重金属Zn2+对凤尾鸡冠花2个品种种子萌发、幼苗生长及生理活性的影响
2021-01-07孙颖苗林琦孟儒臧帅彤高东菊张凤娥
孙颖 苗林琦 孟儒 臧帅彤 高东菊 张凤娥
摘要:以和服(Celosia cristata var. chilsii Kimono)、红塔山(Celosia cristata var. chilsii Hongtashan)2个凤尾鸡冠花品种种子及幼苗为研究对象,设置50、200、350、500 mg/L 4个浓度梯度,分析Zn2+胁迫对2个品种种子萌发及幼苗生长的影响,考察凤尾鸡冠花作为抗重金属污染植物材料的适应范围。结果表明:(1)随着Zn2+胁迫浓度的升高,和服种子的发芽率、发芽势和发芽指数呈逐渐降低趋势。红塔山种子在Zn2+溶液质量浓度为50 mg/L的胁迫下,各项萌发指标有所升高,表现为低促高抑。根据回归方程预测和服种子受Zn2+胁迫的临界值和极限值分别为886.35、1 485.87 mg/L,红塔山种子受Zn2+胁迫的临界值和极限值分别为898.62、1 574.30 mg/L。(2)重金属Zn2+浓度为 50 mg/L时,2个品种凤尾鸡冠花幼苗整株鲜质量、地上鲜质量均高于CK,且在Zn2+胁迫浓度为200、350 mg/L时,幼苗超氧化歧化酶(POD)、过氧化物酶(SOD)活性达到峰值,丙二醛(MDA)含量随胁迫浓度升高而逐渐升高。植物生长虽然受到影响,但仍能继续生长,表明这2个品种的凤尾鸡冠花对重金属Zn2+有一定抗性。
关键词:Zn2+胁迫;重金属;凤尾鸡冠花;种子萌发
中图分类号:S681.301 文献标志码: A
文章编号:1002-1302(2021)24-0133-06
收稿日期:2021-09-22
基金项目:上海农林职业技术学院中青年领军人才培养计划(编号:A2-0273-20-01)。
作者简介:孙 颖(1979—),女,吉林长春人,博士,副教授,研究方向为园林植物种质资源开发与利用。E-mail:46730244@qq.com。
通信作者:张凤娥,博士,讲师,研究方向为园林植物种质资源开发与利用。E-mail:16206@shafc.edu.cn。
重金属胁迫是全球普遍存在的一种非生物胁迫。据不完全统计,我国受重金属污染的土地面积近2 000万hm2,有毒重金属在土壤污染过程中具有隐蔽性、长期性、不可降解和不可逆转性,它们不仅导致土壤肥力下降、植物生长不良,而且通过食物链途径在植物、动物和人体内累积,其危害具有复杂性和严重性[1-3]。种子萌发是植物繁殖过程中重要的一环,种子萌发和幼苗生长是植物对外界环境适应的开始,也是植物对外界反应的敏感期和关键期[4-5]。已有研究表明,重金属对植物的伤害首先会表现在种子萌发和幼苗生长的变化上,因此,在胁迫条件下研究种子的萌发指标和生理变化情况,对了解植物的重金属耐受能力具有重要参考价值[6-8]。
