合成生物学在生物基塑料制造中的应用
2020-10-27徐彦芹杨锡智罗若诗黄玉红霍锋王丹
徐彦芹,杨锡智,罗若诗,黄玉红,霍锋,王丹
(1 重庆大学化学化工学院,化工过程强化与反应国家地方联合工程实验室,重庆400044;2中国科学院过程工程研究所,北京100190)
引 言
合成生物学(syntheticbiology)是一门颠覆性的新兴交叉学科,使人类能够实现像组装机器一样组配生物,是生命科学领域的一次伟大革命。合成生物学是在现代生物学和系统科学基础上,融入工程学思想,将分子生物学、计算机、工程技术和信息技术等综合应用的多学科交叉领域。它通过设计、构建、调试、优化的工程学循环思想,改造或创造出新的生物合成途径。合成生物学已经应用于生物能源[1−5]、环境保护[6−12]、生物医药[13]、食品科学[14−20]和大宗化学品制造[21−28]等领域。目前,合成生物学已经成为全球各国政府和生物科技公司重点关注、投入研发的热点之一。塑料制品与人们生产、生活息息相关,广泛应用于医疗制品、食品包装、汽车和通讯等行业。而在航空、航天、通讯工程和军事领域,工程塑料作为金属的替代品,可广泛地作为工程结构材料使用,在苛刻的化学、物理环境中展现出优良的综合性能。现在使用的塑料制品大部分是由石油化工生产的石油基塑料,随着石油资源逐步匮乏和突出的环保问题,以可再生的天然生物质资源为原料,通过合成生物学构建工程细菌或工程细胞来生产生物基塑料(表1),具有愈加广阔的市场应用前景[39−42]。
1 合成生物学的发展现状
1.1 合成生物学概念的提出
1910 年,合成生物学概念由法国科学家Stéphane Leduc 提出,并且预见了生命科学由描述、分析到合成的发展方向;1965 年,国际上首次人工合成具有生理活性的蛋白质牛胰岛素由我国科学家完成,是生命科学的里程碑事件;1974 年,波兰科学家Waclaw Szybalski 提出将“应用”注入合成生物学概念中;1980年,Hobom B提出合成生物学表述为基因重组技术;2003 年,国际上定义合成生物学为基于系统生物学的遗传工程和工程方法的人工生物系统研究。
合成生物学与集成型建筑工程类似,采用工程化设计理念,工程学模块化思想、系统设计理论,通过自动化和标准化的过程,改造或重建天然的生物体系,最终构建出具有特定功能的可控生命体系[43−45]。21 世纪初,合成生物学突破了数年的自然进化限制,开启了从深入认识和探索生命到设计和创造生命的新篇章,实现了所需品的可控定量表达与生产。因此,当人类面临能源、资源和环境等重大问题时,合成生物学可以为其提供新的解决途径,目前合成生物学已经在人类生产生活中实现了广泛的应用(图1)[46−47]。
1.2 合成生物学的重要技术
1.2.1 生物传感器 合成生物学的发展让人们在细胞天然传感的基础上可以进行构建、加工生物传感器,用来感应细胞内目标靶向代谢产物的变化,实现特定信号的感应和输出。而人工构建的生物传感器又反过来作为一种常规化的技术,指导人们进行代谢途径的构建、产物产量提高及工程酶的筛选。生物传感器主要由生物识别元件和信号输出元件组成。生物识别元件包括核糖开关、转录因子、核糖体干扰肽和压力响应启动子等。信号输出元件感受到识别元件内部的信号变化,以荧光信号或者代谢通路的开闭等不同的形式输出[48]。生物传感器在合成生物学中的应用:例如高通量筛选高产量目标产物的菌株,动态调控代谢物在细胞内的水平等。Binder 等[49]成功设计和开发出用于细胞内检测碱性氨基酸、L−丝氨酸和O−乙酰丝氨酸的传感器。首先,设计出合适的代谢物传感器,在携带传感器的细胞基因组中产生遗传多样性,通过FACS筛选突变文库和选择单个产生细胞,验证和鉴定突变体。通过从随机诱变野生型(WT)细胞库中分离获得产生L−赖氨酸的细菌,对目标细菌测序鉴定,证实了该方法的可行性。
