热带西太平洋M4 海山超微型浮游生物 的生态分布特点*
2020-10-14赵燕楚张武昌李学刚
赵燕楚 赵 丽 董 逸 张武昌 李学刚 赵 苑 ① 肖 天 ①
(1. 中国科学院海洋生态与环境科学重点实验室(中国科学院海洋研究所) 青岛 266071; 2. 青岛海洋科学与技术(试点) 国家实验室 海洋生态与环境科学功能实验室 青岛 266237; 3. 中国科学院大学 北京 100049; 4. 中国科学院海洋大科学研究中心 青岛 266071)
超微型浮游生物(ultraplankton, <10μm)包括全部的微微型浮游生物(picoplankton, 0.2—2μm)和部分微型真核浮游生物(nanoeukaryotes, 2—20μm, NEUK), 是海洋微食物网的重要组成部分(Murphyet al, 1985)。根据营养类型, 微微型浮游生物主要为微微型自养浮游生物和微微型异养浮游生物, 前者主要包括聚球藻(Synechococcus, SYN)、原绿球藻(Prochlorococcus, PRO)、微微型真核浮游生物(picoeukaryotes, PEUK); 微微型异养浮游生物主要包括细菌和古菌, 二者合 称异养原核生物(heterotrophic prokaryotes, HP)(Campbellet al, 1997)。根据核酸含量的高低, 异养原核生物又分为低核酸含量异养原核生物(low nucleic acid content heterotrophic prokaryotes, LNA)和高核酸含量异养原核生物(high nucleic acid content heterotrophic prokaryotes, HNA) (Gasolet al, 1999)。微型真核浮游生物多样性非常高, 大洋中主要由普林藻纲、硅藻纲、青绿藻纲、浮生藻纲、隐藻纲的浮游生物组成(Xuet al, 2018)。尽管超微型浮游生物的体积较小, 但它们数量巨大、分布广泛, 因此是海洋生物量和生产力的主要贡献者, 也是海洋生态系统物质和能流的主要承担者(Azamet al, 2007)。在寡营养海域超微型浮游生物对总浮游植物生物量的贡献可达90%, 对初级生产力的贡献可达80% (Liet al, 1983; Agawinet al, 2000)。此外, 在大洋中它们可以通过形成聚集体或被微型浮游动物或更高营养级的中型浮游动物摄食, 通过粪便和尸体的沉降, 从表层向深海输送碳(Richardsonet al, 2007); 也可以通过微型生物泵将溶解有机碳转化为惰性有机碳, 在储碳过程中发挥重要作用(Jiaoet al, 2010)。
海山是指自海底升高1000m, 且不露出海面的地形隆起(Rogers, 1994)。海山和洋流的相互作用会形成内波、上升流、泰勒柱、涡旋等物理过程, 这些物理过程可能会导致海山区初级生产力和浮游生物生物量升高, 从而支撑较高的鱼类和底栖生物量, 这一现象被称为“海山效应”(Doweret al, 1996; 张武昌等, 2014)。目前, 海山区浮游生态学研究比较少, 海山对浮游生物的影响也没有一致结论。在热带西太平洋Y3 和M2 海山研究中, 初级生产力、叶绿素a、微食物网生物等都没有在山顶附近发现升高的现象(张文静等, 2016; 代晟等, 2017; 赵丽等, 2017)。Sime- Ngando 等(1992)在东太平洋亚北极的Cobb 海山发现纤毛虫生物量升高。