锌既是植物生长必需的微量元素,又是土壤污染的主要重金属,不同浓度锌离子(Zn2+)对植物生长发育有重要影响。在自然环境中,锌既可作为检测土壤等环境污染的重要指标,也可作为筛选重金属超富集植物的指标[9]。凤尾鸡冠花(Celosia cristata var. chilsii)为苋科(Amaranthaceae)青葙属(Celosia)一年生草本植物,原产印度,其花穗形似火炬,花色明艳,花期较长,是夏秋少花季节难得的观赏花卉[10]。有研究发现,鸡冠花是一种重金属超富集植物,目前关于鸡冠花的重金属耐受能力研究已经开展,包括锰胁迫、镉胁迫等[11-12],但该种植物在锌胁迫下的相关研究还未见报道。本研究以凤尾鸡冠花和服、红塔山2个品种的种子为对象,研究二者对锌胁迫的响应机制,明确2个品种锌胁迫的适应范围,以期为重金属土壤污染修复的植物筛选和扩大凤尾鸡冠花的园林绿化应用提供理论依据。
1 材料与方法
1.1 试验材料
试验于2019年5月于上海农林职业技术学院实验室进行。供试的和服凤尾鸡冠花(C. cristata var. chilsii ‘Kimono’)、红塔山凤尾鸡冠花(C. cristata var. chilsii ‘Hongtashan’)種子均购自北京花仙子花卉公司。Zn2+溶液以ZnSO4·7H2O(分析纯)配制。试验设计Zn2+胁迫浓度梯度分别为0(CK)、50、200、350、500 mg/L。
1.2 试验方法
1.2.1 种子受Zn2+胁迫处理 采用纸上发芽法进行种子萌发胁迫试验。每个培养皿中加入等量的重金属溶液,随机且均匀摆放30粒种子,置于25 ℃恒温培养箱中培养。每个处理设3次重复,以等量的蒸馏水(CK)处理为对照。每个培养皿中除了溶液不同,保持其他因素一致。以胚根突破种皮长度超过2 mm作为发芽的标准[13],每天定时观察发芽情况,直到种子不再萌发或处于低水平萌发为止。
根据不同梯度浓度Zn2+溶液处理的2个品种种子发芽率、发芽势及发芽指数,计算种子耐金属胁迫的适宜浓度、半致死浓度及极限浓度。发芽率=正常发芽的种子数/供检种子总量×100%;相对发芽率=发芽率/对照发芽率×100%;发芽势=发芽种子数达到高峰时正常发芽种子总数/供检种子总数×100%;发芽指数(GI)=∑(Gt/Dt)(Gt为与Dt相对应的每天的发芽数;Dt为发芽试验天数)。
种子耐重金属胁迫半致死浓度为发芽率达到对照发芽率的50%的重金属溶液浓度;极限浓度为发芽率达到对照发芽率的25%的重金属溶液浓度。
1.2.2 复水处理 待种子发芽数不再发生变化时,将各处理未发芽的种子用清水清洗3次后放入加蒸馏水的新培养皿中,置于25 ℃恒温培养箱中培养。观察过程与“1.2.1”节一致。
萌发恢复率=复水后种子萌发的总数/重金属胁迫中未萌发的种子数×100%。
