基于EMS诱变的晋麦90号小麦抗寒突变体创制
2020-10-16武银玉范绍强孙黛珍曹亚萍
武银玉,范绍强,孙黛珍,曹亚萍
(1.山西农业大学小麦研究所,山西临汾041000;2.山西农业大学农学院,山西太谷030801)
小麦是我国重要的粮食作物,小麦生产可持续发展对于保障国家粮食安全和维护社会稳定都具有重要意义。近年来,在全球气候变暖的背景下,冬小麦种植区域呈现不同程度的北移西扩,部分栽培品种趋于春性化[1],育成品种也降低了对小麦抗寒性的考验[2],加之冬季持续高温,致使小麦抗寒锻炼不足,生育期提前,遇冬春冻融交替和春季低温(倒春寒)等异常天气,将对小麦生产构成潜在威胁,极端气候成为小麦产量损失的主要灾害[3]。
甲基磺酸乙酯(Ethyl Methane Sulfonate,EMS)是一种高效稳定的烷化类诱变剂,能诱发产生高密度系列等位基因点突变,具有种质创新频率高、遗传变异谱宽、育种周期短等特性,可弥补小麦常规育种短时间难以获得新性状和新基因的不足,在创造作物新品种、新种质、遗传材料以及解决育种工作中某些特殊问题等方面取得突破,育种中多应用于对作物的某一特殊性状进行改良。目前,小麦EMS诱变技术已在国内外得到大规模研究和应用。HUSSAIN等[4]用EMS诱导中抗叶锈小麦品种NNGandum-1,得到9个高抗和2个高感突变体;李晓等[5]用EMS诱变京411,获得Wx-A1基因的7个点突变,其中,有功能变异的4个突变系均可稳定遗传至下一代;ZHANG等[6]用EMS处理YZ4110,获得晚抽穗期突变体m605,该突变体可使秋季播种时间至少延迟7 d。迄今尚未发现有采用EMS诱变技术诱导抗寒突变体的报道。
晋麦90号是山西农业大学(山西省农业科学院)小麦研究所采用远缘种间渐进式复合杂交和穿梭育种相结合的方法选育而成的小麦品种,2011年通过山西省品种审定。该品种抗旱节水,根系发达,抗干热风能力强,抗倒伏性好,成熟落黄好,生物产量和经济产量都较高,是一个优良的旱地小麦品种,品质符合国标中筋优质麦标准,粗蛋白质15.21%,湿面筋35.0%,稳定时间6.2 min。但是该品种尚存在缺陷,即抗冻耐寒性较差,常年抗寒性为2级,遇寒冷灾害年份易对产量造成较大影响。
本研究以晋麦90号小麦品种为试验材料,用EMS处理种子,在鉴选表型突变体的基础上,对抗寒性进行了鉴定和分析,旨在选育抗寒突变体,为小麦抗寒育种和基因研究提供新的种质资源。
1 材料和方法
1.1 试验材料
