。独行菜LaDREB基因在回归线,距双子叶植物分布的位点较近,距单子叶植物分布位点较远,与两种聚类分析结果相一致。
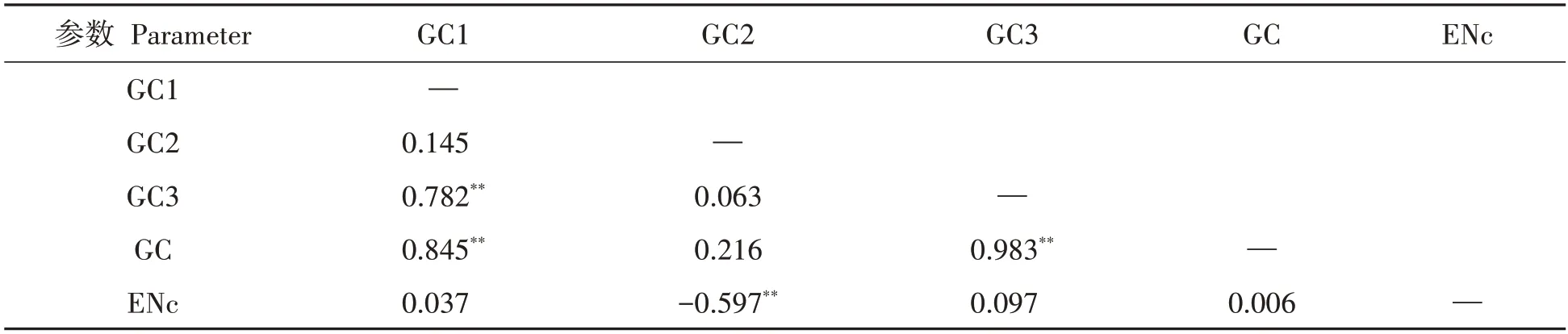
表4 DREB基因密码子成分相关性分析Tab.4 Correlative analysis of condon components of DREB related genes

图3 DREB相关基因密码子使用偏好中性绘图Fig.3 Neutrality plot on the codon usage bias of DREB related genes

图4 DREB相关基因的ENc−plot分布Fig.4 Effective number of codons used in DREB related genes
2.2.5 不同物种DREB基因ENc分析 对不同物种的DREB基因ENc值进行分析。结果显示(图4),ENc与GC3的期望曲线,能够说明突变压力与两者的对应关系,假如基因的对应的点在曲线上,表明该基因的密码子在使用选择上由突变压力所决定[32]。菥蓂、烟草和萝卜的基因在期望曲线上面,欧洲油菜、芥菜、拟南芥和茄等10个物种的基因分布在期望曲线下面,其余基因位点分布在期望曲线的上面。独行菜的基因靠近曲线与标准曲线分布趋势相吻合,且离期望曲线较近。表明独行菜LaDREB基因与相关基因的密码子使用偏性,碱基突变对其影响较大,和中性绘图结果一致。但一些基因离期望曲线位置远一些,表明DREB基因密码子的使用偏性的影响因素,除受碱基突变,其他因素如基因表达量、自然选择和基因长度等也对其有影响[33−34]。
2.2.6 不同物种DREB基因PR2分析 由图5可知,PR2分析27个物种DREB基因的A3/(A3+T3)或G3/(G3+C3)值均不在0.5左右。当比值不在0.5时,表明氨基酸在同义密码子的选择时,碱基突变和其他因素都会影响其偏性[35]。从横坐标显示,位点主要集中在<0.5的区域内,表明密码子在G与C之间偏向于C结尾。从纵坐标显示,位点大部分在小于0.5的范围值内,表明密码子使用选择时在A与T之间也具有偏好性,偏向于A结尾。可知自然选择等其他压力因素,在物种DREB基因之间的基因家族进化中发挥着一定的作用。
2.3 独行菜LaDREB基因受体系统的选择
独行菜的遗传转化体系仍未完善,独行菜基因还需其它完善的遗传转化受体来进行异源转化。将独行菜LaDREB基因与大肠杆菌、酵母菌、拟南芥、烟草、番茄、水稻和玉米基因组中不同密码子选择使用的频率相比较。如表5,独行菜LaDREB基因与大肠杆菌和酵母菌,分别有12和17个密码子偏性差异明显,说明大肠杆菌原核表达体系更适合独行菜LaDREB基因的异源表达。分析比较独行菜LaDREB基因与模式植物拟南芥、烟草和番茄密码子使用偏性,发现其与拟南芥、烟草和番茄的基因组选用频率差异较大个数分别为12、13和13;而水稻为12,玉米为20。表明拟南芥和水稻在独行菜LaDREB基因遗传转化功能验证中,都是适用的异源受体。而拟南芥与独行菜同属十字花科,与独行菜亲缘关系较近,能作为独行菜瞬时表达的良好受体。
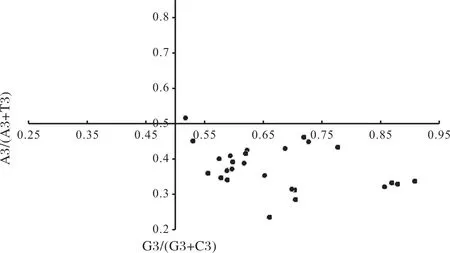
图5 奇偶偏好偏差性分析Fig.5 Parity rule 2(PR2)bias plot