1.2.3 胚根长度的观测 在种子已经不再发芽或者处于低水平发芽时,在已经发芽的各处理中随机选择10粒种子,测量出每个胚根的长度。
1.2.4 幼苗受Zn2+胁迫处理 采用盆栽法进行鸡冠花植株培养。将鸡冠花种子均匀播种于含营养土的花盆(直径21 cm,高12 cm)中,土壤主要理化性质:全氮含量为1.37 mg/kg,P2O5含量为 0.79 mg/kg,K2O含量为0.84 mg/kg,有机质含量≥40%,有效态锌含量接近于0,微量元素含量≥0.2 mg/kg,pH值为6.5~8.5。每盆20粒种子,待种子出苗后,进行间苗,每盆保留10~15株苗,1周后加入等量的不同浓度Zn2+溶液[0(CK)、50、200、350、500 mg/L],放置室外培养。以上处理均为3次重复。
Zn2+胁迫处理5、10、15 d后,随机抽取每个处理组的10株幼苗,用自来水清洗并用吸水纸吸干表面残余水分后,用电子天平称取鲜质量,用游标卡尺测量主根长,计算根冠比。
1.2.5 幼苗生理指标的测定 Zn2+胁迫处理5、10、15 d时,分别选取0.2 g幼苗,置于 5 mL离心管中在球磨机内研磨成匀浆,加入4 mL的0.05 mol/L(pH值=7.8)磷酸缓冲液于离心管中,于 4 ℃条件下 10 000 r/min 冷冻离心 20 min,取上清液立即放入 4 ℃ 冰箱储存待测。参考张博宇等的方法[13-14]测定丙二醛(MDA)含量、过氧化物酶(POD)活性、超氧化物歧化酶(SOD)活性。
1.3 数据分析
采用Excel 2016软件进行试验数据统计及作图,采用SPSS 22.0检验样本间的差异显著性。
2 结果与分析
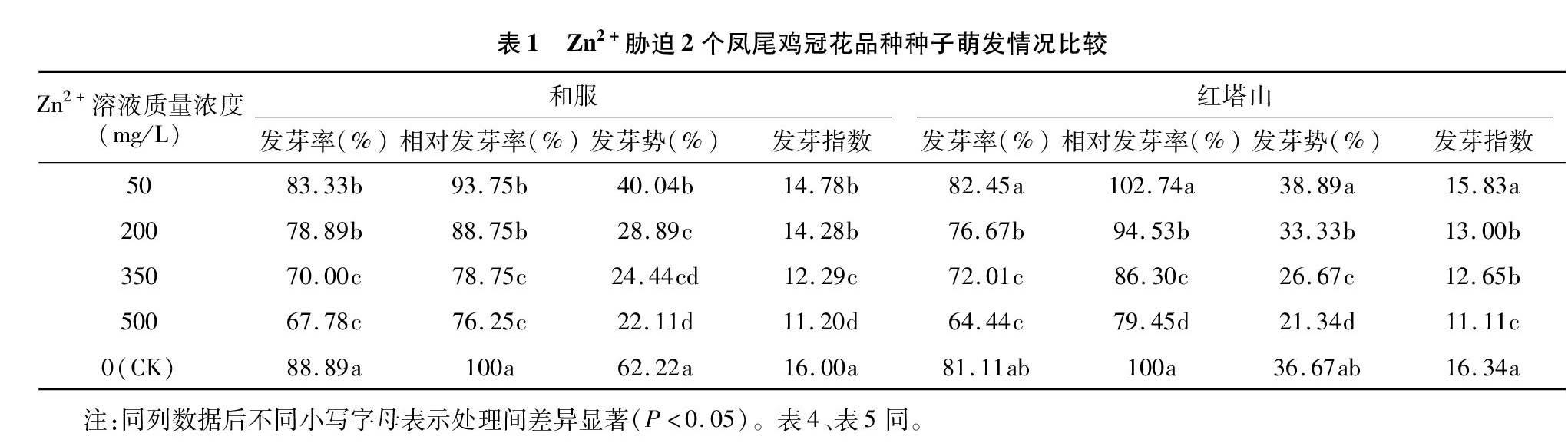
2.1 Zn2+胁迫对2个凤尾鸡冠花品种种子发芽率、发芽势、发芽指数的影响
从表1可以看出,随着Zn2+溶液质量浓度的提高,和服种子的发芽率、发芽势和发芽指数均呈下降趋势,且与CK相比,差异均显著(P<0.05)。50、200 mg/L 处理液之间差异不显著,350、500 mg/L处理液之间差异不显著,但350、500 mg/L Zn2+处理液胁迫下的发芽率显著低于50、200 mg/L处理液胁迫下发芽率(P<0.05)。其中,500 mg/L处理液胁迫下的发芽率为CK的76.25%。
红塔山凤尾鸡冠花种子的发芽情况与和服凤尾鸡冠花有所差异(表1)。主要表现为50 mg/L处理液的种子发芽率较CK有所提高,但差异不显著(P>0.05);处理液质量浓度为200 mg/L时,红塔山凤尾鸡冠花种子的发芽率、发芽势较CK有所降低,差异仍不显著(P>0.05);350、500 mg/L时种子发芽率才开始大幅降低,尤其处理液质量浓度为500 mg/L时,发芽率仅为CK的79.45%。发芽势与发芽率趋势一致,Zn2+处理液质量浓度为50 mg/L时,红塔山种子萌发整齐度比对照更高,说明低质量浓度的Zn2+处理对红塔山凤尾鸡冠花种子萌发有一定的促进作用,高质量浓度的Zn2+处理会抑制种子的萌发,表现为低促高抑。在相同质量浓度的Zn2+溶液处理条件下,红塔山种子的相对发芽率高于和服。