1.1.1 种子诱变处理 晋麦90号由山西农业大学小麦研究所保存。参照曹亚萍等[7]提供的方法,挑选晋麦90号大小均匀、籽粒饱满的种子1 kg,用蒸馏水室温浸种10 h,使种子充分膨胀萌动后,放在吸水纸上晾干,18℃条件下用0.8%EMS诱变剂浸种10 h,装入网袋,用自来水反复冲洗0.5 h,去除表面残留的EMS诱变液,风干,2 d后播种。
1.1.2 诱变材料种植 M1不进行选择,按单株收获,脱粒后按收获籽粒由多到少顺序编号,作为突变体的原始编号,共计1 282份,编号为1~1 282;M2单行等粒点播,根据株高、株型、穗型、叶色、分蘖等选择各种表型突变体,将突变体按株系分单株收获;对M3突变体进行抗寒性鉴定。
1.2 试验设计
M2分2个试点种植,试点1位于山西省临汾市尧都区山西农业大学小麦研究所试验田(N36°6′24″,E111°30′48″),属黄淮中部麦区;试点 2 位于山西省晋中市太谷县山西农业大学试验田(N37°25′16″,E112°34′52″),属黄淮北部麦区。2 个试点种植同一套材料,按原始编号顺序排列,单行区,行长2 m,行距0.25 m,每行30粒,每30行插入1行对照野生型晋麦90号。试点1用于进行表型突变体鉴选,试点2参照试点1突变体鉴选结果,按编号收获能正常越冬成熟的突变株系中的每个单株,各单株编号是在原始编号后加二级编号,如原始编号为302的株系共收获7个单株,则每个单株编号分别为302-1、302-2、……、302-7。M3将试点 2 收获籽粒较多的单株种植于试点1,采用二级编号,顺序排列,3次重复,每单株种植单行区,行长2 m,行距0.25 m,每行40粒,每30行插入1行对照野生型晋麦90号。试验田肥力均匀,播种深度一致,四周设有保护行。
1.3 抗寒性鉴定
依据返青期叶片和茎受冻害程度进行鉴定,按照赵瑞玲等[8]描述的鉴定方法,将冻害分为5级,即0级:叶片没有冻死部分;1级:叶片冻死部分仅限于叶尖,下部叶片未受冻或很少,全田绿色叶较多;2级:叶片冻死部分少于绿叶部分,但下部叶冻死者较多,地面有枯黄叶;3级:叶片冻死部分多于绿叶部分,地面可见整片枯叶,绿色叶片较少;4级:上下部叶片全部冻死枯黄,全是枯叶,甚至全株死亡。记载时,每小区冻害等级依据80%单株表现而定,并增设“-”、“+”符号作补充,如“2-”表示比 2 级冻害轻些,“2+”表示比2级冻害重些。
1.4 统计分析
为方便计算,冻害等级 0、1-、1、1+、2-、2、2+、3-、3、3+ 分别赋予抗寒值为 10、9、8、7、6、5、4、3、2、1。采用Microsoft Excel 2003整理数据并作图,采用SPSS24.0统计软件进行方差分析。
越冬成活率=收获株数/幼苗株数×100%(1)
2 结果与分析
2.1 突变体抗寒性筛选
