林下引入耐阴树种功能性状特征及其与生长的关系
2020-11-14徐海东查美琴成向荣周恒发虞木奎
徐海东,查美琴,成向荣,汤 胜,周恒发,虞木奎,*
1 中国林业科学研究院亚热带林业研究所,杭州 311400 2 永丰县官山林场,江西 永丰 331500
我国人工林面积达6933万hm2,占有林地面积的36.3%,其中杉木(Cunninghamialanceolata(Lamb.)Hook.)人工林占全国人工林面积的19%[1]。杉木作为亚热带地区重要的速生造林树种,发挥了重要的生态和经济效益。长期以来,杉木人工林以纯林经营为主,往往造成土壤酸化和林分生产力的降低,而在纯林内引入常绿阔叶树种构建复层异龄林,对改善土壤肥力,提高林分稳定性具有重要意义[2-3]。植物功能性状是影响植物生长、存活、繁殖能力和最终适合度的一系列可测量的生态功能指标,也是反映环境变化对植物生长发育影响的重要指示指标[4]。根、茎、叶是高等植物主要的构件系统,根茎叶间营养元素分配与形态结构的差异是体现植物功能性状变化最重要的方面[5]。叶是能量生产和物质积累的重要器官,叶面积、比叶面积、单位叶面积氮含量等叶性状间的关联可反映植物适应环境变化所形成的生存对策。例如,比叶面积与单位叶面积的氮含量一般成正比,而与叶寿命均成反比,体现了植物性状对环境的趋同适应策略[6];茎是运输根、叶之间水分和养分的枢纽,并支撑着植株的向上生长,而茎为了抵抗外界环境的干扰往往会提高自身木质密度以及碳含量[7];叶和茎是植物体地上部分的重要营养器官,根作为重要的地下器官,尤其细根是根系吸收土壤水分和养分的关键区域,细根养分元素以及生物量分配对植物生长发育也起到重要作用[8]。此外,关于三者之间功能性状的协调与联系主要有3个假说[9]:(1)功能相似假说。叶与根系性状紧密关联,与茎性状关联较少,因为根系对水分和养分的吸收决定了叶功能的正常运行,而叶为根系提供源源不断的养分物质。(2)结构相似假说。木本植物茎和根系统的木质部和韧皮部的解剖结构一致,因此,茎和根干物质含量、组织密度等性状相似性更高。(3)整体协调假说。植物根茎叶的功能协调合作,共同维持植物体功能系统的运行,即根茎叶之间对应的功能性状也是紧密关联的。因此,植物根茎叶功能性状特征及其协同关系可充分反映植物对环境的响应以及资源的利用。目前,多数研究集中于植物叶功能性状特征或者叶和根功能性状间的协调性[10-12],对于根、茎功能性状特征以及根茎叶性状关联性的研究较少,尤其关于林下耐阴树种根茎叶功能性状及其关联性的研究则更为少见。