热带西太平洋表层水温较高、营养盐含量低、受北赤道流影响, 水文环境较为复杂; 此外全球约一半的海山位于太平洋, 其中大多数在热带西太平洋海域(Yessonet al, 2011), 然而目前对该海域的海山调查研究较少。因此, 本研究选取了位于热带西太平洋的M4 海山, 采用流式细胞术对0—300m 水层超微型浮游生物的丰度、生物量及分布特点进行了调查, 以期了解M4 海山超微型浮游生物的分布特征及与环境因子之间的关系, 为深入了解海山区浮游生态特点提供依据。
1 材料与方法
1.1 调查站位
2017 年8 月7 日—9 月5 日乘“科学号”调查船在热 带 西 太 平 洋 M4 海 山(10.3°—10.9°N, 139.9°— 140.4°E)进行样品采集, 共设A、B 两个断面, 分别为东西向和南北向, 交叉于海山顶(图1)。海山山顶距海面57m, 10 号站最深(5999m)。

图1 M4 海山采样站位图 Fig.1 Sampling stations in M4 Seamount
1.2 理化参数的测定
用CTD(Sea-Bird SBE 9, Sea-Bird 公司, 美国)及附带的叶绿素浊度传感器(WET Labs 公司, 美国)测量各站表层到300m 的温度、盐度、水深和叶绿素a荧光值。按海洋调查规范测定营养盐浓度: 在各水层采集水样250mL, 用0.7μm Whatman GF/F 滤膜过滤后装入250mL 高密度聚乙烯瓶, 用体积分数为2‰的氯仿固定后于-20℃中保存, 用QuAAtro 连续流动分 析仪(SEAL 公司, 德国)分析测定溶解无机氮(DIN)和磷酸盐(34PO-)。其中DIN 浓度为硝酸盐(3NO-)、亚硝酸盐(2NO-)和铵盐(4NH+)的浓度总和。
1.3 超微型浮游生物样品采集及测定
在各水层采集海水4mL, 加入10%多聚甲醛固定, 终浓度为1% (V/V), 在室温下避光固定20min 后放于液氮罐中保存, 用于超微型浮游生物丰度的检测。在实验室内将样品在室温下避光融化, 用流式细胞仪(BD FACSJazz, 美国)检测。分析超微型自养浮游生物时, 取2mL 样品, 加入2μm 荧光微球溶液(Polysciences, 美国)作为标准内参, 上机检测3min。根据前向、侧向散射光(与细胞的大小、密度和形态有关)以及红色、橙色荧光信号(与叶绿素、藻红蛋白有关), 可检测到四个超微型自养浮游生物类群(SYN, PRO, PEUK, NEUK)。对于超微型异养原核生物, 取50μL 样品, 加入250μL TE (Tris-EDTA, 100mmol/L Tris-Cl, 10mmol/L EDTA, pH=8.0, Sigma, 美国)缓冲溶液稀释, 再加入 4μL 的 SYBR Green I 染料(Molecular Probes, 美国)避光染色20min 后上机检测30s。根据侧向散射光和绿色荧光(与核酸含量有关)信号, 可检测两个异养原核生物类群(LNA 和HNA)。