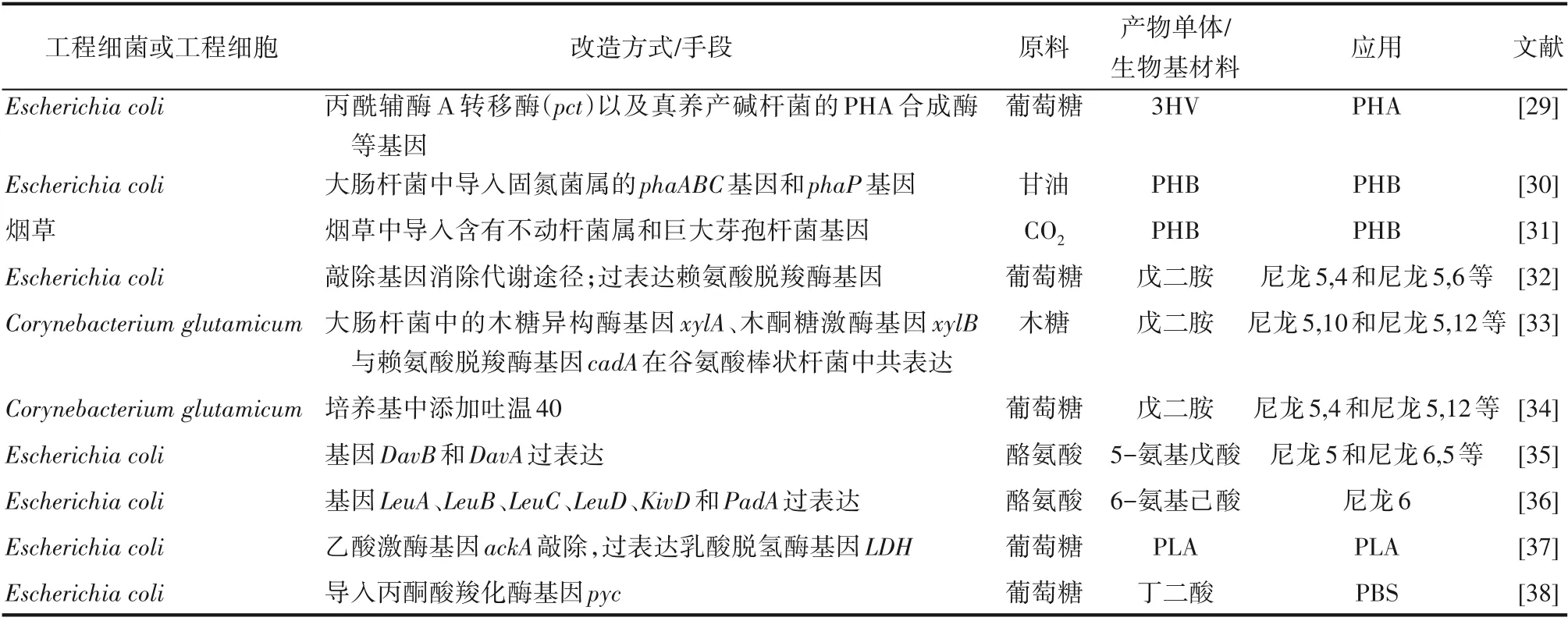
表1 工程细菌或工程细胞生产生物基塑料应用实例Table 1 Examples of applications of engineered bacteria or engineered cells to produce bio-based plastics
图1 合成生物学的应用领域Fig.1 Applied fields of synthetic biology
1.2.2 基因编辑技术 基因编辑技术最早是在同源序列间引入外源基因,实现基因编辑的同源重组技术(HR)。而后出现了人工核酸酶的基因编辑技术,主要有ZFN、TALEN、CRISPR/cas9 和单碱基编辑技术[50]。CRISPR/cas9 是适用性较强和打靶效率高的基因编辑技术,生物体基因组特定位点被精确地修饰,通过敲除、敲入和替换基因片段,生物体的特性或性状被改变,此方法能够实现高通量基因编辑,且设计简便、效率高、成本低,但也会出现脱靶问题。CRISPR/cas9 系统由内切酶cas92 和gRNA 两部分组成。工作原理为设计出预定目标序列的gRNA,利用3'端序列与cas9结合以及5'端序列与靶基因互补配对,定位PAM 前3~5 个碱基的距离对其中任一序列进行插入、敲除及突变等修饰(图2)[51−52],而 gRNA 可 以 通 过 在 线 网 站(OptimizedCRISPRDe−sign.com等)设计。
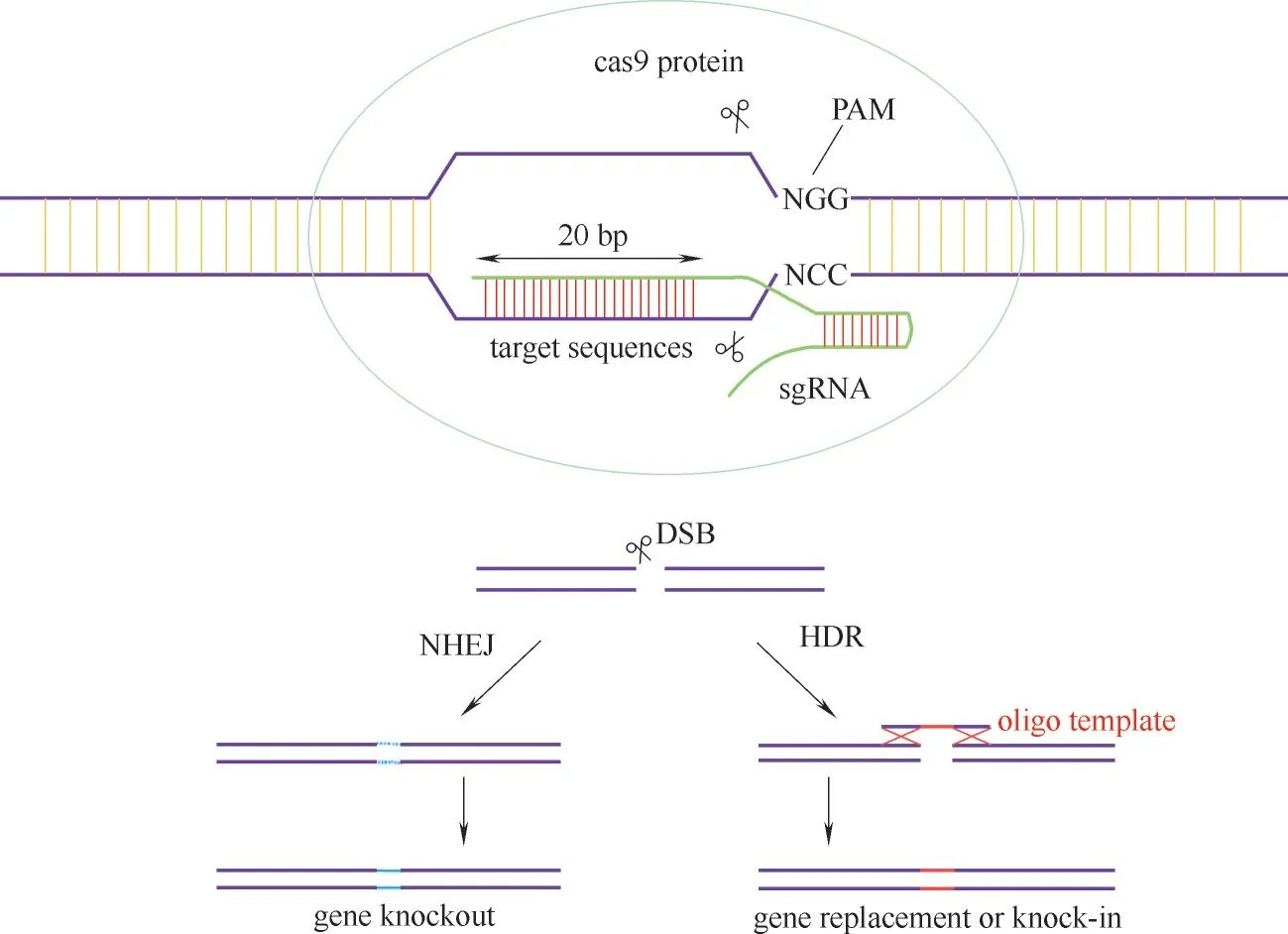
图2 CRISPR/cas9 基因编辑技术Fig.2 CRISPR/cas9 gene editing technology
1.2.3 基因芯片技术 基因芯片技术也是合成生物学的重要技术,未知序列的核酸序列与已知序列的核酸探针杂交检测,在固相支持物上原位合成(in situ synthesis)寡核苷酸或者直接将大量预先制备的DNA 探针以显微打印的方式有序地固化在支持物表面构成二维阵列,再将待测的标记样品基因按照碱基对配对原理进行杂交,从而检测待定基因。Ren等[53]研究了基因芯片对酿酒酵母中DNA 结合蛋白的功能和基因组的结合区域,通过酿酒酵母全部基因组信息的基因芯片技术,并结合经过修正的染色质免疫沉析,实验结果证明此技术能够有效发现酿酒酵母细胞体内转录激活子直接控制的基因。Zimmer 等[54]利用基因芯片技术比较了超水平表达的Ntrc 激活的突变菌株和野生型菌株的RNA水平,发现了大肠杆菌中存在调节子Ntrc,大肠杆菌基因组中约有2%的基因表达被调节子Ntrc 控制。基因芯片技术具有高集成、高通量、自动化和多样化等优点。虽然在技术与设备方面还存在一些不足,但是基因芯片技术已经展现出较大价格下降空间,在合成生物学领域代谢途径构建中基因表达分析、DNA 测序和发现新基因等方面具有巨大的应用价值。
此外,在工程酶构建、基因组信息分析、基因表达量调控等方面,合成生物学也发展了许多新技术,并且与目前的人工智能和机器学习等新技术相结合,促进了可控生物体系构建过程的自动化和标准化。
2 生物基塑料