植物功能性状与生长率关系紧密,植物种间和种内功能性状的变异是物种共存的基础,因此,种间、种内功能性状与生长率之间的联系有助于我们更好地了解树种生存策略以及预测树种特异性生长策略的动态变化[13]。构建异龄复层林迫切需要了解引入树种功能性状对植物生长指标的驱动机制,植物根茎叶功能性状的早期表现可能有助于更好的筛选林下引入树种。有研究报道幼树功能性状比成年树能更好的预测植物生长率[14],而幼树的生长及生存状况是林下引入树种构建异龄复层林成败的一个关键阶段。明确林下引入树种功能性状与其生长指标关联性,筛选影响植物生长的关键功能指标,对于构建稳定复层林生态系统具有重要意义。迄今为止,许多研究集中在植物单一功能性状对生长率的预测,尤其关于生长率与叶片功能性状的研究较多[15-16],而关于林下引入耐阴树种根茎叶功能性状如何影响其生长率的研究仍非常贫乏。为此,我们重点研究了杉木人工林林下引入的5种常绿阔叶树种根茎叶功能性状特性及其对树种生长率的影响,并提出以下问题:(1)耐阴树种根茎叶功能性状之间是否存在显著的耦合关系?(2)耐阴树种种间和种内关键性状与生长率的相关性是否一致?(3)叶性状是否可以作为林下引入树种早期生长率变化的关键指标?通过解决以上问题,以期为亚热带杉木纯林人促更新技术、构建杉木复层林树种选择提供理论支撑和实践参考。
1 材料与方法
1.1 试验区概况
研究区位于浙江省开化县林场城关分场小桥头林区(118°25′E,29°09′N)。该区年日照总时数1334.1 h,年平均气温16.4 ℃,年平均降雨量1814 mm,无霜期252 d,气候属亚热带季风气候。供试杉木纯林均为1996年春季造林,初植密度2500株/hm2,2006年进行1次抚育间伐,目前保留密度1358株/hm2,林分郁闭度约0.85。土壤类型为红黄壤,pH为4.0—4.9。
1.2 试验方法
2011年3月,在小桥头林区选取间伐后生长和立地条件基本一致的杉木人工林,将引入的5种生态效益和经济价值较高的耐阴常绿阔叶树种[17](分别是:洒金叶珊瑚,Aucuba japonica var.variegata;杨桐,Adinandramillettii(Hook. et Arn.) Benth. et Hook. f. ex Hance;蚊母树,DistyliumracemosumSieb.et Zucc.;栀子,GardeniajasminoidesEllis;海桐,Pittosporumtobira(Thunb.) Ait.)沿等高线块状栽植在杉木林下,每个树种栽植面积约0.5hm2,栽植密度3000株/hm2,栽植苗木均为1年生健壮容器苗。在每个林分内随机布设3块10 m × 20 m的固定样地,定期调查林木生长状况。引入树种存活率在85%以上,树种栽植时树高(H0)、地径(B0)和30个月后树高(H1)、地径(B1)的统计数据见表1。