在试点1对M21 282份诱变材料进行农艺性状鉴定,全生育期每3~5 d田间调查一次,观察记载幼苗、叶片、茎、分蘖、穗部、育性等性状,共筛选到126份表型突变体。试点2因平均气温较低、气候条件严酷,与试点1对应的126个突变体中有114个有正常越冬植株,共收获692株,越冬成活率仅为18.3%,明显低于野生型品种晋麦90号的平均越冬成活率(52.3%),说明突变群体中抗寒性弱的突变体多于强突变体,EMS诱变对晋麦90号抗寒性具有负向效应。
2.2 突变体抗寒性鉴定
选择M2收获籽粒较多的突变体共计265份,在试点1对M3进行抗寒性鉴定,结果表明,晋麦90 号突变体冻害等级分布在 0、1-、1、1+、2-、2、2+、3-、3、3+,共 10 个等级,由于 M2在寒冷生态区进行了抗寒性筛选,无4级冻害突变体出现,其中,晋麦90号属“2-”级。在所有突变体中,冻害级别分布主要集中在2-、2、2+、3-这4个等级,分别占调查株系总数的16.23%、19.62%、19.62%和16.98%;其次为 1、1+、3、3+ 等级,分别占调查株系总数的3.78%、8.68%、7.93%、5.28%;抗寒性强的突变体较少,0级和1-级分别占总数的0.75%和1.13%。
从M3突变体抗寒性分布来看,3个重复抗寒性趋势一致(图1)。在鉴定突变体抗寒性时,对冻害等级进行了更为详细的划分,每一等级鉴定出数量不等的突变体,表明晋麦90号突变体抗寒性分布由强到弱存在连续性,自交纯合后将得到抗寒性由强到弱的系列表型变异,有利于进行小麦抗寒基因结构与功能研究,为推动抗寒性基础研究储备遗传材料。
2.3 突变体抗寒性差异分析
对M3所有株系抗寒性按由强到弱排列并进行新复极差比较,在1%差异水平下,较晋麦90号抗寒性极显著增强的有38个株系、极显著减弱的有176个株系,无极显著差异的有51个株系;在5%差异水平下,较晋麦90号抗寒性显著增强的有38个株系、显著减弱的有183个株系,无显著差异的有44个株系。数据比较分析结果表明,265个突变体抗寒性由强到弱呈连续性分布,分别发生在114个诱变的原始单株上,推测可能存在不同的突变位点,可得到抗寒性由强到弱的系列表型变异和突变基因;以野生型品种晋麦90号为对照,抗寒性弱的突变体多于强突变体,加之M2冷胁迫选择剔除了部分弱抗寒性突变体,再次证明EMS诱变对小麦抗寒性突变具有负向效应。
将5个抗寒性最强的突变体分析结果按顺序列于表1,可以看出,这5个材料抗寒值最低为9,冻害等级为1-,田间调查发现,仅下部部分叶片叶尖受冻,表现为极强的抗寒性;这5个突变体抗寒性也具有极显著差异,112-1抗寒性极显著强于927-1、718-5、316-4这 3 个材料,542-3抗寒性极显著强于316-4、718-5这2个材料。鉴定出的5个突变体分别来源于不同的原始单株,如突变位点存在差异,则将得到5个强抗寒性基因,成为小麦抗寒育种的优异基因资源。