塑料根据原料来源不同分为石油基塑料和生物基塑料。石油基塑料来源于不可再生的石油资源,生物基塑料原料为天然可再生生物资源。而随着世界各国对石油资源的争夺和日益显著的环境问题,生物基塑料相比石油基塑料,具有原料种类丰富、可再生和生产过程环保等优点。生物基塑料不等同于生物降解塑料,两者本质上是不同的,目前生物基塑料大部分是可降解塑料,有些生物基塑料是不可降解的。生物基塑料占据原料优势,采用合成生物学先进技术,可实现优化合成过程,提高合成效率,得到精确化、多功能化及可控的合成途径。因此,生物基塑料在推行低碳经济的当今世界具有广阔的市场应用前景。
2.1 市场发展状况
2017 年,欧洲生物塑料协会报告了生物基塑料行业市场(图3)及预测。根据欧洲生物塑料行业协会分析报告[55],2016 年全球塑料年产量约4 亿吨,其中生物基塑料占当年塑料年产量的1%,预测2016年至2021 年,全球生物基塑料将从416 万吨增长至610 万吨[56]。目前,随着人们生活水平的提高,环保意识的增加,国家管理部门的政策性倾向,合成生物学技术的快速发展,市场需求量将会大幅增长,将会涌现出更多种类丰富、结构复杂的生物聚合物。就我国生物基塑料应用市场与国外相比,欧洲市场增幅突显,国内市场目前尚方兴未艾。
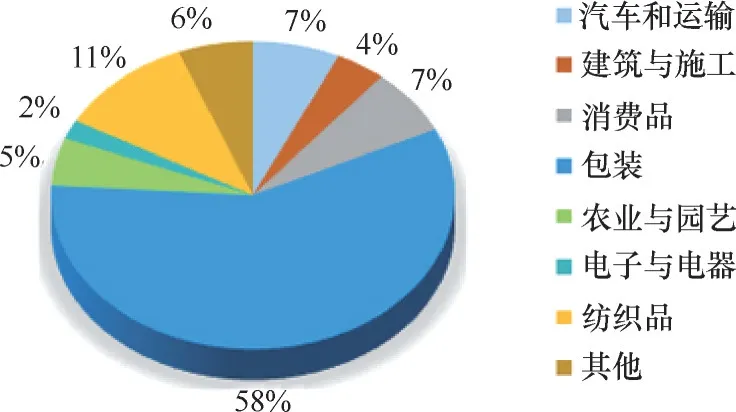
图3 2017年生物基塑料的应用市场分布Fig.3 Application market distribution of bio−based plastics in 2017
2.2 新型材料的发展
目前市场上生物基塑料商业化种类较多,占据较多的生物基塑料为可生物降解的PHA(聚羟基链烷酸酯)和PLA(聚乳酸)两种创新型生物聚合物,也是生物基塑料制品增长的主要产品。PHA 作为聚合物家族的重要组成,技术较为成熟,已经实现商业化。PLA 被称为工业万能材料,可以替代PS(聚苯乙烯)、PP(聚丙烯)和ABS(丙烯腈丁二烯苯乙烯)等传统材料。新型的生物质材料PHA 和PLA 与传统石油基塑料结构完全相同,除生产原料外,生产设备的初始至终端均完全相同。此外,当前国内外市场上,技术成熟的生物基单体主要有丁二酸、丁二醇、乳酸和丙二醇等。为更好地推动生物基塑料的发展,欧洲一些国家和组织通过立法推动生物基塑料的应用和市场发展。法国于2017年1月1日执行超市果蔬包装袋和商业邮件塑料包装制品必须为生物基材料,可降解袋和堆肥袋制品中生物基碳含量由2017年30%增长到2020年60%[56]。因此,塑料制品的市场发展方向将由石油基塑料逐渐转向生物基塑料。
3 合成生物学在生物基塑料中的应用
在过去的数百年里,合成有机化学主导着化学品的生产,合成的塑料制品为人类的生活带来了极大的便利。合成有机化学消耗的是愈发匮乏的石油资源,且生产过程造成环境污染。世界各国因石油资源的争夺战争不断,石油基塑料的价格不断上涨,企业的利润明显降低。生物基塑料采用取之不竭的绿色再生生物质资源(如玉米、甘蔗、秸秆和木屑等)合成,避免了化石资源的依赖。随着合成生物学的迅猛发展,采用微生物发酵制备生物基塑料得到各国政府、科研机构及产业界的极大重视。合成生物学已经发展成为制备生物基塑料的重要手段[57−59],此应用为人类工业制品的发展提供了新的方向,解除了对石油资源的依赖,解决了废弃生物质处理问题,缓解了日益严峻的环境问题,该方向也成为了当前环境、材料领域学术研究的重点热点之一。本文将综述合成生物学在聚羟基烷酸酯(PHA)、尼龙家族工程塑料、聚乳酸(PLA)和聚丁二酸丁二醇酯(PBS)几种典型生物基塑料的应用。
3.1 聚羟基烷酸酯(PHA)