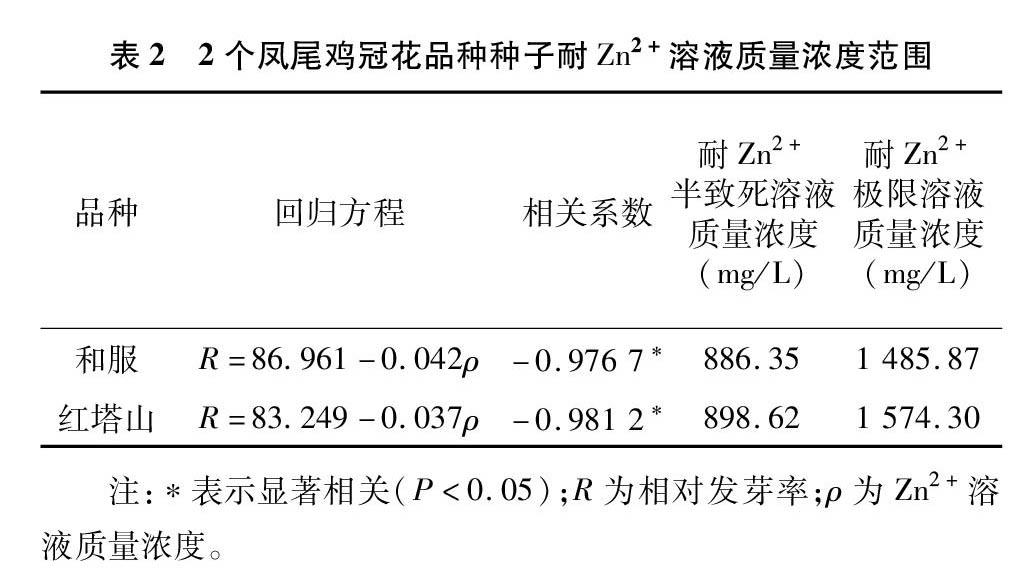
将Zn2+胁迫时种子发芽率与Zn2+质量浓度作相关分析,发现2个凤尾鸡冠花品种的发芽率与Zn2+溶液质量浓度均呈显著负相关(表2)。根据回归方程预测和服种子Zn2+胁迫的临界值和极限值分别为886.35、1 485.87 mg/L;红塔山种子Zn2+胁迫的临界值和极限值分别为898.62、1 574.30 mg/L。
2.2 Zn2+胁迫时2个凤尾鸡冠花品种种子复水效果
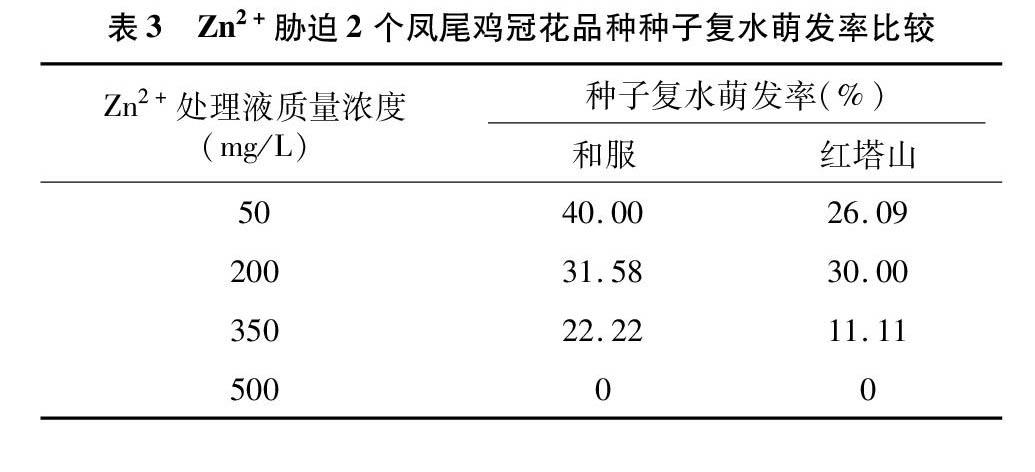
本试验中2个凤尾鸡冠花品种种子在Zn2+各胁迫浓度下的最终发芽率都超过60%。将胁迫后未萌发的种子进行复水萌发试验,和服种子在低质量浓度(50 mg/L)Zn2+溶液胁迫后,有40.00%的种子复水萌发(表3)。200、350 mg/L处理液胁迫后,复水萌发率虽没有50 mg/L处理液的高,但也有20%以上的发芽率。
红塔山种子总体的复水萌发率没有和服高。在200 mg/L处理液胁迫后的复水萌发率最高,为30.00%。其次為50、350 mg/L处理液胁迫后的复水萌发率。在500 mg/L处理液胁迫后,凤尾鸡冠花这2个品种种子的复水萌发率均为0。
2.3 Zn2+胁迫对2个凤尾鸡冠花品种种子胚根的影响
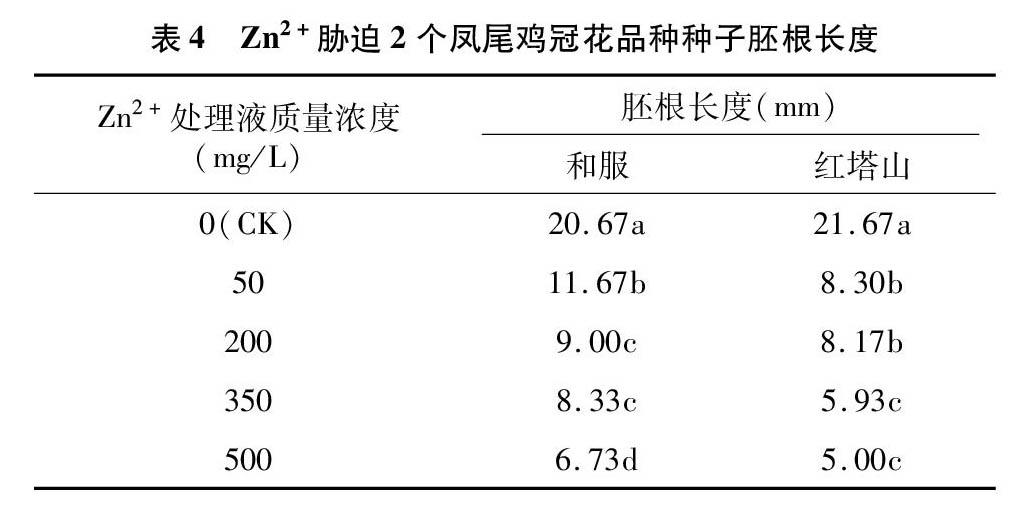
经过重金属Zn2+胁迫后的2个凤尾鸡冠花品种胚根生长速度明显减缓,胚根长度随胁迫浓度的提高而降低(表4)。当Zn2+处理液质量浓度为 50 mg/L 时,其对胚根的抑制作用很显著。当Zn2+处理液质量浓度达到500 mg/L时,和服和红塔山胚根长度仅分别为CK的32.56%、23.07%。同时观察发现,随着胁迫浓度的提高,2个凤尾鸡冠花品种种子的胚根颜色变深,根尖呈现灰色,子叶颜色由浅绿色变为黄绿色,上面夹杂黄斑。
2.4 Zn2+胁迫对2个凤尾鸡冠花品种幼苗生长的影响
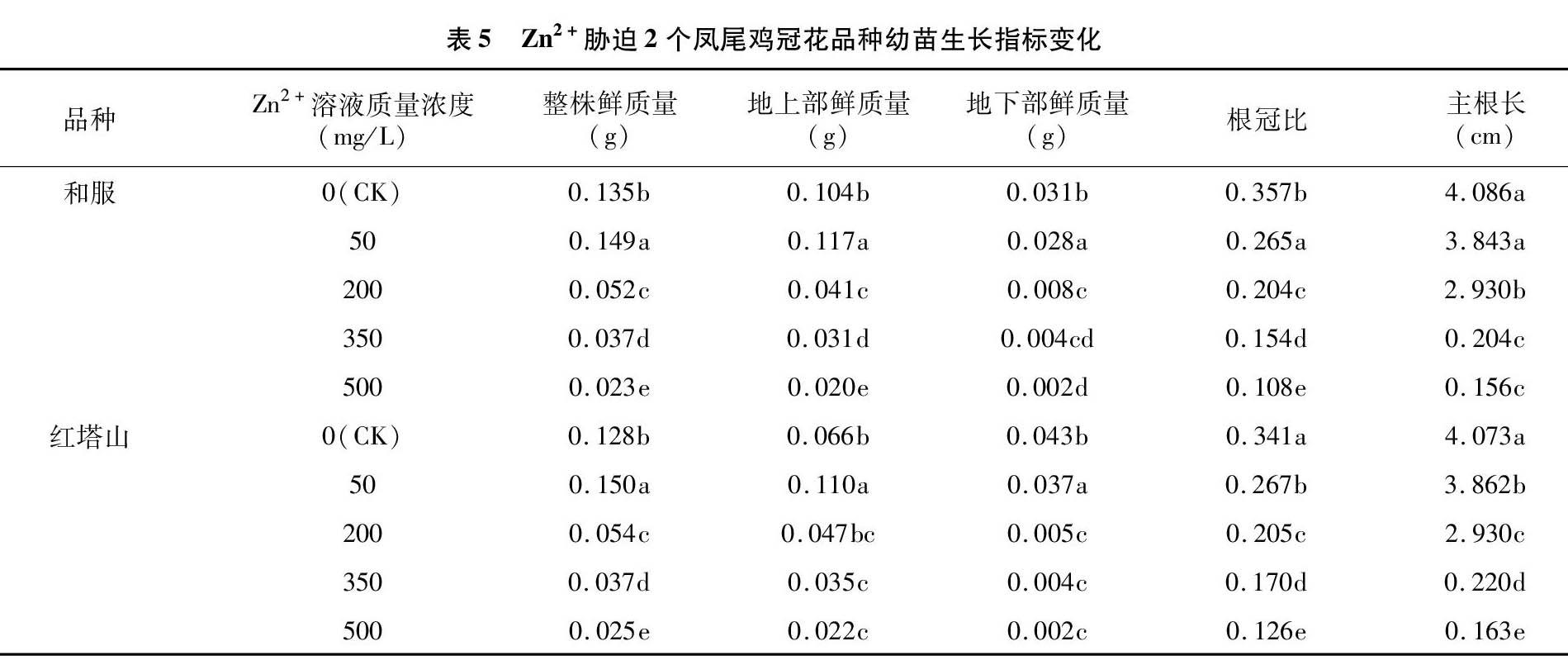
从表5可以看出,和服与红塔山2个凤尾鸡冠花品种经过不同浓度重金属Zn2+溶液处理后,幼苗地上部、整株鲜质量随处理液浓度升高呈现先增加后减少的趋势,地下部鲜质量则呈逐渐降低的趋势。在Zn2+处理液质量浓度为50 mg/L时,与CK比较,和服与红塔山的地上部分鲜质量显著增加,红塔山地下部分鲜质量增加也较为显著。当Zn2+处理液质量浓度为200 mg/L时,2个品种凤尾鸡冠花地上部分鲜质量及整株鲜质量明显降低,就整株鲜质量而言,和服较CK降低61.84%,红塔山较CK降低57.81%,在地上部鲜质量方面,两者分别较CK降低60.58%、28.79%,再随着处理液浓度的增加,地上鲜质量及整株鲜质量逐渐降低。