表5 独行菜LaDREB基因与模式生物基因组密码子使用偏性比较Tab.5 Comparison of codon usage preference between LaDREB in Lepidium apetalum and other representative organisms

续表5
3 讨论
生物在长期进化中,形成了保证核苷酸与氨基酸之间的遗传信息能够准确传递特定的密码子选用偏好。基于基因突变和环境选择等因素[36−38],自然选择使基因表达时偏向使用最优密码子,然而长期的进化突变使得基因中出现一些非最优的密码子,最终导致物种基因在使用密码子时表现出偏性。
3.1 独行菜LaDREB基因密码子偏好性分析
本研究发现,独行菜LaDREB基因密码子的GC含量为45.60%,ENc值为50.49,CAI值为0.262。说明独行菜LaDREB基因的大部分密码子以AT结尾,碱基选用时偏向于GC,但选择偏性弱,且基因表达水平可能不高。根据独行菜LaDREB基因的RSCU值分析,GCU、CGU、AUU和CCA这4个密码子具有极强的使用偏性。
物种间DREB基因密码子使用偏性分析发现:单子叶植物以G/C结尾的密码子居多,与Wang等[39]研究结果一致。双子叶植物的也大部分以GC结尾密码子,但独行菜、毛果杨、胡杨等偏向以AT结尾的密码子。造成上述结果的原因,可能是基因的功能和类型有差异[40];也可能是不同植物的DREB基因在选择进化的过程中逐步选择偏向与以GC结尾的密码子,使其能够与基因的功能相适应。独行菜LaDREB基因密码子相关参数与双子叶植物相似,与十字花科植物最近。表明独行菜LaDREB基因密码子使用偏性与双子叶植物对密码子使用的选择偏性相似,且亲缘关系越近越相似。
3.2 独行菜LaDREB基因聚类分析比较
亲缘关系越近的物种,密码子使用偏好也越相近[41]。不同基因的密码子特殊选用偏性,造成聚类结果有时不能完全准确的反映真实系统分类和物种亲缘关系[8]。密码子使用偏好性聚类只在分类较小的单元中可作为分类依据,而在较大物种样本中,只能用于补充传统分类[42]。本研究中,RSCU和CDS聚类分析有一些差异。CDS聚类分析,将27个含DREB基因的物种分为两大类,同属独行菜属的独行菜LaDREB基因与宽叶独行菜DREB基因单独归为一小类。在聚类分析中,宽叶独行菜和独行菜并没有聚为一小类,但单子叶植物和4种杨柳科杨属植物就仍旧各归为一小类,与密码子偏性的RSCU聚类分析结果相似。因此,RSCU和CDS聚类分析结合有利于分类的精确,能更客观的体现物种间DREB基因的进化规律。
3.3 物种间DREB基因密码子使用偏性影响因素
密码子使用偏好性受自然选择和受突变压力的影响较多。然而密码子偏好性还与基因碱基组成、tRNA丰度、GC含量、基因表达水平、基因长度、甲基化、氨基酸保守性和mRNA二级结构等有关[43−44]。本研究对27个物种DREB基因的密码子进行PR2偏好性分析、中性绘图和ENc绘图,结果表明DREB基因的密码子偏好性受碱基突变影响较大,自然选择等其他因素对其密码子偏性影响较小。
3.4 独行菜LaDREB基因遗传表达转化受体系统的选择
独行菜LaDREB基因密码子使用频率分析发现,大肠杆菌原核表达系统更适用于独行菜LaDREB基因异源表达。但仍要根据使用的实际情况对某些密码子进行优化。通过与模式植物对比,得知独行菜与拟南芥、水稻的偏性差异一样最小,且拟南芥和独行菜亲缘关系较近同属于十字花科植物,故拟南芥是独行菜LaDREB基因最理想的遗传转化受体。但LaDREB基因是否具有较高的表达水平,还需要综合mRNA的结构、转化效率和转录后水平调控等因素。因此,独行菜LaDREB基因在异源植株高效遗传表达仍需深入研究。
本研究利用多种生物信息学分析方法,获得了独行菜LaDREB基因密码子使用偏性特征,明确了较合适的遗传表达受体。但还需根据情况在使用独行菜LaDREB基因时,对其密码子进行相应的改造和优化,以提高其在遗传转化研究中的表达效率。