PHA 是一种结构多样化、生物可降解的高分子聚合物,具有光学活性、生物相容性、生物降解性、热塑加工性和气体相隔性等优点,被广泛应用于塑料制品、人造医用支架材料、药物载体和电学材料等。PHA 材料单体为手性R 型的羟基脂肪酸,单体有多种侧链,单体链长多数为3 ~14 个碳原子。单体的组合方式、聚合反应方式和分子量等不同,使PHA 具有多样性,常见的PHA 单体为3−羟基脂肪酸。PHA 材料的多种单体结构采用合成生物学技术被成功设计和多种微生物合成,通过代谢途径从头构建和改造工程菌,可以拓展PHA 的合成种类,当前已经实现了PHA的可控合成和可设计性。
2010 年,聚羟基脂肪酸酯(PHA)合成技术被达沃斯世界经济论坛评为全球最具创新力技术。PHA在我国的研究已有20多年历史,清华大学陈国强课题组是我国PHA 的领先研究团队,依托此团队完成了5 t PHA的中试。Chen等[60]以葡萄糖为碳源,通过多种途径合成多种PHA,并对盐单胞菌(Halomonas)进行改造,实现了PHA 的无灭菌开放连续发酵低成本合成,推动PHA 的产业发展。此外,两种新的大片段基因组DNA 克隆技术被该团队开发。第一种是基于体外单链互补突出末端退火黏连的大片段基因簇调取方法(single−stranded overlapping annealing, SSOA)[61],此技术应用于克隆大肠杆菌基因组中的任意片段,效率都较高,克隆低于30 kb 的基因组片段,阳性率可达80%。第二种是体内同源重组的大片段DNA 克隆技术,可以免除依赖抗性基因,此方法已经应用于PHA 材料聚β−羟基丁酸酯(PHB)的生产中[62]。具体过程为通过大肠杆菌过表达乙酰辅酶A 和(或)氢酶−3 基因簇,在厌氧情况下,产氢与PHB 的合成相偶联,无论是否添加碳源乙酸,均能增加H2和PHB 的产量,H2含 量 提 高10 倍 以 上,PHB 产 量 提 高9 倍;PHB 产量由0.55 mg PHB/g 葡萄糖提高到5.34 mg PHB/g 葡萄糖。
目前,大部分常见的PHA 材料均可以葡萄糖为唯一碳源,运用合成生物学和代谢工程技术合成。具体过程为通过调控基因的表达量和细胞的代谢网格优化途径,即通过调节核糖体结合位点RBS 翻译强度或基因启动子转录强度达到PHA 代谢流的控制,进一步提高PHA 的产量。Poblete−Castro 等[63]通过敲除葡萄糖脱氢酶基因gcd,获得恶臭假单胞菌重组菌株,PHA 的合成能力比野生型恶臭假单胞菌提高了100%。CRISPR 及其衍生技术CRISPRi 能够通过基因敲除、替换等抑制目标基因的表达量达到调控代谢途径的目的。Lv等[64]采用CRISPRi技术抑制琥珀酸半醛脱氢酶sad 基因,获得不同抑制效果的大肠杆菌重组菌株,以葡萄糖为碳源,有效控制了合成4HB 的代谢流,合成了不同4HB 含量(4HB 的摩尔分数1% ~9%)的P3HB4HB。进一步抑制琥珀酸脱氢酶基因(sucC、sucD、sdhA 和sdhB)和琥珀酰辅酶A合成酶基因,P3HB4HB中4HB的含量调控范围扩大为1.4%~18.4%。
另一种常见的PHA 材料为3−羟基丁酸与3−羟基戊酸共聚酯[P(3HB−co−3HV)],简称PHBV,具有高阻隔性、完全的生物相容性、耐冲击能力强、较好的弹性和韧性等优点。由于PHBV 的合成前体是丙酰辅酶A,因此,更多的研究目标集中在丙酰辅酶A。如Yang 等[65]以葡萄糖为碳源,在大肠杆菌中成功构建了两条丙酰辅酶A 的代谢途径来合成PHBV。
3.2 尼龙家族工程塑料