随Zn2+处理液质量浓度增加,和服与红塔山根长增长速度明显降低,主根长度也随之降低。当Zn2+处理液质量浓度为350 mg/L时,2个品种凤尾鸡冠花主根长明显降低,和服较CK降低95.01%,红塔山较CK降低94.55%。
2.5 Zn2+胁迫浓度对2个凤尾鸡冠花品种幼苗MDA含量及POD、SOD活性的影响
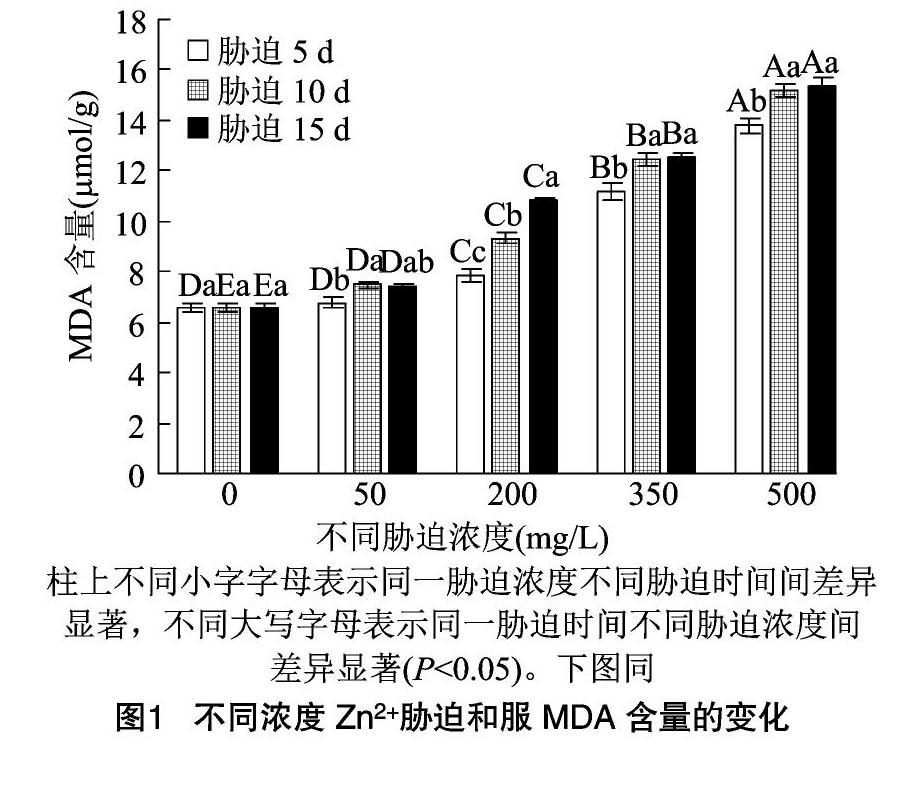
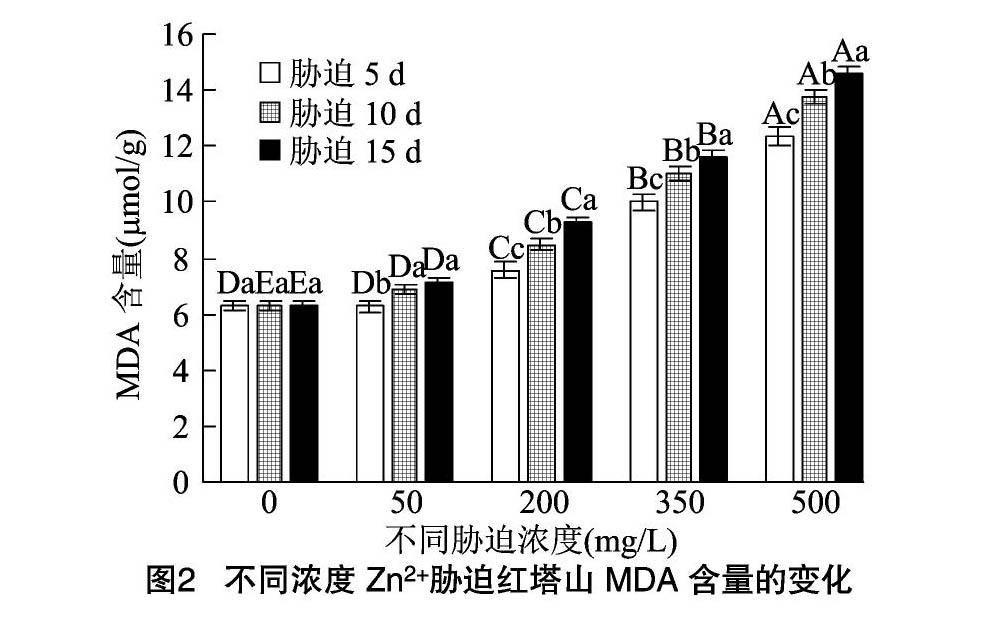
2.5.1 MDA含量 从图1、图2可以看出,2种凤尾鸡冠花的MDA含量随着胁迫浓度的提高呈现升高的趋势。当Zn2+处理液质量浓度为50 mg/L时,和服、红塔山受胁迫5、10、15 d的MDA含量与CK差异不显著(P>0.05);Zn2+处理液质量浓度为 200 mg/L 时,和服胁迫10 d后MDA含量有显著上升,达到CK的1.42倍,胁迫15 d时MDA含量达到CK的1.65倍;当Zn2+处理液质量浓度为500 mg/L时,和服胁迫5、10、15 d的MDA含量分别为CK的2.10、2.32、2.34倍,达到13.80~15.38 μmol/g。
Zn2+处理液质量浓度为200mg/L时,胁迫5、10、15 d红塔山的MDA含量分别为CK的1.20、1.35、1.47倍。当Zn2+处理液质量浓度达到 500 mg/L 时,红塔山受胁迫3个时间点的MDA含量分别为CK的1.96、2.19、2.32倍。表明200 mg/L的Zn2+胁迫对和服的影响较红塔山大,至500 mg/L时,红塔山的MDA含量也显著提高,尤其在胁迫 15 d 后,红塔山的MDA含量和和服持平。
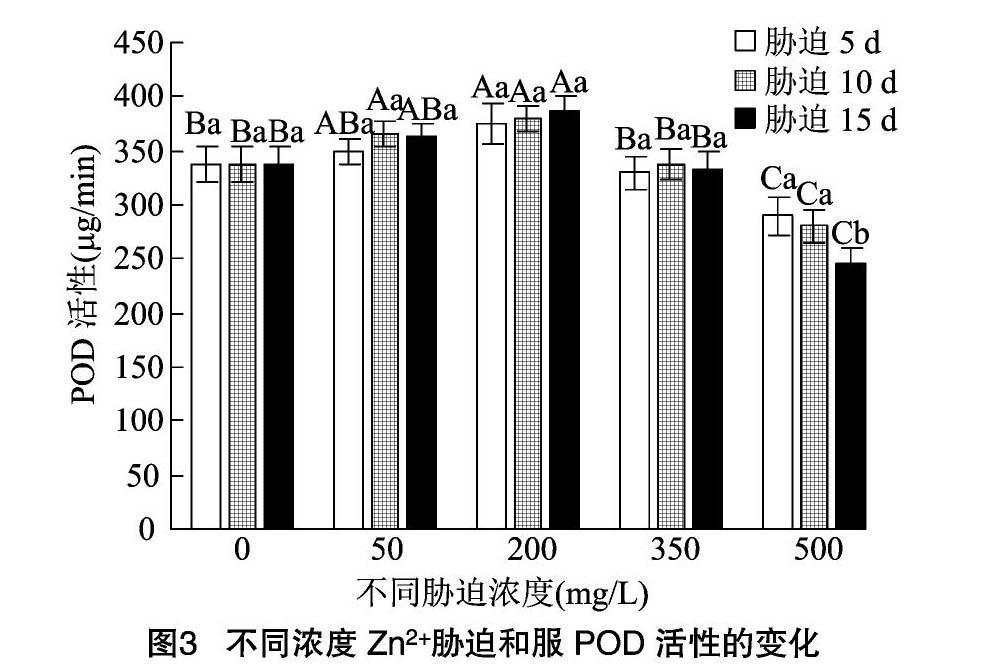
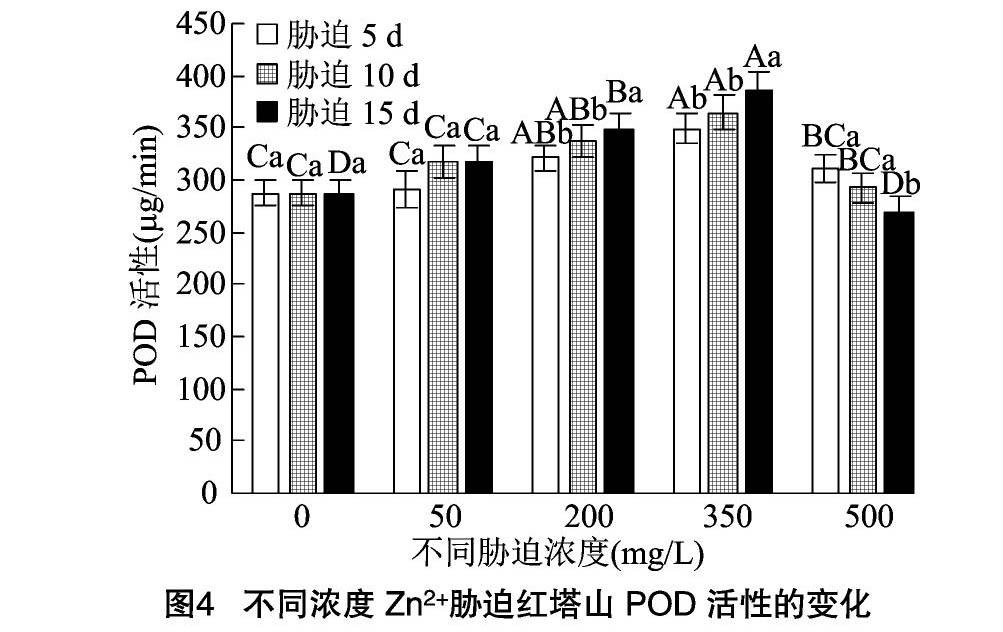
2.5.2 POD活性 从图3、图4可以看出,2种凤尾鸡冠花的POD活性则呈现先升高后降低的趋势。和服的POD活性在Zn2+处理液质量浓度为 200 mg/L 时达到峰值,胁迫5、10、15 d的POD活性分别为CK的1.11、1.13、1.15倍,3个时间节点之间差异不显著(P>0.05)。胁迫浓度为500 mg/L时,3个时间点的POD活性下降明显,分别为CK的0.86、0.83、0.73倍。其中胁迫15 d的POD活性降低幅度最大。