1.3 根茎叶性状测定和生长率计算
2013年9月从每个树种选择3个平均样株测定根茎叶形态和生物量等指标,把植株的根茎叶等器官分离并测定鲜重,每个样株随机选择20—30个完整叶片用LI—8400plus叶面积仪测定叶面积(Leaf area,LA),并挑选细根(< 2mm的根系)用Epson根系扫描仪测定根长(Root length,RL)、根体积(Root volume,Rv)和根表面积(Root surface area,RSA),LA和RL、Rv、RSA分别采用Win FOLIA叶面积和WinRHIZO根系分析系统确定。随后将已测量的叶片和根系样品以及未测量样品分别置于75℃烘箱烘至恒重(约为72 h),并测定干重。以下指标均由计算得出:比叶面积(Specific leaf area,SLA)=扫描叶面积/扫描叶干重;比根长(Specific root length,SRL)=细根长/细根干重;比根面积(Specific root area,SRA)=细根表面积/细根干重;根组织密度(Root tissue density,RTD)=细根干重/细根体积;叶干物质含量(Leaf dry matter content,LDMC)=叶片干重/叶片鲜重;茎干物质含量(Stem dry matter content,SDMC)=茎干重/茎鲜重;根干物质含量(Root dry matter content,RDMC)=根干重/根鲜重;叶生物量占比(Leaf biomass fraction,LBF)=叶生物量/林木单株总生物量;茎生物量占比(Stem biomass fraction,SBF)=茎生物量/林木单株总生物量;根生物量占比(Root biomass fraction,RBF)=根生物量/林木单株总生物量。树高相对增长率计算公式:RGR-H=(lnH1-lnH0)/(t1-t0);地径相对增长率计算公式:RGR-B=(lnB1-lnB0)/(t1-t0),H0和B0分别是栽植时树高和地径,H1和B1分别是栽植30个月后的树高和地径,t1-t0为栽植前后时间间隔。
根茎叶碳(C)含量采用重铬酸钾氧化-外加热法测定;根茎叶氮(N)含量用浓硫酸-过氧化氢消煮-半微量凯氏定氮法测定;根茎叶磷(P)含量采用钼锑抗比色法测定;根茎叶钾(K)含量采用火焰光度计法[18]。
1.4 数据处理
