大豆质核互作雄性不育系花器官发育生理生化特性的变化
2021-01-07崔天宇曹霞孙佳尧李志刚刘鹏王鹏年
崔天宇 曹霞 孙佳尧 李志刚 刘鹏 王鹏年



摘要:以大豆质核互作雄性不育系JLCMS9A及其同型保持系JLCMS9B为试验材料,测定花芽期、花蕾期、成花期中超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)的活性以及丙二醛(MDA)、淀粉、可溶性蛋白、可溶性糖、游离脯氨酸的含量,分析3个时期生长素(IAA)、赤霉素(GA3)、异戊烯基腺嘌呤核苷(iPA)、脱落酸(ABA)的含量及变化。结果表明,在花芽期不育系中的SOD、CAT活性以及MDA、游离脯氨酸的含量均高于保持系,而在花蕾期和成花期相反,其值均低于保持系;不育系的POD活性在花芽期显著低于保持系,在花蕾期和成花期相反,其值均高于保持系;不育系9A的淀粉、可溶性糖含量整体呈上升趋势,在花蕾期和成花期均低于保持系9B。花器官整个发育过程中,不育系9A的IAA含量呈先升后降的变化趋势,iPA含量为逐渐升高趋势,不育系各激素含量均低于保持系。不育系的IAA/ABA、IAA/GA3、IAA/iPA、ABA/GA3的比值与保持系存在差异。由此推断,花器官发育生理特性的异常与大豆质核互作雄性不育有关,花蕾期可能是大豆质核互作雄性不育系花器官生理生化指标发生异常的关键时期。
关键词:大豆;质核互作雄性不育;花器官;生理生化特性;抗氧化酶活性;内源激素含量
中图分类号:S565.101 文献标志码: A
文章编号:1002-1302(2021)24-0075-07
收稿日期:2021-04-07
基金项目:博士科研启动基金(编号:BS514);内蒙古自治区科技储备项目(编号:2018MDCB02);内蒙古自治区科技计划(编号:2018KJJH1702);内蒙古民族大学科研立项项目(编号:NMDSS2159)。
作者简介:崔天宇(1996—),女,内蒙古赤峰人,硕士,主要研究方向为作物遗传育种。E-mail:2105842265@qq.com。
通信作者:李志刚,博士,教授,主要研究作物遗传育种。E-mail:13948651158@126.com。
大豆原产于我国,迄今已有5 000多年的栽培史,因此我国有丰富的大豆资源。大豆作为主要粮食作物之一,是人类所需蛋白质和食用植物油的主要来源。随着我国经济的发展,各方面对大豆的需求也日益增加。但由于大豆的产量和经济效益较低,导致其种植面积低于玉米、水稻、小麦等主要粮食作物,我国大豆总产量明显供不应求。因此培育高产、优质、抗性强的大豆是我国大豆育种的首要目标,而实现这一目标最有效的措施就是利用杂种优势。大豆质核互作雄性不育系的发现为大豆杂种优势的利用提供了基础[1]。
近年来,对于棉花[2]、水稻[3]、油菜[4]、烟草[5]、白菜[6-8]、玉米[9]等生理生化特性的研究较多,而关于大豆不育系及其生理生化特性的研究却鲜有报道。研究者们发现植物雄性不育与其器官的物质代谢、能量代谢、内源激素以及其抗氧化酶活性有关。雄性不育系体内的物质含量减少,代谢速度降低。研究指出抗氧化酶[超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)]具有清除植物代谢过程中产生的氧自由基的作用,可以保持质膜的稳定性[10],从而维持植物体内活性氧的平衡。内源激素与植物的不育性有关,是调节其生长发育的重要因子[11-12]。