红塔山的POD活性在Zn2+处理液质量浓度为350 mg/L时达到峰值,此时3个时间节点的POD活性分别为CK的1.21、1.27、1.34倍。Zn2+处理液质量浓度为500 mg/L时,POD活性大幅降低,此时3个时间节点的POD活性分别为CK的1.08、1.02、0.94倍。其中胁迫15 d时的POD活性已经降到CK水平之下。
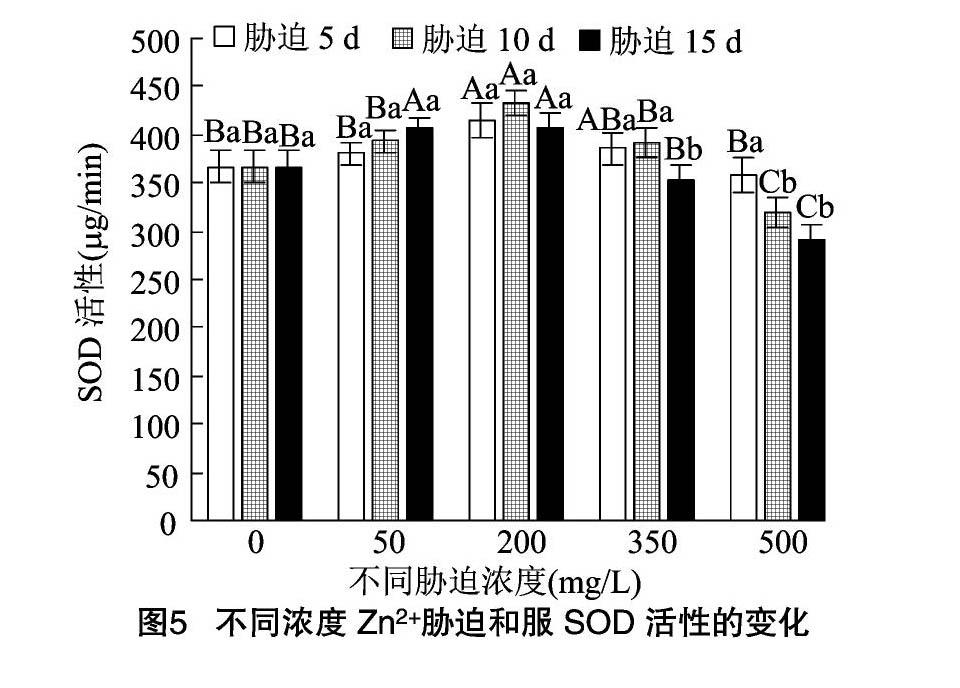
2.5.3 SOD活性 从图5、图6可以看出,2种凤尾鸡冠花的SOD活性变化趋势与POD类似,也呈现先升高后降低的趋势。和服的SOD活性在Zn2+处理液质量浓度为 200 mg/L 时达到峰值,在3个时间点分别为CK的1.13、1.18、1.11倍。胁迫浓度为350~500 mg/L时,3个时间点的SOD活性逐步下降。胁迫浓度达到500 mg/L时,SOD活性分别为CK的0.98、0.87、0.79倍。其中胁迫15 d的SOD活性降低幅度最大。
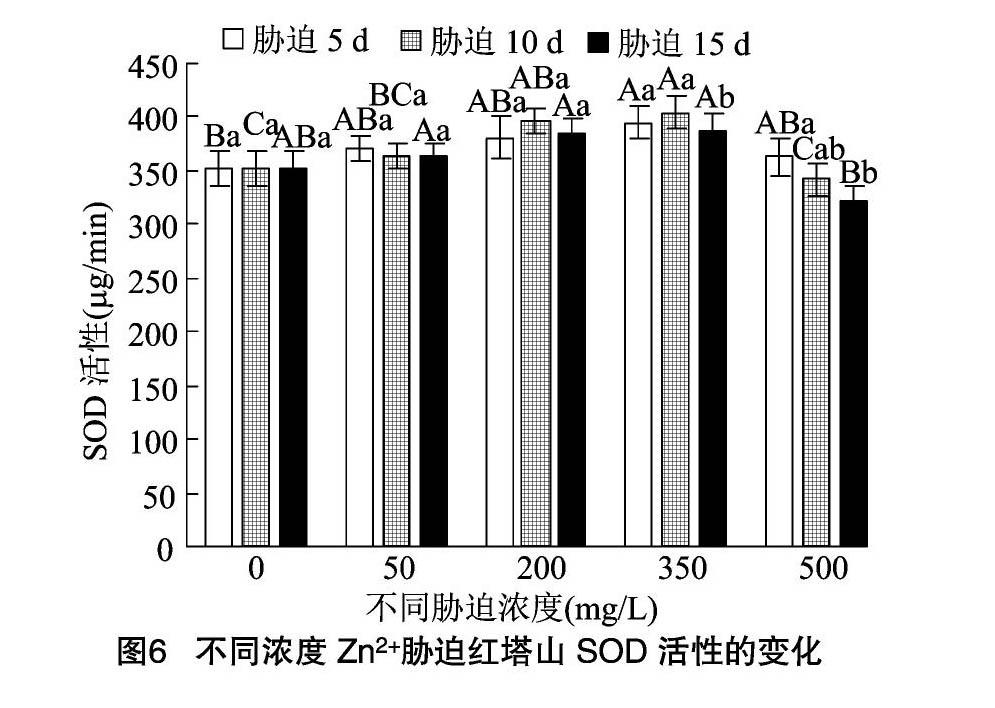
红塔山的SOD活性在Zn2+处理液质量浓度为350 mg/L 时达到峰值,200 mg/L的Zn2+胁迫与 350 mg/L 的Zn2+胁迫之间的SOD活性差异不显著(P>0.05)。500 mg/L Zn2+浓度条件下胁迫5 d时红塔山的SOD活性稍高于CK的SOD活性水平,胁迫10、15 d时SOD活性降至CK的0.97、0.91倍。
3 结论与讨论
对种子植物而言,种子萌发和幼苗生长不仅是植物生活周期的起点,也是植物感知外界环境的最初生命阶段。种子萌发时期的生长状况将直接影响植物以后的生长[15]。在重金属污染环境中,种子的生长情况会受到很大程度的影响,如种子出苗率低、出苗不整齐、活力低,将导致植物减产,最终达不到植物对于重金属污染的显著治理效果[16]。锌(Zn)作为一种植物必需的微量元素,是RNA聚合酶的必需元素,可以稳定核糖体结构[17]。在一定浓度范围内,Zn能直接或间接参与植物体内的光合作用、呼吸作用以及碳水化合物的合成、运转等过程,促进植物的生长和提高其抗逆能力[18-19]。然而,当环境中的 Zn 含量超过某一临界阈值时,会降低植物光合作用速率,破坏植物体的离子平衡,造成缺铁性失绿等,从而使植物生理紊乱、生长状况不良,甚至导致植物死亡[20-21]。俞明惠等的研究表明,随着 Zn2+浓度的增加,矮秆波斯菊(Cosmos bipinnata)种子发芽势、发芽率、发芽指数表现为低促高抑,其中10 mg/L 的促进作用最为明显[22];陈丽丽等在对稗草和地肤种子的研究中也发现,Zn2+在设置的浓度范围内对2杂草种子萌发呈现明显的低促高抑现象,但各指标表现不完全一致[23]。钟静等发现,40 mg/L 硫酸锌处理对薏苡种子萌发并无明显抑制效果,而40 mg/L硫酸锌处理则显著抑制薏苡幼苗主根和苗的生长,说明薏苡幼苗对锌胁迫的敏感性要明显高于种子[24-25]。