尼龙即聚酰胺,简称PA。常见的石油基尼龙种类为尼龙6 和尼龙66。生物基尼龙采用生物法制取,种类有尼龙66、尼龙46、尼龙6、尼龙5、尼龙56、尼龙11、尼龙1010、尼龙610和尼龙612等。生物基尼龙制备方法有生物发酵法和植物油裂解法。目前大多数企业选用植物汕裂解法作为生物基尼龙的主要生产路线。但是随着合成生物学技术的快速发展,采用生物发酵法可大大降低尼龙单体的生产成本,具有较大的工业应用潜力和发展空间。接下来介绍几种合成生物学在尼龙家族工程塑料中的应用实例。
3.2.1 尼龙66 和尼龙46 尼龙66(PA66),学名聚己二酰己二胺,简称聚酰胺66,是一种重要的热塑性树脂,具有机械强度大、硬度高,刚性强等优点,作为一种新型工程塑料,广泛应用于机械设备、合成纤维、汽车配件和精密仪器等领域。工业上尼龙66的合成方法为己二酸和己二胺制成尼龙−66盐后缩聚而得。尼龙46(PA46),学名聚己二酰丁二胺,简称聚酰胺46,与工程塑料与耐热塑料相比,具有使用期长、耐疲劳性佳、耐摩擦和耐磨耗性均较好等优点,适用于电子电器材料。己二酸是生产尼龙66和尼龙46的重要原料,是一种具有重要商业用途的脂肪族直链二羧酸。
己二酸不存在天然合成途径,但可以通过工程酵母菌株产生的α 或ω 型氧化的长链正烷烃或脂肪酸等合成[66−68]。文献报道热带念珠菌通过ω 型氧化途径生产6−羟基己酸(6−hydroxycaproicacid)[69],此反应由NADPH 细胞色素P450 还原酶(CYP)和NADPH 细胞色素P450 还原酶(CRP)组成的细胞色素P450 羟化酶复合物引起,通过脂肪醇脱氢酶(ADH)或脂肪醇氧化酶(FAO)将6−羟基己酸转化为6−氧代己酸(6−oxohexanoicacid),6−氧代己酸经脂肪醇脱氢酶(AldDH)转化为己二酸。Picataggio等[70]过表达热带念珠菌HexS、CPR、CYP450、ADH、FAO和AldDH基因,并对其他基因进行了修饰,使更多的碳流被导入到构建的己二酸代谢途径中。经过6 d 的培养,C. tropicalsAA580 以椰子油为原料可产生4 g/L 以上的己二酸。Picataggio 等[71]进一步发展了发酵技术,己二酸的浓度在120 h 内能够提高到50 g/L以上,也是文献报道的最高滴度。
己二酸也可由粘糠酸经过化学催化加氢制得。Lin 等[72]以邻氨基苯甲酸为前体,加入两个外源酶,最后成功制得粘糠酸和儿茶酚;然后以分支酸经异分支酸合成酶(ICS)和异分支酸丙酮酸裂解酶(IPL)催化合成水杨酸,水杨酸单加氧酶(SMO)和儿茶酚双加氧酶(CDO)催化也可以生成粘糠酸。经过条件优化,粘糠酸产量能够达到1.5 g/L,开辟了由粘康酸制备己二酸的新途径。
尼龙66 的另一单体己二胺的生物合成未见文献报道。
3.2.2 尼龙6 尼龙6,学名6−氨基己酸(6−aminocaproicacid,6ACA),是一种重要的工程塑料,广泛应用于机械、化工、仪表、汽车、医疗设备及针织品等领域。6ACA 也是一种重要的非蛋白质直链氨基酸,也是临床常用的止血药,能抑制纤维蛋白的溶解而止血[73]。
6ACA 可作为己内酰胺的前体,己内酰胺单体聚合可合成出尼龙6。生物方法合成6ACA 的文献报道较少,且均采用微生物体内代谢途径构建法,前体来自于TCA循环。Turk等[74]构建了两条代谢途径生产6ACA,一条代谢途径为过表达3−oxoadipyl−CoAthiolase, 3−hydroxyadipyl−CoAdehydrogenase,enoyl−CoAhydratase,hexenoyl−CoA−reductase,kdcA和vfl 基因,由乙酰CoA 和丁二酰CoA 缩合经己二酰CoA 制备6ACA,另一条途径为过表达nifV,aksD,aksE,aksF,kdcA 和vfl 基因,由TCA 循环的中间产物α−酮戊二酸(AKG)经碳链延伸得到酮庚二酸,最后经脱羧和转氨作用,得到6ACA,其浓度为160 mg/L。