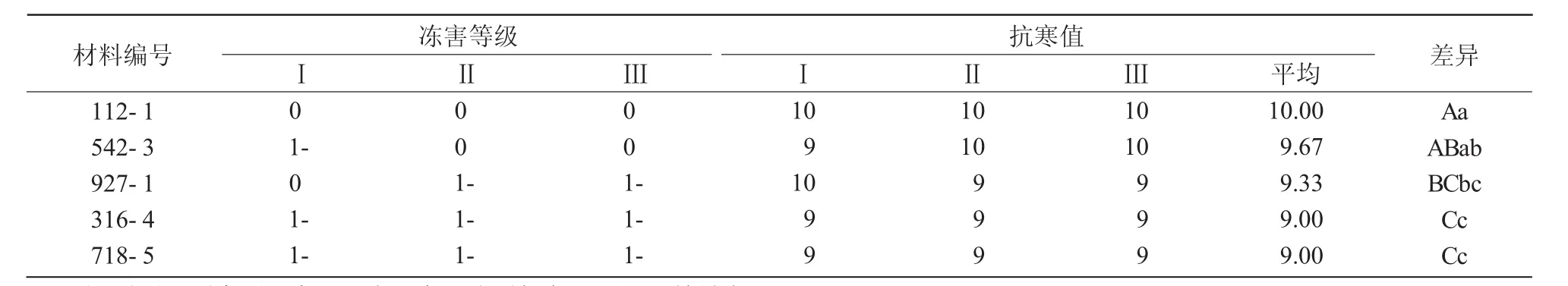
表1 强抗寒性突变体差异性比较
对原始编号相同且二级编号种植数目≥5的8个M3群体进行方差分析,结果表明(表2),不同群体的抗寒性在3个重复间均表现一致,没有显著差异,说明试验田生态条件同质性较好,生育期间试验田未出现局部寒流影响,同时也说明M3各株系内抗寒性趋于稳定;而株系间方差除186群体外,其余均存在极显著差异,说明突变体抗寒性在M3株系间仍表现较大分离。
对株系间存在极显著差异的7个群体分别进行新复极差比较,结果表明(表3),各群体内株系间抗寒性差异表现为2种类型:3个群体株系间抗寒性呈间断性分布,如295群体差异明显分为3个等级,295-10 抗寒性最好,295-1 次之,295-7、295-8、295-9株系较差,404群体、506群体内株系间抗寒性差异同样可明显分为2个等级;4个群体株系间抗寒性呈连续性分布,如原始编号为326的群体,5个株系抗寒性不能进行明确分类,326-9与326-3、326-5、326-1 存 在 极 显 著 差 异 ,326-3、326-5、326-1株系间无显著或极显著差异,但326-6仅与326-1存在极显著差异,而与326-3、326-5间无极显著差异,302、718、812群体也存在这种情况。另外,各群体内株系间抗寒值极差也存在较大差异,如718群体,株系间抗寒值极差为6,718-5株系抗寒性最强,冻害等级为1-,718-10、718-11抗寒性最弱,冻害等级均为3-,说明该群体内株系间抗寒性分离较大;而326群体抗寒值极差仅为1.33,说明该群体整体抗寒性趋于一致。
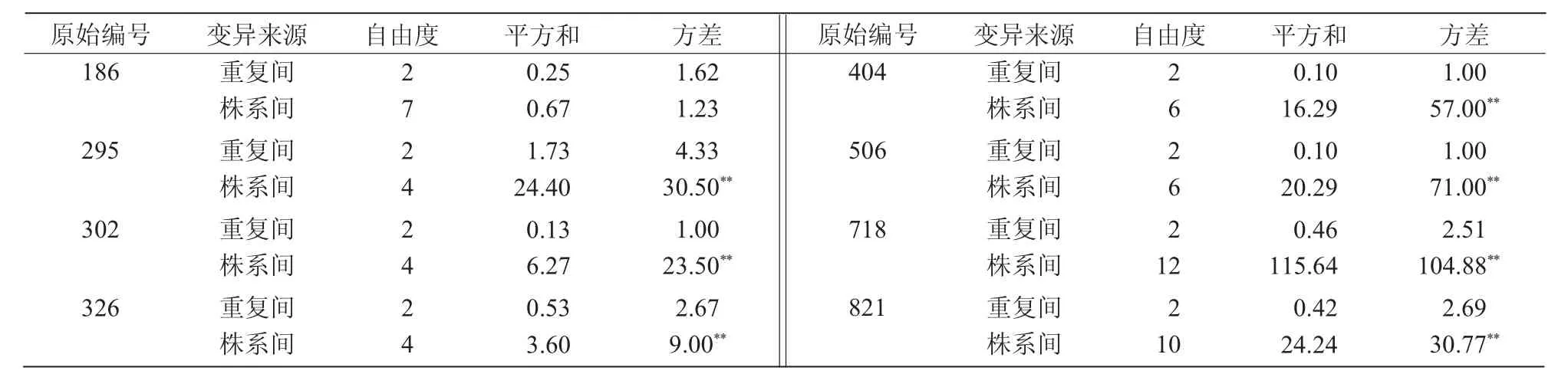
表2 突变体抗寒性方差分析
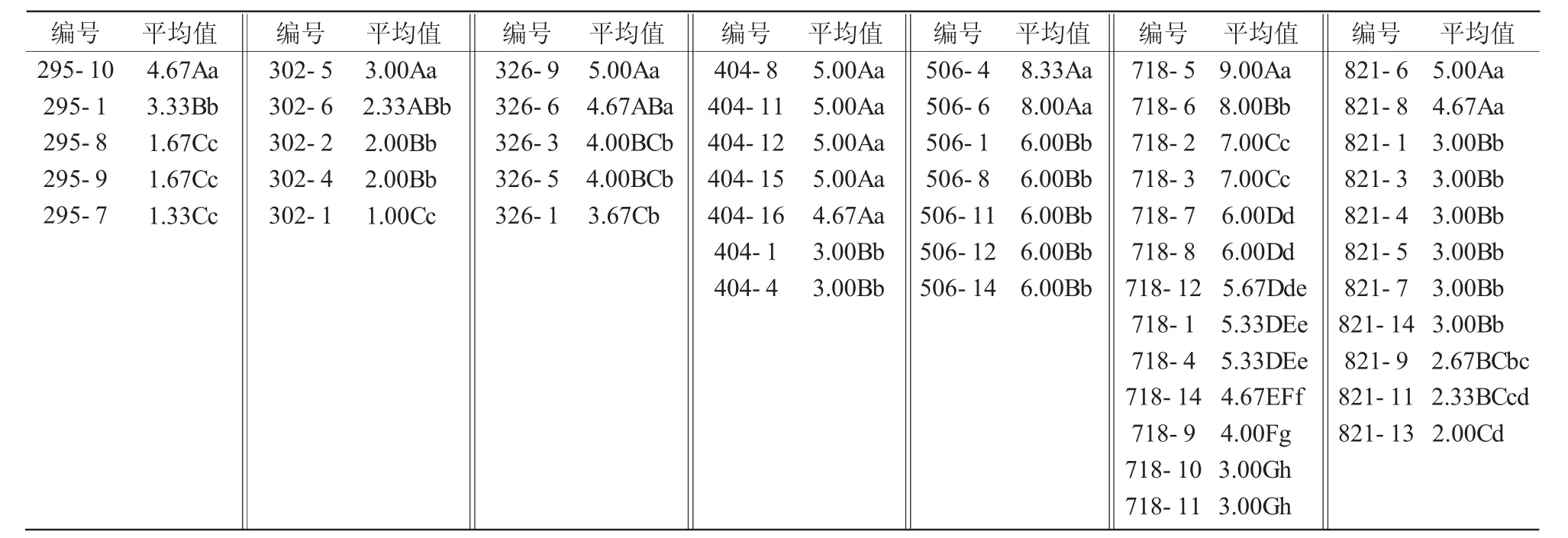
表3 突变体株系抗寒值间新复极差比较
3 结论与讨论
低温冻害是黄淮麦区频发的自然灾害之一,严重影响并制约着小麦的生长发育和产量形成[9-10]。提高小麦抗寒性的途径可归纳为3个:一是外源诱导。WANG等[11]将水杨酸(SA)溶液喷洒在扬麦16小麦叶片上,诱导内源H2O2和ABA积累,可显著提高品种的抗冻性;刘丽杰等[12]通过叶面喷施E-BR可促进小麦幼苗对可溶性蛋白、可溶性糖和脯氨酸的积累,从而增强小麦对低温的抗性。