我国大豆雄性不育的主要类型有细胞核雄性不育[13-15]、质核互作雄性不育[16-18]和光温敏雄性不育3种[19-21]。本试验以大豆质核互作雄性不育系及其同型保持系作为试验材料,通过测定花器官发育不同時期中的SOD、POD、CAT活性,丙二醛(MDA)、淀粉、可溶性蛋白、可溶性糖、游离脯氨酸的含量以及生长素(IAA)、赤霉素(GA3)、异戊烯基腺嘌呤核苷(iPA)、脱落酸(ABA)等内源激素含量及其动态含量的变化,探究其与不育性的相关性,以期为选育大豆优良不育系及不育机制研究提供理论依据。
1 材料与方法
1.1 试验材料
以吉林省农业科学院大豆研究所提供的大豆不育系JLCMS9A与同型保持系JLCMS9B为试验材料,下文分别用9A、9B表示。于2019年5月播种于内蒙古民族大学北区试验地,试验设为3个小区,每个小区的面积为20 m2,采用等行距相间种植,株行距为15 cm×60 cm,每个小区种植6行。试验材料均在同一时期进行播种,且在生长期间采用相同的管理措施进行田间管理。
1.2 样品采集
于盛花期开始取样,对主茎第4节位以上的花芽、花蕾、成花分别进行取样,重复3次,将样品放入液氮进行速冻,然后放入-80 ℃冰箱保存。
1.3 测定项目与方法
可溶性蛋白含量采用考马斯亮蓝G-250染色法测定,可溶性糖及淀粉含量采用硫酸蒽酮比色法测定,游离脯氨酸含量采用酸性茚三酮法测定,CAT活性采用紫外吸收法测定,SOD活性采用氮蓝四唑(NBT)光还原法测定,POD活性采用愈创木酚法测定,MDA含量采用硫代巴比妥酸(TBA)法测定。
采用间接酶联免疫(ELISA)法分别测定不育系及同型保持系花器官不同发育时期生长素(IAA)、赤霉素(GA3)、异戊烯基腺嘌呤核苷(iPA)和脱落酸(ABA)的含量,3次重复。
1.4 数据分析
利用软件DPS 16.05对测定指标数据间的差异显著性进行分析,使用Excel 2016进行柱状图绘制。
2 结果与分析
2.1 花器官发育不同时期抗氧化酶活性的变化
如图1所示,在花器官发育过程中,不育系9A的SOD活性呈下降趋势,且在成花期达到显著性差异;保持系9B的SOD活性则是先升高后降低,且花蕾期达到最高值(254.58 U/g),达到显著性差异。在花蕾期、成花期,保持系9B的SOD活性分别是不育系9A的1.18、1.58倍,均达到显著性差异。在成花期,不育系9A的SOD活性骤降,推测成花期可能是发生败育的时期。
不育系9A及保持系9B的POD活性在花器官发育时期均呈先升高后降低的变化趋势,且二者同时在花蕾期达到最高值,但差异不显著。在花芽期9B的POD活性比9A高9.5%,差异显著。成花期9B的POD活性大幅降低,是9A的94.5%,差异显著。不育系与保持系的POD活性在成花期较花蕾期均显著降低,但在整个发育时期不育系和保持系表现出相同的倒U形变化规律。因此,推测POD活性与大豆雄性不育相关性不大。