在本研究中,和服凤尾鸡冠花种子的发芽率、发芽势、发芽指数,均随着Zn2+处理液质量浓度的升高而降低。本研究中各指标与胁迫浓度呈负相关关系,但红塔山凤尾鸡冠花种子在Zn2+处理液质量浓度为50 mg/L时,各项萌发指标有所升高,说明低质量浓度的Zn2+处理刺激了红塔山凤尾鸡冠花种子自身的抗逆性反应,在形态学上呈现发芽率、发芽势暂时升高的现象。而高质量浓度的重金属Zn2+离子,破坏植物的自我防御机制,为了在胁迫下维持细胞内离子平衡,消耗植物的有效能量,必将造成植物生长发育受阻[26-27]。本研究结果中,2个凤尾鸡冠花品种在质量浓度为350 mg/L的处理液中各项萌发指标开始出現显著降低,但在500 mg/L处理液中仍有64%以上的发芽率,说明这2个凤尾鸡冠花品种对Zn2+胁迫具有较强的耐受性,其中红塔山的耐受性略高于和服的耐受性。
通常重金属对植物根的抑制作用大于芽,种子萌发后,根最先突破种皮吸水,从而使根尖的重金属累积量以及受胁迫时间较重[28-29]。从2个凤尾鸡冠花品种胚根的生长情况看,Zn2+处理对二者胚根生长均表现出较强的抑制作用,这种抑制作用从低质量浓度Zn2+处理时即开始出现。Zn2+质量浓度为50 mg/L时,胚根颜色变深说明低质量浓度已经造成了根尖细胞的异常分裂,胚根生长滞缓。而较高质量浓度的Zn2+胁迫甚至导致凤尾鸡冠花种子根尖受害显著,发芽后几乎不再生长,成坏死状态。2个凤尾鸡冠花品种在Zn2+质量浓度为500 mg/L时仍有较高的萌发率,但在萌发后会出现胚根受Zn2+胁迫影响严重的现象,说明裸露的胚根对Zn2+胁迫反应更为敏感。
本研究中,2个凤尾鸡冠花幼苗在重金属Zn2+的胁迫下,其整株鲜质量均呈现低浓度(50 mg/L)时升高,高浓度时则下降,其幼苗POD、SOD活性在胁迫浓度为200~350 mg/L时达到峰值,这些与种子低促高抑表现趋势相同。MDA含量反映植物受外界侵害程度,2个品种凤尾鸡冠花幼苗在胁迫浓度从350 mg/L到500 mg/L时仍旧能够继续生长,说明红塔山与和服均对重金属Zn2+存在抗性。 当植物处于逆境中,会产生大量活性氧,POD与SOD均可清除活性氧,维持植物生长[30]。本研究中,和服与红塔山幼苗受到胁迫时,POD、SOD的活性呈现先升高后下降的趋势,说明这2种凤尾鸡冠花对重金属Zn2+有一定抗性,但当胁迫过强时,2种酶的活性开始下降,植物的抵抗能力也有所下降。
如何通过螯合剂诱导植物修复技术、接种菌根强化植物吸收技术及施加土壤改良剂来提高凤尾鸡冠花的吸附能力,同时对凤尾鸡冠花在Zn2+污染土壤上的生长情况、地上部分产量情况进行定量分析将是今后研究的重点。
参考文献:
[1]陶 玲,任 珺,祝广华,等. 重金属对植物种子萌发的影响研究进展[J]. 农业环境科学学报,2007,26(增刊1):52-57.