Chae 等[75]通过nifV,aksF,aksD,aksE,vfl 和kdcA 共表达,获得6ACA 作为生产己内酰胺的中间产物。Zhou 等[76]在pMB1 质粒上构建了两个基于PT7 启动子的操纵子,表达nifV,aksF,aksD 和aksE,在pBBR1质粒上构建了基于PT7 启动子的操纵子,实现vfl 和kdcA共表达,6ACA的最终产量为48 mg/L。
近期,本课题组构建了多酶分子机器RaiP+LeuABCD+KivD+PadA,见图4。以大宗化学品赖氨酸为原料,通过赖氨酸脱氨基产物2−酮酸和乙酰辅酶A 聚合引发链延伸循环,实现了+1C 获得产物6ACA[36]。但此循环反应体系中,碳链不断增长,实际产物中既出现了+2C 反应产物7AHA,同时仍有大量未能成功+1C 的副产物5AVA 生成。最终产物为5AVA 99.2 mg/L,6ACA 47.0 mg/L,7AHA 4.78 mg/L,即总的非天然直链氨基酸(NNSCAAs)的产量为140.98 mg/L。下一步拟通过定向进化和溶剂强化等技术,进一步提高6ACA 的合成效率,并解析碳链延伸循环圈的作用机制。
3.2.3 尼龙5和尼龙56 尼龙5和尼龙56是两种重要的新型工程塑料。 5−氨基戊酸(5−aminovalericacid,5AVA)是尼龙5 和尼龙56 的前体,也是合成5−羟基戊二酸、戊二酸和1,5−戊二醇的C5平台化合物。化学法和生物法合成均可合成5AVA,化学法合成过程复杂,产率较低,且造成环境污染。5AVA 的天然合成途径存在于恶臭假单胞菌(Pseudomonas putida)中,5AVA 的人工合成途径也已经成功被构建。Park 等[77]通过重组大肠杆菌WL3110 表达假单胞菌putidadavAB(基因编码delta−aminovaleramidase) 和 赖 氨 酸 2−monooxygenase,成功制备出5−氨基戊酸盐(5AVA)和戊二酸。Ma 等[78]设计了L−赖氨酸转化为5AVA 的生物途径,重组大肠杆菌过表达信号肽PelB 和DAP转运蛋白CadB,可生产221 g/L 5AVA,5AVA 产量提高了12%。
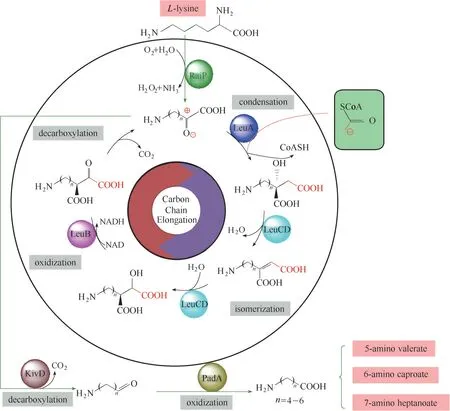
图4 产6ACA的回归循环多酶分子机器[36]Fig.4 Regression cycle multienzyme molecular machine of 6ACA[36]
本课题组通过RaiP 的过表达,构建了生产5AVA 异源途径系统[79],见图5。首先,构建了质粒pCJ01,将其转入大肠杆菌BL21(DE),获得了在T7启动子作用下RaiP 过表达的工程菌株CJ01。赖氨酸在大肠杆菌中被赖氨酸脱羧酶CadA 天然降解为戊二胺。为了减少底物的降解和提高底物的利用率,将CadA进行敲除,获得了敲除CadA减少赖氨酸消耗的工程菌株ML03。L−赖氨酸可以被RaiP 氧化脱氨,得到2−酮−6−氨基己酸,随后在氧化剂H2O2的作用下相继脱羧和氧化为5AVA。菌株CJ00、CJ01和CJ02 在添加L−赖氨酸后产生了5−氨基戊酸。再利用4% 乙醇和10 mmol/L H2O2处理全细胞生物催化剂CJ02RaiP,催化L−赖氨酸HCl 反应48 h 后,5−氨基戊酸的产量达到了50.62 g/L。