不育系9A和保持系9B的CAT活性呈不同的变化趋势,9A呈逐渐降低的变化趋势,而9B呈倒U形的变化趋势。随着花器官的发育,9A、9B成花期CAT的活性均低于花芽期,且达到了差异显著水平;其中,在花芽期9A比9B高1.75倍,而在成花期9B比9A高2.03倍,说明随着花器官的发育,CAT活性在逐渐降低,且不育系9A比保持系9B显著性降低。从花芽期至成花期,不育系9A的MDA含量呈先降低后升高的变化趋势,而保持系9B呈逐渐下降的变化趋势。在花芽期、花蕾期、成花期,不育系9A与保持系9B均达到了显著性差异,并且不育系9A的MDA含量分别是保持系9B的1.22、1.23、1.41倍。在成花期不育系9A与保持系9B的MDA 含量差距最大, 说明成花期MDA含量对育性影响较大。
2.2 花器官不同发育时期的营养物质含量比较
由图2可以看出,不育系9A、保持系9B的淀粉含量均呈递增的变化趋势,均在成花期达到最高值,分别为11.41、21.76 mg/g。在花器官发育过程中,9B的淀粉含量始终高于9A,且在花蕾期和成花期差异显著。在花蕾期、成花期,9B的淀粉含量比9A分别高出52.4%、90.7%,说明随着花器官的发育,淀粉含量对大豆育性起关键作用。由于后期淀粉亏缺,花器官发育缺少能量和营养物质,使花器官发育异常,导致雄性不育。
在整个花器官发育过程中,不育系9A和保持系9B的可溶性蛋白含量在花器官发育时期均呈逐渐下降趋势,且不育系9A与保持系9B的可溶性蛋白含量均在花芽期达最大值,分别为157.25、169.86 mg/g,且达到了显著性差异。随着花器官的发育,可溶性蛋白含量逐渐降低,在成花期无显著性差异;但在花芽期、花蕾期,保持系9B的可溶性蛋白含量分别是不育系9A的1.08、1.09倍,且达到了显著性差异,说明花芽期、花蕾期可能是发生不育的时期。
从花芽期到成花期,不育系9A和保持系9B的可溶性糖含量变化一致,均呈递增变化趋势。在花器官发育过程中,9B的可溶性糖含量均高于9A,且在花芽期和花蕾期差异不显著;而在成花期达到最大值,分别高达11.33、16.15 mg/g,且9B是9A的1.43倍,差异显著。说明花器官发育后期,可溶性糖对育性起着关键性作用,与发生不育的时期有关。
在花器官整个发育过程中,不育系9A与保持系9B的游离脯氨酸含量均呈倒U形变化趋势。随着花器官的发育,花蕾期的游离脯氨酸含量高于花芽期和成花期,且达到了显著性差异;其中,在花芽期9A显著高于9B,是9B的1.37倍;在花蕾期、成花期,9B则显著高于9A,分别是9A的1.30、1.40倍,说明不育系中游离脯氨酸含量的显著降低可能导致雄性不育。
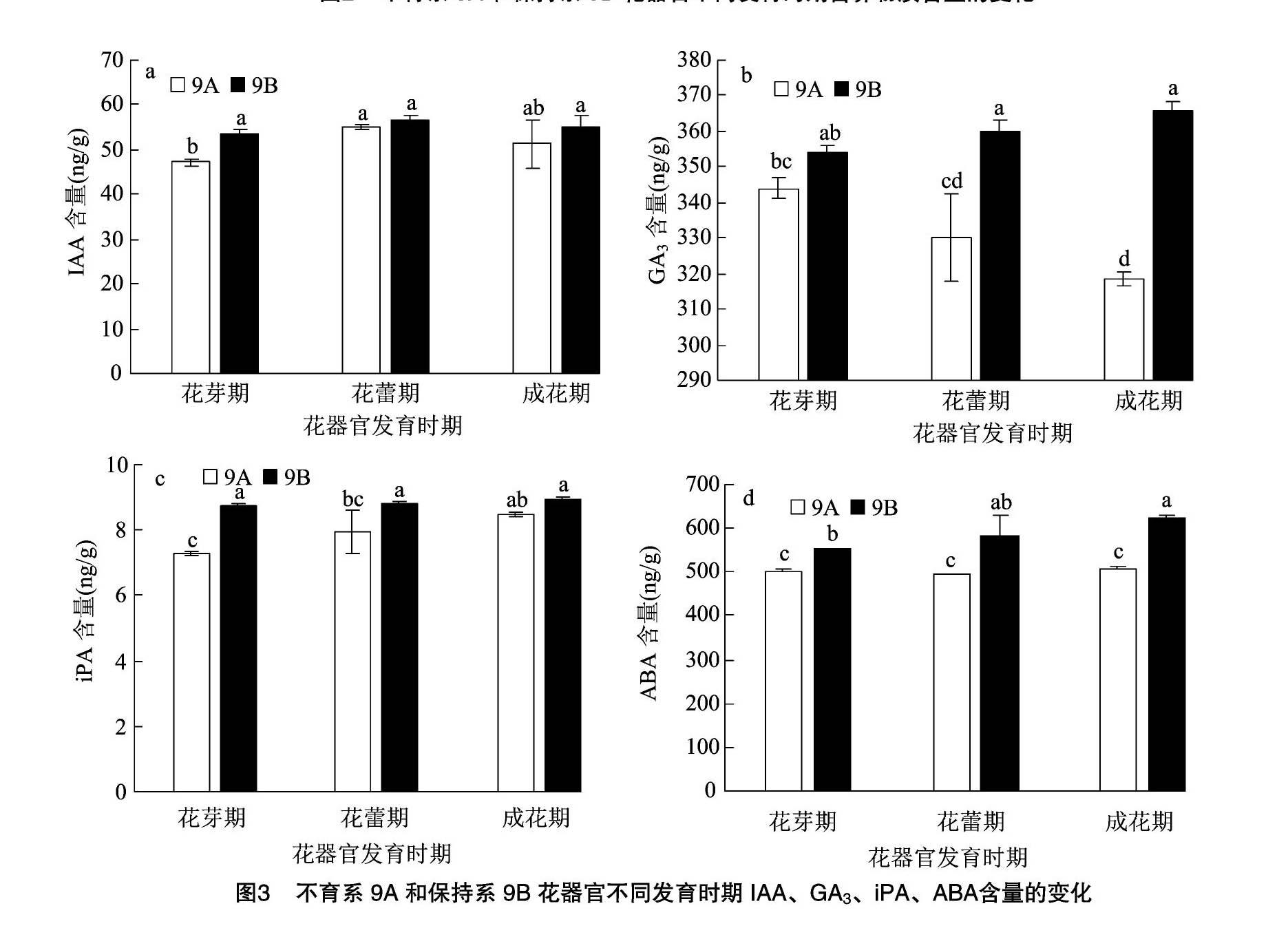
2.3 花器官发育不同时期内源激素含量的变化
如图3所示,不育系9A和保持系9B的IAA含量变化趋势一致,均呈倒U形变化,在花蕾期均达到最大值,分别为55.12、56.73 ng/g,但差异不显著。在花器官发育过程中,9B的IAA含量均高于9A,且在花芽期9B的IAA含量是9A的1.14倍,差异显著;而在花蕾期、成花期均未達到显著性差异。由此推断,花芽期IAA的缺乏可能导致不育的发生。
在花器官发育过程中,不育系9A和保持系9B的GA3含量呈不同的变化趋势,9A呈逐渐降低趋势,9B呈逐渐升高趋势。在花器官发育过程中,9B的GA3含量均高于9A,且在花蕾期、成花期,9B分别是9A的1.09、1.15倍,达到了显著性差异。说明GA3的含量与大豆的育性关系密切,尤其在花器官发育的后期,这可能是不育发生的时期。