二是抗寒基因。目前小麦21条染色体已发现有15条携带抗寒相关基因[13],5A和5D染色体携带有抗寒主效基因[14-15],研究较多的基因有Vrn、Fr等;Fr2(Frost resistancs 2)为抗霜冻基因,包括几个串联重复的CBF(C-repeat bindingfactor),诱导COR基因(cold-regulated gene)的表达,可显著增强小麦对干旱和霜冻胁迫抗性[16-17];游光霞等[18]研究表明,Vrn1基因是小麦抗寒性遗传调控的关键位点,其会显著降低品种的抗寒性,具有2个或3个Vrn1基因的品种一般抗寒性较弱,3个位点均为隐性基因的小麦品种具有较强抗寒性,建议黄淮海北部麦区加强选育、推广vrn-A1、vrn-B1、vrn-D1基因型品种。三是抗寒育种。MICHEL等[19]研究表明,用抗寒性好的材料进行杂交,并用抗冻性QTLFr-A2进行标记辅助选择,可培育耐寒性小麦品种;GRABOVETS等[20]分析了俄罗斯国家38个品种,发现有29个具备抗寒性并可承受-18℃低温,表明选用遗传差异较大的材料进行杂交,在霜冻频繁地区采用系谱法选择,可以获得抗寒性强的小麦品种。迄今尚未发现运用诱变方法进行小麦抗寒种质创新进而提高小麦品种抗寒性的研究。本研究用EMS诱变剂处理晋麦90号小麦种子,M2选择各种表型突变体,并在寒冷生态区进行抗寒性筛选;M3进行抗寒性鉴定,得到5个强抗寒性突变体,创新了小麦抗寒种质创制方法。
本研究在抗寒新种质鉴选时,M2选择了2个试验点,试点2位于晋中市太谷县,该区平均气温较低,气候条件严酷,秋冬季冷空气活动频繁,常年种植强冬性高抗寒品种,在该试点对M2进行冷胁迫强化选择,可以剔除大批低温敏感材料,减轻后代选择压力;M3在试点1临汾市尧都区进行抗寒性鉴定,恰逢暖冬干旱年份,2019年12月平均气温为0.07℃,较2018年(2018年12月平均气温-1.85℃)高1.92℃,导致小麦冬季持续生长,抗寒锻炼时期缩短,抗寒能力降低,2020年1月5—7日出现21.67 mm降水(雪),1月11日积雪消融,冻融交替后又遇到1月14日春寒,同期播种小麦普遍受冻,为突变体的抗寒性鉴定提供了良好的生态条件。
本研究通过EMS诱变和抗寒性筛选,创制出5个抗寒性好的突变体,即112-1、316-4、542-3、718-5、927-1,分别来源于不同的原始单株,方差分析结果表明,其抗寒性具有差异,推测这5个突变体可能突变位点不同,都将成为小麦抗寒育种的优异种质资源;此外,本研究还鉴定出抗寒性由强到弱的系列突变体,有利于进行小麦抗寒基因结构与功能研究,为推动抗寒性基础研究储备了遗传材料。