1.4 生物量
超微型浮游生物的生物量是由丰度乘以单个细胞的碳转换系数得到。SYN、PRO、PEUK、NEUK、HP (LNA 和HNA)的转换系数分别为: 250、49、671、3498、20fgC/cell (Campbellet al, 1997; Giraultet al, 2013)。
1.5 数据处理与分析
用Surfer 13 和Origin 8.5 软件分析超微型浮游生物的垂直分布情况。用CANOCO for Window 4.5 进行冗余分析(Redundancy analysis, RDA), 研究超微型浮游生物丰度与环境因子之间的相关性, 其中超微型浮游生物丰度的数据经过以10 为底的对数转换。
2 结果
2.1 水文环境
M4 海山0—300m 的水温范围在9.68—31.00°C, 由表层至300m 水层, 温度逐渐降低(图2)。盐度范围在33.00—35.13, 平均值为34.27±0.45。等温线和等盐线在山顶处出现下凹(图2), 在100—200m 水层可观察到温度和盐度跃层。营养盐DIN 和PO43-的平均值分别为5.55±3.00µmol/L 和0.19±0.30µmol/L, 两者的垂直分布趋势一致, 即200m 以浅浓度较低, 200m以深逐渐升高, 在东西方向山顶附近DIN 和PO43-等值线上凸(图2)。
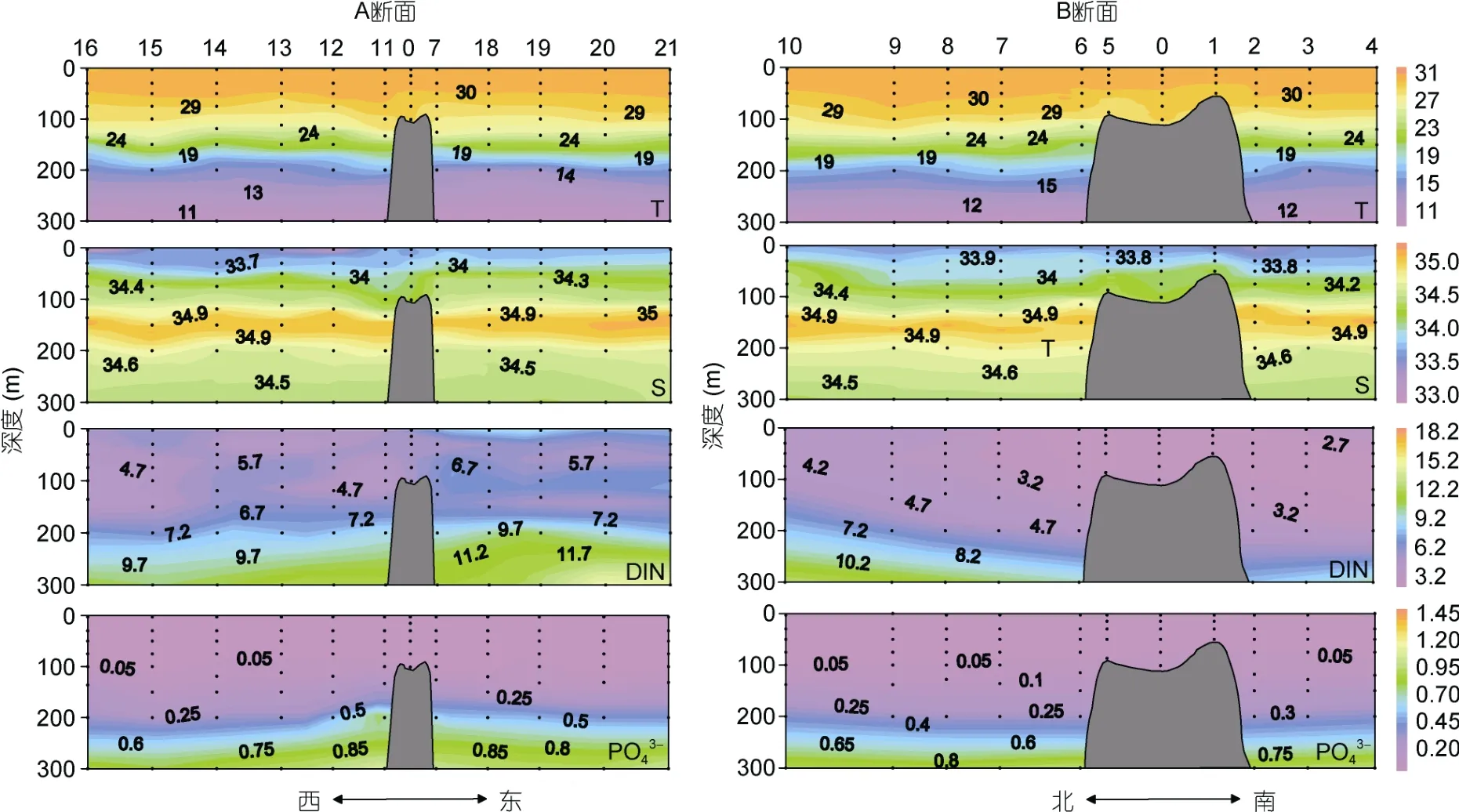
图2 M4 海山A、B 断面温度(T, °C)、盐度(S)、营养盐浓度(DIN, PO 34-, µmol/L)垂直分布 Fig.2 Vertical distributions of temperature (T, °C), salinity (S), nutrient concentrations (DIN, PO 34-, µmol/L) along Transects A and B in M4 Seamount
2.2 超微型浮游生物各类群丰度的分布及其与环境因子的关系
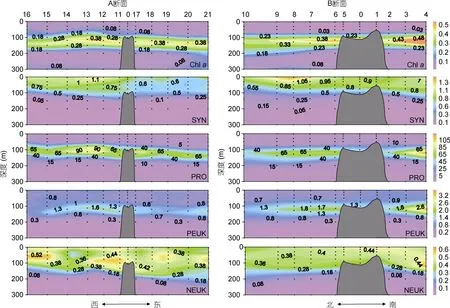
图3 M4 海山A、B 断面叶绿素a 荧光值(chl a), 聚球藻(SYN, ×103cells/mL)、原绿球藻(PRO, ×103cells/mL)、微微型真核浮游生物(PEUK, ×103cells/mL)、微型真核浮游生物(NEUK, ×103cells/mL)丰度垂直分布 Fig.3 Vertical distributions in in-situ chlorophyll a fluorescence (chl a) and abundances of Synechococcus (SYN, ×103cells/mL), Prochlorococcus (PRO, ×103cells/mL), picoeukaryotes (PEUK, ×103cells/mL), and nanoeukaryotes (NEUK, ×103cells/mL) along Transects A and B in M4 Seamount