采用Excel 2013对数据进行整理与计算。首先,进行单因素方差分析(One-way ANOVA),采用最小显著差异法(LSD)对不同耐阴树种同一功能性状指标的比较(显著性水平α=0.05);其次,利用“FactoMineR”和“corrplot”包分别进行主成分分析和皮尔逊相关性分析。对不同树种生活型的划分以及根茎叶功能性状之间的协调性进行主成分分析,根茎叶功能性状与生长率的相关性分析采用皮尔逊相关法进行;最后,根茎叶关键性状与生长率在种间和种内的关联性网络分析采用“qgraph”包完成,性状网络中心性由度(Degree,表示一个节点的所有边数之和)和加权度(Weighted degree,一个节点的所有显著相关系数的和)2个指标衡量[19],上述分析均用R(Version 3.5.1)完成。图表中数据为平均值±标准差(Mean±SD)。
2 结果与分析
2.1 生长率和功能性状特征
如表1所示,树高平均生长率(RGR-H,0.022 cm cm-1月-1)高于地径生长率(RGR-B,0.016 mm mm-1月-1)。其中,树高在30个月后增长了1.5—3.5倍,地径增长了1.5—1.8倍。洒金叶珊瑚的树高生长率以及洒金叶珊瑚和栀子的地径生长率显著高于其余树种(P< 0.05)。
耐阴树种种间和种内的根茎叶功能性状存在不同程度的变异,种内变异系数普遍低于种间变异系数(表2)。种间根、茎、叶性状的平均变异系数分别为24.86%、20.02%和32.63%,而种内所有根、茎、叶性状的平均变异系数均未超过10%;种间根茎叶各性状变异范围差异较大,根茎叶碳含量种间变异系数均未超过10%,而叶面积、叶钾含量、茎钾含量和根长变异系数均超过50%,其中叶面积具有最高变异系数(91.99%);根茎叶各性状值差异也较大,平均叶生物量占比(0.48)和茎生物量占比(0.47)高于根物量占比(0.25),而平均叶干物质含量(0.32 g/g)小于根和茎的干物质含量(分别为0.37 g/g和0.47 g/g)。根茎叶碳含量差异不大,但叶氮磷钾含量远高于根和茎的氮磷钾含量,尤其叶氮含量分别是根和茎氮含量的2.0和2.6倍。此外,叶N/P比值(16.91—27.49)也高于根和茎(10.76—16.89)。
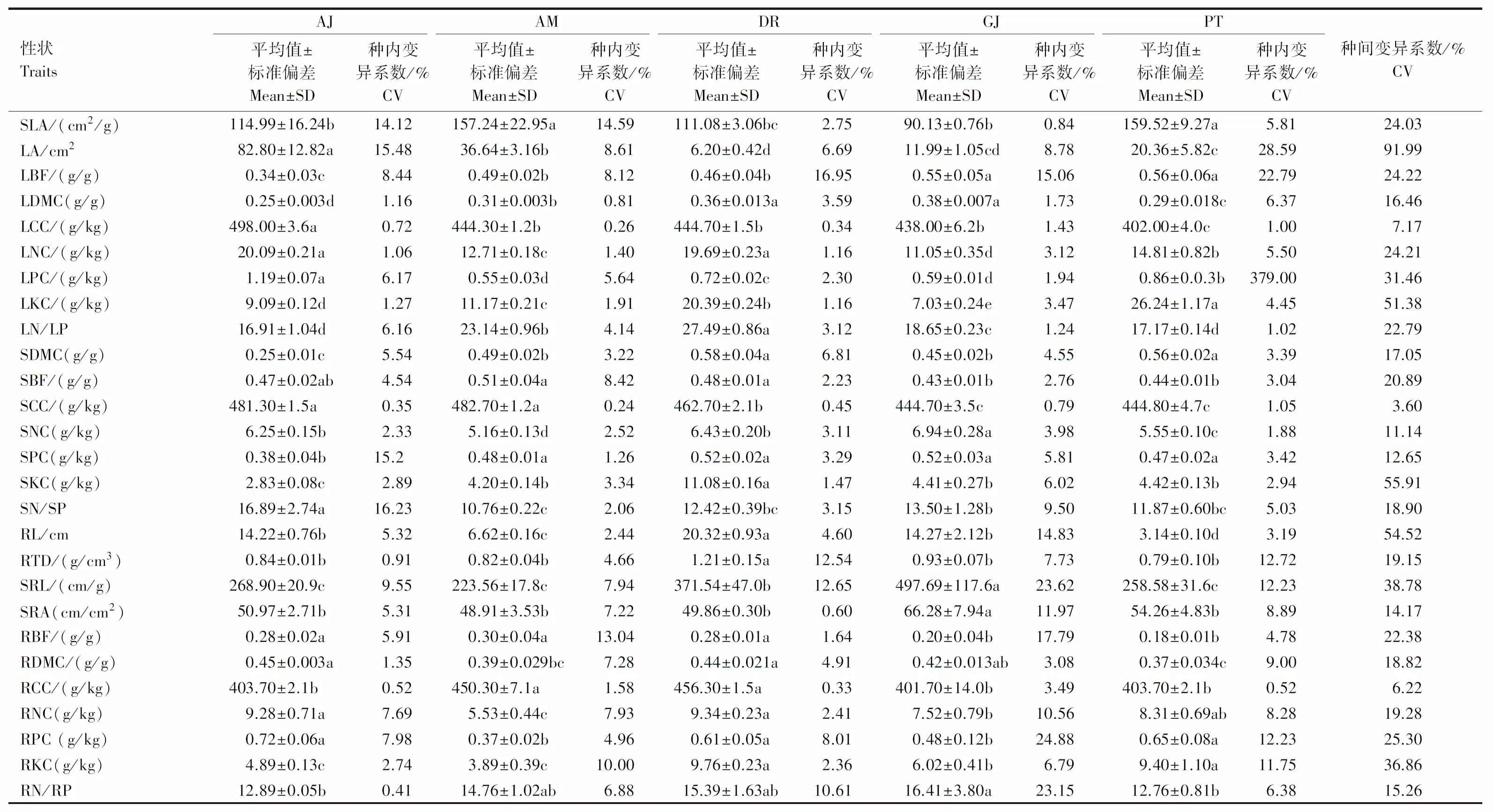
表2 耐阴树种性状值及种间和种内变异系数Table 2 Trait values and inter-specific, intra-specific coefficient variation of shade-tolerant tree species