不育系9A与保持系9B的iPA含量在花器发育时期均呈逐渐上升趋势,在成花期均达最大值,但是差异不显著;在花芽期和花蕾期均达到了显著性差异,其中,花芽期9B是9A的1.20倍,花蕾期达到1.11倍,说明大豆的雄性不育可能发生在花芽期或花蕾期。
由花芽期至成花期,不育系9A和保持系9B的ABA含量呈完全不同的变化趋势,9A呈U形变化趋势,而9B呈逐渐升高趋势。在花器官发育过程中,9B的ABA含量均显著高于9A,花芽期、花蕾期、成花期9B分别是9A的1.10、1.18、1.23倍,而不育系9A在花器官发育过程中均没有显著性差异。说明ABA含量的积累影响了不育系9A花器官后期的正常发育,从而导致雄性不育,育性的转变影响植物体内ABA含量的积累。
2.4 花器官发育不同时期内源激素之间的平衡关系
由表1可以看出,随着花器官的发育,不育系9A的IAA/ABA的比值先升后降,保持系9B的IAA/ABA比值与9A明显不同,呈逐渐降低趋势。9B的IAA/ABA比值除了在花芽期高于9A,其余2个时期均小于9A。
对于IAA/GA3,不育系9A与保持系9B的比值均在花蕾期达最大值,且二者的变化趋势相同,均为先升后降。花芽期不育系9A的IAA/GA3的值低于保持系9B,花蕾期与成花期不育系9A的 IAA/GA3 的值均高于保持系9B,差距较小,2个时期二者比值均差0.01。
不育系9A与保持系9B在花器官发育过程中IAA/iPA的比值均呈先升后降趋势,且二者在花蕾期的比值明显上升;不育系9A的IAA/iPA的比值在成花期最低,而保持系9B则在花芽期的值最小,不育系9A花芽期与成花期IAA/iPA的值的差距明显,保持系9B的比值的差距较小,为0.02。花芽期与花蕾期不育系9A的IAA/iPA的值明显高于保持系9B,在成花期不育系9A的IAA/iPA的值骤降,明显低于保持系9B。
随着花器官的发育,不育系9A、保持系9B的ABA/GA3的值均在成花期达到最大,分别为1.60、1.71,两者的ABA/GA3的值均随着花器官的发育逐渐升高。各时期内不育系9A的ABA/GA3的值均低于保持系9B。
3 结论与讨论
3.1 抗氧化酶活性与雄性不育的关系
研究报道指出,SOD、POD、CAT具有清除植物体内毒性氧的作用,可以维持植物体内的动态平衡,使植物能进行正常的生理生化活动和正常发育[22-24]。蒋会兵等对厚轴茶不育株的生理生化特性进行研究发现,花蕾发育过程中不育株的POD活性呈上升趋势,均高于可育株,不育株中的SOD、CAT活性先升后降,显著低于可育株[25]。王永琦等研究发现,西瓜不育株中SOD、POD活性均高于可育株,而CAT活性则低于可育株[26]。杨龙树等通过研究大豆不育系的花芽发现,不育系的POD活性显著上升,且显著高于同型保持系[27]。张勤等对玉米不育系研究发现,不育株体内的SOD、POD活性均显著高于可育株,CAT活性低于可育株[28]。本试验结果显示,在大豆花器官发育过程中,不育系的SOD、CAT活性均呈下降趋势,其同型保持系的SOD、CAT活性均呈先升后降趋势,不育系的SOD、CAT活性在花芽期均高于其同型保持系,在花蕾期与盛花期均低于保持系;不育系的POD活性在花芽期低于保持系,在花蕾期、成花期显著高于保持系。该结果与蒋会兵等对厚轴茶的研究结果相似,蒋会兵研究发现在花蕾发育过程中,厚轴茶不育株的POD活性均低于可育株[25]。本研究结果表明,在花蕾期和成花期,不育系的POD活性高于保持系,推测可能与不同作物的不同育性有关,具体原因有待进一步研究。