[2]Nedelkoska T V,Doran P M.Characteristics of heavy metal uptake by plant species with potential for phytoremediation and phytomining[J]. Minerals Engineering,2000,13(5):549-561.
[3]赵香香,韩超南,吴 昊,等. 藻体分解对沉积物-水中硫迁移转化的影响[J]. 森林工程,2020,36(6):36-41.
[4]李晓科,武玉珍,张义贤.酸雨对Cd胁迫下大麦种子萌发及幼苗生长的影响[J]. 种子,2019,38(7):71-75.
[5]陈帅全,张 驰,王 印,等. 碱热处理生物质碳源材料比选研究[J]. 森林工程,2020,36(5):99-105.
[6]何俊瑜,任艳芳.镉胁迫对水稻种子萌发、幼苗生长和淀粉酶活性的影响[J]. 华北农学报,2008,23(增刊1):131-134.
[7]宋玉芳,许华夏,任丽萍,等. 土壤重金属对白菜种子发芽与根伸长抑制的生态毒性效应[J]. 环境科学,2002,23(1):103-107.
[8]陈泽华,焦 思,余爱华,等. 土壤重金属污染评价方法探析:以南京市为例[J]. 森林工程,2020,36(3):28-36.
[9]张小敏,张秀英,钟太洋,等. 中国农田土壤重金属富集状况及其空间分布研究[J]. 环境科学,2014,35(2):692-703.
[10]王巨媛,翟 胜,任秋萍,等. 不同盐胁迫下鸡冠花种子萌发期的耐盐性研究[J]. 广东农业科学,2010,37(6):92-93.
[11]陈利云,呼丽萍,汪之波.内生菌醇提取物对鸡冠花生长与抗逆性的影响[J]. 江西农业学报,2020,32(3):61-64.
[12]雷 玲,刘 杰,蒋萍萍,等. 超富集植物青葙叶片对锰胁迫的生理和激素代谢响应[J]. 应用与环境生物学报,2020,12(18):1-9.
[13]张博宇,滕维超.铅胁迫对黄花风铃木幼苗生长和生理指标的影响[J]. 东北林业大学学报,2020,48(7):7-10,16.
[14]陈茂铨,应俊辉,王东明,等. 铅胁迫对萝卜种子萌发、幼苗生长及生理特性的影响[J]. 江苏农业科学,2010,38(2):172-174.
[15]刘拥海,俞 乐,林馥丽.不同重金属胁迫对绿豆种子萌发和幼苗初期生长影响的差异[J]. 种子,2007,26(11):41-44.
[16]帅海威,孟永杰,罗晓峰,等. 生长素调控种子的休眠与萌发[J]. 遗传,2016,38(4):314-322.
[17]梁 剑. 锌胁迫对油橄榄生理因子的影响[J]. 环境研究与检测,2016(4):13-16.
[18]Gupta N,Ram H,Kumar B.Mechanism of zinc absorption in plants:uptake,transport,translocation and accumulation[J]. Reviews in Environmental Science and Biotechnology,2016,15(1):89-109.
[19]Valko M,Jomova K,Rhodes C J,et al. Redox-and non-redox-metal-inducedformationoffreeradicals and their role in human
disease[J]. Archives of Toxicology,2016,90(1):1-37.
[20]Michael P I,Krishnaswamy M.The effect of zinc stress combined with high irradiance stress on membrane damage and antioxidative response in bean seedlings[J]. Environmental and Experimental Botany,2011,74:171-177.
[21]Borowiak K,Gsecka M,Mleczek M,et al. Photosynthetic activity in relation to chlorophylls,carbohydrates,phenolics and growth of a hybrid Salix purpurea×triandra×viminalis 2 at various Zn concentrations[J]. Acta Physiologiae Plantarum,2015,37(8):1-12.
[22]俞明惠,程 玉,范志强,等. 锌离子对矮秆波斯菊种子萌发及幼苗生长的影响[J]. 现代农业科技,2020(1):116-117,119.
[23]陈丽丽,付媛媛,王艳萍,等. 镉锌胁迫对小麦和2种杂草种子萌发及幼苗生长的影响[J]. 西南林业大学学报,2019,39(1):50-57.
[24]钟 静,王亮节,曾国平,等. 锌胁迫对薏苡种子萌发和幼苗生长的影响[J]. 北方园艺,2020(2):113-118.
[25]张 珂,厉萌萌,刘德权,等. 镉胁迫对小麦、玉米种子萌发及幼苗生长的影响[J]. 种子,2019,38(5):90-94.
[26]Zhou L Y,Zhao Y,Wang S F,et al. Lead in the soil-mulberry (Morus alba L.)-silkworm (Bombyx mori) food chain:Translocation and detoxification[J]. Chemosphere,2015,128:171-177.
[27]王 波,黃 攀,吕德雅,等. 铅、镉对南荻种子萌发和幼苗生长的影响[J]. 生态环境学报,2018,27(9):1768-1773.
[28]陈俊任,柳 丹,吴家森,等. 重金属胁迫对毛竹种子萌发及其富集效应的影响[J]. 生态学报,2014,34(22):6501-6509.
[29]张 航,战金雨,杨 柳,等. NaCl胁迫对榆树种子萌发的影响[J]. 江苏农业科学,2021,49(9):127-131.
[30]张永侠,原海燕,顾春笋,等. 红籽鸢尾(Iris foetidissima L.)的抗旱性[J]. 江苏农业科学,2014,42(8):174-177.