2.2 功能性状分布与关键性状
对5个耐阴树种功能性状和生长率的主成分分析发现,叶性状主要分布在主轴左侧,根茎性状集中分布在右侧,且根茎功能性状关联更为紧密。此外,洒金叶珊瑚分布在主轴左侧,具有较高的生长率、叶面积、叶生物量占比和较低的根茎叶干物质含量,而其余树种均分布在主轴右侧;基于皮尔逊相关性分析发现,树高和地径相对增长率与叶面积、叶生物量占比、叶碳含量、茎氮磷比以及根氮含量均显著正相关,与茎生物量占比、茎干物质含量、茎碳含量、根碳含量以及根干物质含量均显著负相关(图1),本文将与树高和地径相对增长率均显著相关的10个功能性状作为预测生长率的关键性状进行后续的网络关联性分析(图2)。
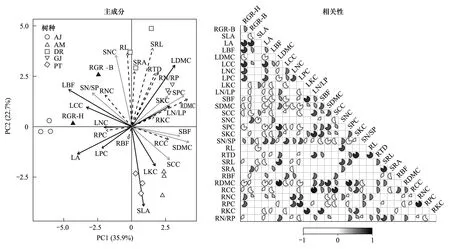
图1 耐阴树种功能性状和生长率主成分及相关性分析Fig.1 Principal component analysis and correlation between functional traits and growth rates in shade-tolerant tree species黑色实线表示叶性状,灰色实线表示茎性状,虚线表示根性状;AJ:洒金叶珊瑚;AM:杨桐;DR:蚊母树;GJ:栀子;PT:海桐;图中黑色扇形表示显著正相关,白色扇形表示显著负相关(P < 0.05),空白处表示不显著(P > 0.05);SLA:比叶面积;LA:叶面积;LBF:叶生物量占比;LDMC:叶干物质含量;LCC:叶碳含量;LNC:叶氮含量;LPC:叶磷含量;LKC:叶钾含量;LN/LP:叶氮磷比;SDMC:茎干物质含量;SBF:茎生物量占比;SCC:茎碳含量;SNC:茎氮含量;SPC:茎磷含量;SKC:茎钾含量;SN/SP:茎氮磷比;RL:根长;RTD:根组织密度;SRL:比根长;SRA:比根面积;RBF:根生物量占比;RDMC:根干物质含量;RCC:根碳含量;RNC:根氮含量;RPC:根磷含量;RKC:根钾含量;RN/RP:根氮磷比
2.3 种间、种内关键性状和生长率相关性
耐阴树种生长率与10个关键功能性状在种间和种内关系网中表现出较大差异(图2)。在种内(个体)水平上,叶生物量占比(除洒金叶珊瑚外)和叶面积与树高相对增长率均显著正相关(P< 0.05),而其余关键性状与树高和地径相对增长率的网络关联性较为复杂;在种间(所有物种)水平上,树高和地径相对增长率与叶面积和叶生物量占比表现出强烈的正相关,而与根茎干物质含量和根茎碳含量呈较强的负相关(P< 0.05)。此外,根和茎性状与大部分叶性状呈负相关,而大多数根与茎性状呈正相关(图2)。