3.2 营养物质含量与雄性不育的关系
蛋白质、氨基酸、丙二醛、淀粉与糖类作为植物花器官发育过程中的重要物质,对不育系的形成有重要影响,植物花器官中花药与花粉的异常、败育均与此类营养物质缺乏有关[29]。本试验结果表明,大豆不育系的可溶性蛋白含量低于其同型保持系,与冯文鹏等的研究结果[30]一致;游离脯氨酸含量在花芽期高于保持系,而在花蕾期、成花期则是低于保持系,推测可能是与作物类型和取样时期有关。张勤等研究发现,玉米不育株的可溶性糖含量显著低于可育株[28],本试验结果与其一致。王莹等研究发现,紫花苜蓿不育株中的淀粉含量在花蕾发育过程中均低于可育株[31],本试验结果与其一致,说明淀粉含量的亏缺能引起花器官发育异常,从而导致不育。据此可推测,花器官发育过程中淀粉含量亏缺可导致大豆花器官发生败育,从而导致不育的发生。MDA作为脂质发生过氧化的终产物,会引起蛋白质、核酸等生命大分子发生聚合反应,使细胞具有毒性,从而影响花器官的育性[32]。本试验发现,在花芽期、花蕾期、成花期,不育系中MDA的积累量较多,随着花器官的发育先降后升,且均高于保持系;在成花期MDA含量是保持系的1.41倍,说明MDA的含量与花器官的育性密切相关,且高积累量更容易引发花器官的败育。
3.3 内源激素含量与雄性不育的关系
生长素、赤霉素与异戊烯基腺嘌呤核苷均具有促进细胞生长与调控细胞体内养分的作用,脱落酸则会抑制细胞生长,以上几种激素的含量均对植物雄性不育起着重要作用[33]。王永琦等研究发现,西瓜不育株的IAA、GA3、iPA含量均发生严重缺损,在小孢子发育过程中均显著低于可育株[26],在白菜[34]、小麦[35]等雄性不育的研究中也得到了相似的结果。本试验结果与王永琦等的研究结果[26]相似,在大豆花器官的发育过程中,不育系的IAA、GA3、iPA含量均低于其同型保持系,因此导致9A的败育。王学德研究发现,棉花不育株中IAA、GA3含量偏低,而ABA含量偏高[36]。本试验结果表明,不育系体内的ABA含量在各时期内均低于其同型保持系,推测可能与不育发生的时期不同有关。
3.4 激素含量比值与雄性不育的关系
植物体内的激素并不是单独存在的个体,激素之间具有相互促进与拮抗作用,这种激素间的相互作用会导致植物发生不育[37]。本试验结果表明,不育系中IAA/ABA的比值在花器官发育的花蕾期、成花期均高于其同型保持系;在花芽期不育系IAA/GA3的比值比保持系低,在花蕾期、成花期正好相反;在花芽期、花蕾期不育系IAA/iPA的比值均高于保持系,成花期则相反;随花器官的发育,不育系和保持系ABA/GA3的比值均呈递增变化趋势,且不育系均低于保持系。不育系及其同型保持系各时期内ABA/GA3的比值變化均为逐渐上升趋势,且各时期内不育系的比值均低于其同型保持系。孙希禄等对萝卜不育株花蕾激素之间的比值进行分析发现,不育株与可育株的IAA/ABA、IAA/GA3、GA3/ABA之间的比值变化不一致[38]。刘红艳等对芝麻不育株进行研究后发现,其不育株和可育株的IAA/ABA、IAA/SA、IAA/JA比值的变化趋势差异较大[33]。由此可知,激素比值与作物种类有关,不同的作物其比值的变化趋势各不相同。