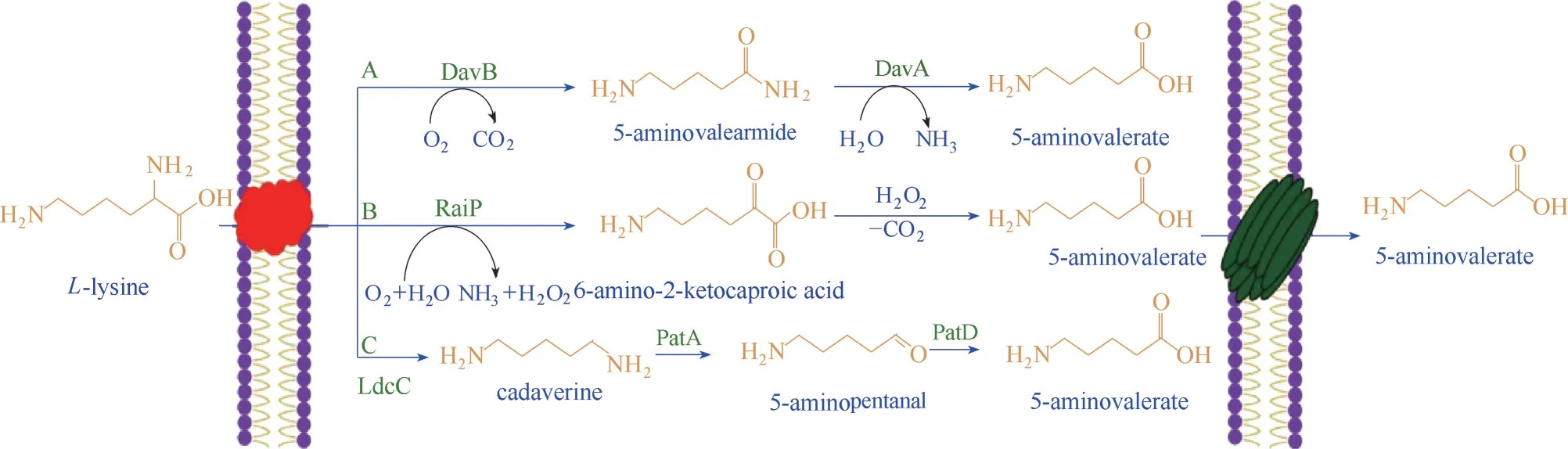
图5 利用L−赖氨酸在微生物中合成5−氨基戊酸的各种途径[79]Fig.5 Various routes of 5AVA biosynthesis from L−lysine in microorganisms[79]
3.3 聚乳酸(PLA)
聚乳酸(PLA)是一种新型的生物基材料,具有良好的生物降解性、生物相容性、热稳定性、抗溶剂性和易加工性等优点,广泛应用于服装制造、建筑和医疗卫生等领域,是合成生物学在材料领域的最早应用之一。
PLA 的合成单体是乳酸(2−羟基丙酸),是一种具有较大市场空间和发展潜力的化工原料,具有很高的商业价值。PLA 在发酵工业中用于控制pH,在食品行业中充当调味剂。时梦询[37]采用原始菌株大肠杆菌BL21(DE3),运用染色体重组技术,构建敲除乙酸激酶基因ackA 的自杀质粒pRE112(图6),抑制了葡萄糖到乙酸的代谢流,并且过表达乳酸脱氢酶基因LDH,增强了葡萄糖到乳酸的代谢流,高产乳酸菌E.coli BL21(DE3)△ackA/LDH−pTrcHis2B 被成功构建,实验结果为构建的工程菌生产乳酸产量比野生菌高48.20%。Grabar 等[80]同时将pflB、富马酸还原酶基因(frd)、adhE 和ackA 基因删除,构建的工程菌株在无机盐培养基中发酵积累的D−乳酸(光学纯度95%)达110 g/L,将甲基丙酮醛合成酶基因(mgsA)进一步删除,乳酸产量能够增加到118 g/L,且达到99.9%的光学纯度。周丽[81]通过单基因删除和多基因组合删除手段研究乳酸在大肠杆菌的代谢,为减少产物中的副产物,采用选择性地组合方式删除了野生型大肠杆菌中的8 个基因:ackA、pta、磷酸烯醇式丙酮酸合酶基因(pps)、pflB、FAD依赖型D−乳酸脱氢酶基因(dld)、丙酮酸氧化酶基因(poxb)、adhE 和富马酸还原酶基因(frdA),有效控制了多种副产物的含量,使产物D−乳酸产量达到125 g/L,光学纯度大于99.9%。
3.4 聚丁二酸丁二醇酯(PBS)
聚丁二酸丁二醇酯(PBS)是一种典型的可完全生物降解的聚合物材料,具有良好的生物相容性和生物可吸收性,易被自然界中多种微生物或动植物体内的酶分解,代谢终产物为CO2和H2O。
PBS是由丁二酸和丁二醇单体聚合得到。其中单体丁二酸又称琥珀酸,是一种基础平台化合物,具有较高的经济价值。目前丁二酸的产量无法满足PBS产业化需求,通过生物质生产丁二酸,丁二酸转化丁二醇,合成的环保廉价生物基PBS 具有广阔的市场应用前景。自然界中存在丁二酸的天然合成途径,在厌氧条件下,野生型大肠杆菌混合酸发酵可合成少量丁二酸,将大肠杆菌基因工程改造后,可将有机酸产物的竞争途径消除,丁二酸的合成代谢流增强,大大提高大肠杆菌合成丁二酸的能力。Vemuri 等[38]重组大肠杆菌(失活丙酮酸甲酸裂解酶基因pfl 和乳酸脱氢酶基因ldh)筛选出葡萄糖转移酶基因ptsG 产生自发突变菌株AFP111,甲酸和乳酸在该突变株中将不再合成,再将AFP111 菌株导入丙酮酸羧化酶基因pyc,丁二酸的代谢途径增强,经过好氧-厌氧二阶段发酵培养,丁二酸产量高达99.2 g/L,生产速率达1.3 g/(L·h)。此外,分子改造或进化代谢选育手段能够恢复或增强大肠杆菌菌株在厌氧条件下对五碳糖的利用能力[82−83]。
与国外相比,我国丁二酸发酵商业化还存在一定差距,但是中国科学院过程工程研究所、华东理工大学、南京工业大学和重庆大学等研究人员对丁二酸发酵研究已经取得了丰硕的成果。