由图2和表3可知,在种内关系网中,根干物质含量、茎生物量占比和叶生物量占比分别是洒金叶珊瑚、杨桐和蚊母树、栀子和海桐中心性最高的关键性状;在种间关系网中,中心性最高的性状是叶面积。与地径相对增长率(平均权重和节点数分别为6.52和8.17)相比,树高相对增长率无论在种间还是种内均具有较高的网络权重值(平均权重为7.75)和网络节点数(平均节点数为9.5)。
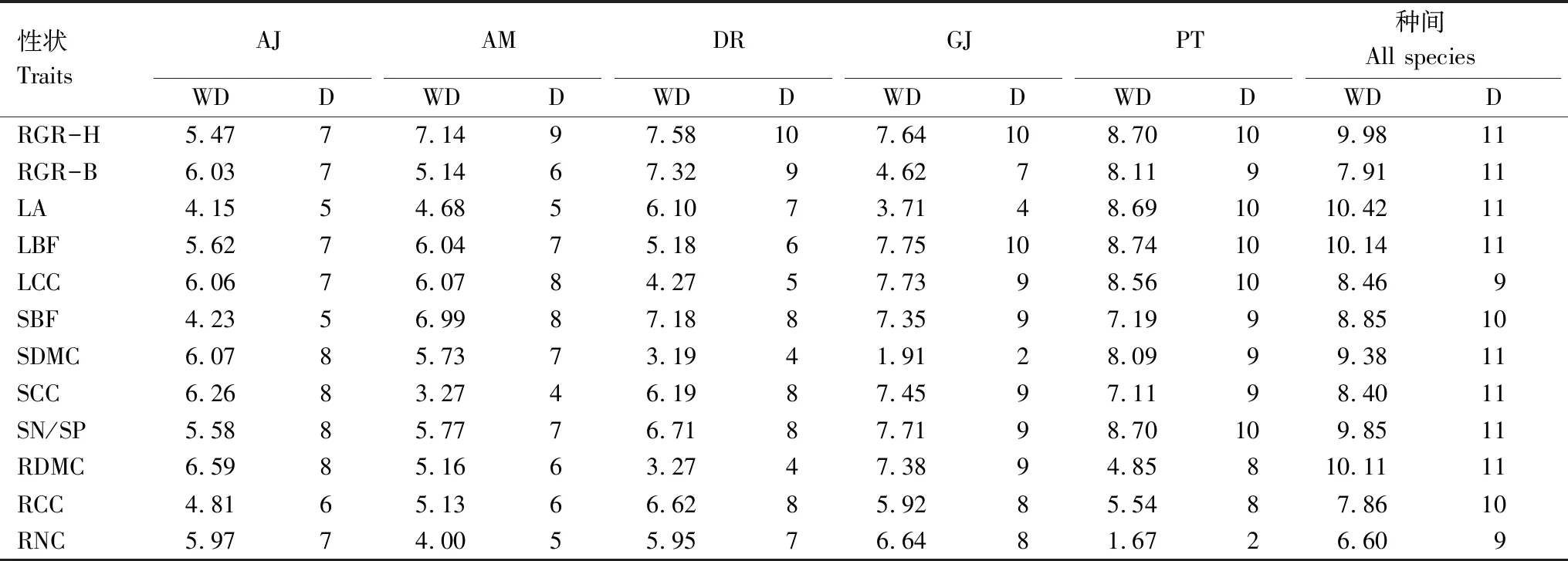
表3 种间和种内性状与生长率关系网的节点和权重Table 3 Network nodes and weights of relationship between inter-and intra-specific traits and relative growth rate
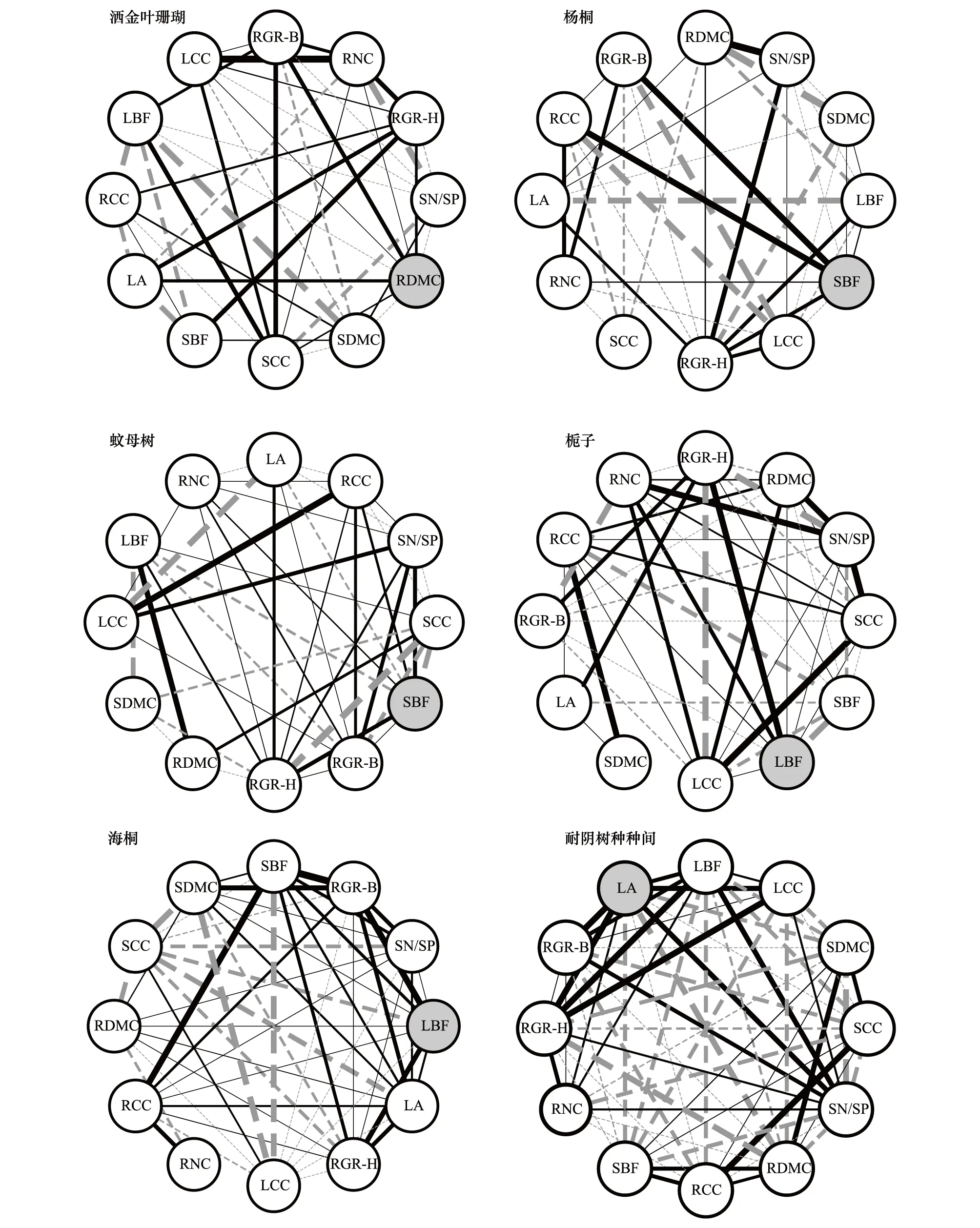
图2 耐阴树种种间和种内关键性状与生长率关系网Fig.2 The correlation network between inter-and intra-specific key traits and relative growth rate of shade-tolerant tree species图中实线表示显著正相关,虚线表示显著负相关(P < 0.05),无连接线表示相关性不显著(P > 0.05),灰色背景圆圈表示中心性最高的关键性状指标
3 讨论
3.1 耐阴树种性状特性和主要适应策略
植物功能性状的变异特征及性状间协调关系是物种共存的基础,尤其性状之间的权衡和协同更有助于我们了解树种对生态过程的自我调节和适应机制[20-21]。本研究发现,耐阴树种种间叶性状的平均变异系数高于根茎功能性状,说明这些耐阴树种叶片可塑性更强,通过改变个体叶性状(尤其是叶面积)来更好的适应林下多变的环境以及更充分的利用混交林有限的空间资源,这与Garnier等[22的研究结果一致。有研究报道植物长期适应于复杂土壤环境会导致根性状变异系数偏大[23-24],本研究却发现种间根性状平均变异系数小于叶性状,这可能与林下弱光环境有关。因为光是影响林下植被生长和发育的主要限制因素,叶片作为光合作用的主要器官对光环境的变化非常敏感,通常植物通过叶片的形态、生理和解剖结构等可塑性来适应林下光环境变化,而根系可能对水分、养分等环境因素的变化更为敏感[25-26]。相比根叶性状,茎主要起支撑与运输作用,性状变异较小[7]。此外,植物功能性状之间的权衡关系有助于我们充分了解耐阴植物的生活史对策[27]。