综上所述,大豆不育系花器官发育过程中SOD、CAT活性比保持系低,而POD活性随花器官的形成先升后降;不育系中MDA积累量较保持系高;可溶性糖、可溶性蛋白、淀粉含量均显著性低于保持系;游离脯氨酸含量先升后降;不育系的IAA、GA3、iPA、ABA含量均显著低于保持系,且激素比值差异较大,引起激素平衡失调。因此,不育系9A的败育与花器官发育过程中激素含量的缺失有关,据此推测以上结果受到大豆质核互作雄性不育的影响。花器官发育至花蕾期后,SOD、CAT活性,MDA、淀粉、可溶性糖、游离脯氨酸、GA3、ABA含量等多项生理生化指标在不育系9A与保持系9B间差距较大,花蕾期可能是大豆质核互作雄性不育系生理生化指标发生异常的关键时期。本研究结果仅反映了大豆质核互作雄性不育系花器官发育过程中的生理生化变化趋势,旨在为进一步研究大豆质核互作雄性不育的遗传机制提供理论基础。
参考文献:
[1]王曙明,王跃强,李建平,等.田间开放条件下大豆不育系制种技术研究[J]. 大豆科学,2010,29(3):385-389.
[2]马小定,邢朝柱.棉花雄性不育研究和应用进展[J]. 棉花学报,2006,18(5):309-314.
[3]陈 秒,尹 超,刘 烨,等.水稻温敏核不育系育性转换因素及生理生化特性[J]. 作物研究,2013,27(2):164-168.
[4]鲁美宏,孙万仓,孔德晶,等.白菜型冬油菜不育系LRCMS花器生理生化特性及其雄蕊发育特征研究[J]. 西北植物学报,2014,34(3):509-515.
[5]刘齐元,刘飞虎,何宽信,等.烟草雄性不育的细胞形态学及生理生化研究进展[J]. 亚热带植物科学,2004,33(1):69-72.
[6]汤伟华,张蜀宁,孔艳娥,等.不同倍性不结球白菜Pol CMS及保持系生理生化特性比较[J]. 西北植物学报,2009,29(1):80-84.
[7]孔艳娥,张红梅,韩业飞,等.茎芥菜胞质四倍体白菜雄性不育系及保持系生理生化特性研究[J]. 江西农业学报,2011,23(7):12-15.
[8]王书强,崔辉梅,石国亮.大白菜Ogura雄性不育系及保持系生理生化分析[J]. 新疆农业科学,2011,48(1):110-115.
[9]邹 佳,蔺万煌,罗红兵,等.玉米C型胞质雄性不育系POD、CAT、SOD活性及POD酶谱分析[J]. 玉米科学,2009,17(6):45-49.
[10]胡小娟.几种活性物质与超氧化物歧化酶相互作用的研究[D]. 合肥:安徽大学,2012.
[11]冯小磊,范光宇,苏 旭,等.植物雄性不育生理生化研究进展[J]. 作物杂志,2012(3):6-11.
[12]何长征,萧浪涛,刘志敏,等.植物激素与雄性不育关系的研究进展[J]. 中国农学通报,2002,18(3):65-69.
[13]余建章,荐 立.沈农雄性核不育大豆 L-78-387等位性测验研究[J]. 沈阳农学院学报,1985,16(4):19-24.
[14]杨守萍,曾维英,段美萍,等.大豆雄性不育突变体NJ89-1核不育基因的SSR标记和定位[J]. 大豆科学,2006,25(4):344-348.