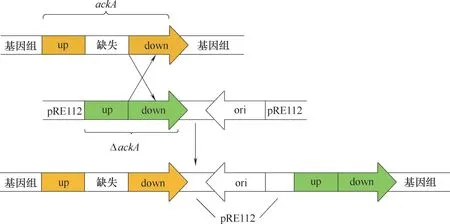
图6 ackA基因敲除原理[37]Fig.6 Gene ackA knockout principle[37]
中国科学院过程工程研究所邢建民课题组[84]基于Red重组系统,采用“一步法”敲除了大肠杆菌菌株基因组上的染色体片段,将大肠杆菌菌株W3110的ldhA 和pflB 基因敲除,成功构建出双基因敲除菌株BS002[85−86]。根 据GenBank 公 布 的A.succinogenes 130ZPCK 序列,分别设计了引物pck−F 和pck−R,引物带有BglII 和KpnI 酶切位点,通过PCR 扩增获得目的片段,进行双酶切处理PCK 基因片段和pTrcHisA 质粒,连接后将表达质粒pTrcHisA−130ZPCK 转入BS002,成功构建了重组菌株BS101。工程菌BS101发酵8.14 g/L葡萄糖可积累4.77 g/L丁二酸和2.65 g/L 丙酮酸,丁二酸产率达到0.90 mol/mol,比野生型大肠杆菌提高3 倍,此过程几乎不产生甲酸、乙酸和乳酸。
华东理工大学吴辉课题组[87]采用醋酸盐生产丁二酸。通过丁二酸脱氢酶(sdhABCD 编码)的失活阻断了TCA 循环,使葡萄糖在有氧条件下积累丁二酸。由于乙醛酸旁路成为丁二酸生物合成的主要途径,其中aceBAK 操纵子的IcR 抑制子抑制乙醛酸旁路,IeR 的缺失促进了葡萄糖产生丁二酸。因此,菌株MG02 在MG01 中敲除iclR。与MG01 相比,MG02 在72 h 内多消耗了一点乙酸盐(12.4 mmol/L),产生了更多的丁二酸盐(2.81 mmol/L)。
本课题组[88]研究了厌氧诱导的PSnirB 启动子替代外源诱导剂,工程菌株生产丁二酸的情况。以葡萄糖、混合糖或者红薯渣水解液为直接碳源,与IPTG 诱导的Trc 启动子调控下的工程菌SD133 和SD134 相比,在厌氧诱导的PSnirB 启动子调控下的工程菌HD133 和HD134 可以获得更高浓度的丁二酸。以红薯渣水解液为直接碳源摇瓶发酵48 h 后,工程菌HD133 和HD134 可分别产生16.73 g/L 和18.65 g/L 的丁二酸,与SD133 和SD134 合成的16.01 g/L 和17.18 g/L 丁二酸相比,产量分别提高了4.50%和8.56%,且无须添加昂贵的外源诱导剂IPTG。
4 展 望
随着合成生物学相关技术迅速发展到日益成熟,合成生物学在生物基塑料领域的应用也越来越广泛。生物基塑料原料丰富且可再生,过程绿色智能可控,条件转换温和,产品的种类也越来越丰富。合成生物学除了合成出可降解的生物基塑料PHA、尼龙家族、PLA等,也合成了一些非生物降解的生物基塑料PE、PET 和多元醇聚氨酯等。当前生物基塑料的发展也面临一些挑战,原料多采用粮食作物,采用农业废弃物或者生活废弃物等原料将是未来发展方向;生产成本还存在普遍偏高,且有些生物基塑料的综合性能难以全面替代石油基塑料,有些需要进一步改性才能满足需求。因此,开发低成本、高性能生物基塑料将是生物基塑料未来发展的方向。生物基塑料价格控制可从多方面降低成本,如选择价格低廉的糖类化合物或者废弃物为底物;利用代谢工程对微生物改造,开发新的代谢途径,通过从细胞的整体代谢网络着手,调控和优化代谢途径,提高底物到产物的转化率;选用特殊底盘菌如嗜盐菌等可抑制其他细菌的生长,可节省灭菌消耗的高温高压蒸汽的能量费用和高温高压设备需求的成本,使发酵生产过程中的耗能降低。总之,生物基塑料的发展前景具有极大潜力和生机活力,市场潜力也是无可估量的。