根茎叶生物量分配不仅反映植物适应特定环境的构型塑造,也是研究植物生活史对策的重要内容。多数研究发现光照资源有限的条件下植物叶的生物量分配会更高,并以此获取更高的光合效益[28-29],本研究同样发现叶生物量分配均高于根和茎。因为在高郁闭度杉木林下,光照是限制植物生长的主要因子,植物为了获取更高的光合效益,通常采取增大获取限制性资源的器官分配,减少获取非限制资源的器官分配的策略[30-31]。此外,耐阴树种根茎叶碳含量分配较一致,但叶片氮磷钾含量远大于根和茎。其中,叶片氮和磷含量的提高会促进叶片光合作用[32],这也是林下树种提高光合能力的重要生态策略。目前,根茎叶功能性状之间存在“整体协调假说”、“功能相似假说”和“结构相似假说”,本文基于主成分分析发现耐阴树种根和茎功能性状相关性更为紧密,比较符合结构相似假说[33]。此外,耐阴树种比叶面积和比根长、比根面积及根长之间均表现为显著负相关(图1C),表明光照不足环境下植物会分配给叶更多的资源以提高光能利用率,而不是投入给细根的生长,这在一定程度上也不支持功能相似假说以及整体协调假说。氮和磷是植物生长不可或缺的营养元素,本文发现叶片氮磷和细根氮磷之间均存在显著正相关,这与之前多数研究结果一致[34-35],说明耐阴树种氮磷元素在叶和细根之间可能也遵循一定的分配机制。此外,植物叶片N/P比值的阈值(14和16)影响植物的生长,当N/P < 14时,氮是最主要的限制因子,N/P > 16时,磷是最主要的限制因子[36],本文发现所有树种叶片N/P > 16,说明林下引入的耐阴树种可能主要受磷限制。因此,引入树种时也应考虑磷利用效率较高的耐阴植物。
3.2 耐阴树种关键性状与生长率关系网
性状关系网络能够直观的表现植物性状在种间和种内的多元关系,对评价植物可塑性及其生态适应策略提供了更加准确的方法[19]。植物性状间的组合,使植物能够充分利用所获资源,以达到其自身生长最大化的目的。已有研究关注根茎叶中单一性状与生长率相关性的分析,这忽视了不同营养器官的性状组合对植物生长的影响[15-16],本文在种间和种内水平上将耐阴树种根茎叶功能性状与树高和地径相对增长率关联性进行全面分析。观察发现,与生长率显著相关的种间关键性状在种内水平上与生长率表现出更为复杂的关系网,说明耐阴树种可通过自身较高的性状可塑性以及多元的适应策略来弥补有限资源环境下的生长。基于主成分分析发现,耐阴树种可明显分为两个功能群落,以杨桐、蚊母树、栀子、海桐为主的“硬叶植物”(表现为叶干物质含量高、叶生物量占比及叶面积小)和洒金叶珊瑚类型的“薄叶植物”(叶干物质含量低、叶生物量占比及叶面积大)。