[15]赵团结,盖钧镒.大豆不育性自然变异的发现与鉴定[J]. 中国农业科学,2006,39(9):1756-1764.
[16]卫保国.大豆光温敏感型雄性不育系发现初报[J]. 作物品种资源,1991(3):12.
[17]彭玉华,杨国保,袁建中,等.一个对播种期反应敏感的不育大豆特征分析[J]. 作物学报,1998,24(6):1010-1013.
[18]王 芳,卫保国,李贵全,等.大豆光敏雄性不育株88-428BY-827小孢子母细胞的细胞学观察[J]. 中国农业科学,2004,37(8):1110-1113,1251.
[19]孙 寰,赵丽梅,黄 梅.大豆质-核互作不育系研究[J]. 科学通报,1993,38(16):1535-1536.
[20]张 磊,戴瓯和.大豆质核互作不育系W931A的选育研究[J]. 中国农业科学,1997,30(6):90-91.
[21]盖钧镒,丁德荣,崔章林,等.大豆质核互作雄性不育系NJCM S1A的选育及其特性[J]. 中国农业科学,1999,32(5):23-27.
[22]张文静,刘 亮,黄正来,等.低温胁迫对稻茬小麦根系抗氧化酶活性及内源激素含量的影响[J]. 麦类作物学报,2016,36(4):501-506.
[23]刘淑娟,朱 祺,幸学俊,等.植物雄性不育影响因素研究进展[J]. 中国农学通报,2014,30(34):46-50.
[24]O’Brien J A,Daudi A,Butt V S,et al.Reactive oxygen species and their role in plant defence and cell wall metabolism[J]. Planta,2012,236(3):765-779.
[25]蒋会兵,许 燕,宋维希,等.厚轴茶雄性不育株花蕾发育过程中的生理生化变化[J]. 植物生理学报,2020,56(9):1807-1817.
[26]王永琦,杨小振,莫言玲,等.西瓜雄性不育系‘Se18’抗氧化酶活性和内源激素含量变化分析[J]. 园艺学报,2016,43(11):2161-2172.
[27]杨龙树,李佳佳,贺亭亭,等.大豆质核互作雄性不育系NJCMS1A与其保持系NJCMS1B的生理生化特性比较分析[J]. 大豆科学,2017,36(3):391-398.
[28]张 勤,金圣浩,方 芳,等.玉米光温敏雄性不育系CB1208-82的生理生化代谢研究[J]. 玉米科学,2019,27(3):48-53.
[29]盛云燕,常 薇,矫士琦,等.甜瓜雄性不育植株雄蕊发育结构及生理生化特征[J]. 植物生理学报,2016,52(7):1028-1034.
[30]冯文鹏,周书栋,杨博智,等.辣椒反向温敏雄性核不育突变体E6421S不育特征及遗传规律研究[J]. 园艺学报,2019,46(6):1112-1122.
[31]王 莹,王英哲,徐安凱,等.紫花苜蓿细胞质雄性不育系及其保持系花蕾的生理生化特性[J]. 中国草地学报,2018,40(1):24-28,34.
[32]Zheng B B,Fang Y N,Pan Z Y,et al.iTRAQ-based quantitative proteomics analysis revealed alterations of carbohydrate metabolism pathways and mitochondrial proteins in a male sterile cybrid pummelo[J]. Journal of Proteome Research,2014,13(6):2998-3015.
[33]刘红艳,吴 坤,杨敏敏,等.芝麻显性细胞核雄性不育系内源激素、可溶性糖和淀粉含量变化[J]. 中国油料作物学报,2014,36(2):175-180.
[34]王永勤.白菜核不育两用系育性基因表达特征分析及其小孢子发育相关基因的分离[D]. 杭州:浙江大学,2003.
[35]张自刚,马小飞,张红霞,等.小麦光温敏雄性不育系BNS育性转换与内源激素的关系研究[J]. 植物遗传资源学报,2016,17(5):913-919.
[36]王学德.棉花细胞质雄性不育的研究与利用[J]. 中国农业科学,2019,52(8):1341-1354.
[37]丛 庆,张 琪,宋丽莉,等.激素在植物冷胁迫应答中的角色[J]. 核农学报,2016,30(3):614-619.
[38]孙希禄,许小勇,张鲁刚.萝卜雄性不育系花蕾发育过程中内源激素分析[J]. 北方园艺,2011(19):11-15.