有研究报道薄叶植物普遍具有较高的养分吸收速率和生长率[5],本文同样发现,与硬叶植物相比,洒金叶珊瑚具有更高的氮磷含量和生长率,说明在弱光环境下薄叶植物可能仍表现出较高的生长率。林下树种为提高生长率可能会采取不同的生态策略。例如,洒金叶珊瑚种内关系网中表现出较高中心性的性状是根干物质含量和茎碳含量(表2),两者含量的增加会提高植物根茎的物理支持以及水分、养分运输的安全性,尤其有助于支撑较大叶面积的薄叶植物,从而加强了植物叶光合作用的稳定运行和生长率的提高;茎生物量占比是杨桐和蚊母树关系网中心性最高的性状,说明杨桐和蚊母树为了提高生长率,可能采取分配茎更多生物量的生态策略,而茎生物量分配的增加会增强其顶端优势[37];而栀子和海桐为适应弱光环境下的生长,通过增大对最高中心性性状(叶生物量占比)的投入来提高光合效能和生长率[38]。基于性状网络关联性分析,本文还发现树高相对增长率的中心性远高于地径相对增长率,说明树高相对增长率可作为衡量耐阴植物生长率的主要指标。此外,为了提高功能性状对生长率预测的准确性,对耐阴种间和种内关键性状与生长率关系网进行了全面评估,我们发现10个关键性状中只有叶面积是种间和种内关系网中均与树高相对增长率显著正相关的指标。说明叶面积更适合作为指示林下引入常绿阔叶树种生长率变化的关键性状,这可能主要归因于大叶片物种在阴暗环境下往往具有较高的光合效益[39],从而导致叶面积对生长率的贡献更高。由于本研究仅是初步结论,引入树种在杉木林下生境的长期生长表现还有待进一步研究。鉴于本研究区立地条件和环境因子较一致的前提下,耐阴树种的功能性状更能准确反映其生长的差异,但相应结论也需要对更多林下树种及更大尺度范围进行验证。
4 结论
本文重点分析了林下引入耐阴树种根茎叶功能性状特征及其与生长率关系,并筛选关键功能性状,探讨了种间和种内关键性状与生长率的关系,主要结论如下:
(1)根茎功能性状相关性更紧密,但种间根茎性状平均变异系数要小于叶性状,即叶性状的可塑性更强。
(2)种间和种内关键性状与生长率关系差异较大。叶面积是种间中心性最高的性状,根干物质含量、茎和叶生物量占比是种内中心性最高的性状。此外,树高相对增长率的中心性远高于地径相对增长率。
(3)叶面积与树高相对增长率的显著正相关是种间和种内均存在的唯一关系。综上所述,为提高功能性状对生长率预测的准确性,需要我们综合考虑种间和种内功能性状和生长率的协调关系。本文发现耐阴树种叶面积在种间、种内均显著关联早期生长率,因此,具有较大叶面积的耐阴树种(如洒金叶珊瑚、草珊瑚等)可作为林下引入树种的首选,这对高效构建复层林提供了理论依据和实践参考。