叶绿素a荧光高值分布在75—200m 水层, 深层叶绿素最大值(Deep Chlorophyll Maximum, DCM)位于100—150m 水层, 与温度和盐度跃层的深度相近(图3)。SYN 的丰度范围在(0.01—1.49)×103cells/mL, 平均值为(0.54±0.40)×103cells/mL, 其垂直分布的高值区在0—100m 水层之间。PRO 是丰度最高的超微型自养浮游生物, 丰度变化范围在(0.0 9—1 5 3.8 9)× 103cells/mL, 平均值为(18.06±30.24)×103cells/mL。 PRO 的垂直分布与叶绿素a荧光相似, 丰度的高值分布在 75—150m 水层之间。PEUK 的丰度范围在(0.00—3.51)×103cells/mL, 平 均 值 为(0.73±0.55)× 103cells/mL。PEUK 的垂直分布趋势与PRO 类似, 丰度高值都位于DCM 层附近, 但PEUK 的高值区略深于PRO, 主要在100—150m 水层。NEUK 丰度远低于其它超微型自养浮游生物, 丰度变化范围在(0.00—0.65)×103cells/mL, 平 均 值 为(0.31±0.19)× 103cells/mL。NEUK 的垂直分布范围比较广, 从表层至150m 水层丰度都较高, 在站位10、19、20、21 丰度高值可深达200m (图3)。
HP 是丰度最高的超微型浮游生物, 丰度范围在(0.73—7.32)×105cells/mL, 平均值为(4.07±1.94)×105cells/mL, 比超微型自养浮游生物高约两个数量级。HP 的丰度在200m 以浅较高, 最高值位于150m 以浅水体。在各站位均检测到LNA 和HNA 两个异养原核生物类群。LNA和HNA 的丰度平均值分别为(1.56±0.74)×105cells/mL和(2.51±1.25)×105cells/mL, 它们的垂直分布与总HP 一致, 高值区均位于150m 水层以浅(图4)。LNA/HNA 的比值范围在0.31—1.41, 平均值为0.65±0.17。LNA/HNA的比值在DCM 层显著高于其它水层。此外, 在DCM层60 个采样点中有7 个该比值超过了1(即LNA 的丰度高于HNA), 这7 个采样点分别在站位2、3、10、16、18 (图4)。
M4 海山超微型浮游生物丰度与环境因子之间的RDA 分析结果显示, 第一和第二轴共解释了75.9%的超微型浮游生物的变化, 以及99.7%的超微型浮游生物与环境因子的关系(图 5)。SYN、NEUK、PEUK、LNA、HNA 与温度呈正相关关系, 与深度、PO43-、DIN 呈负相关关系; PRO 与叶绿素a荧光值呈正相关关系。LNA 占总异养原核生物的百分比与叶绿素a荧光值呈正相关关系。
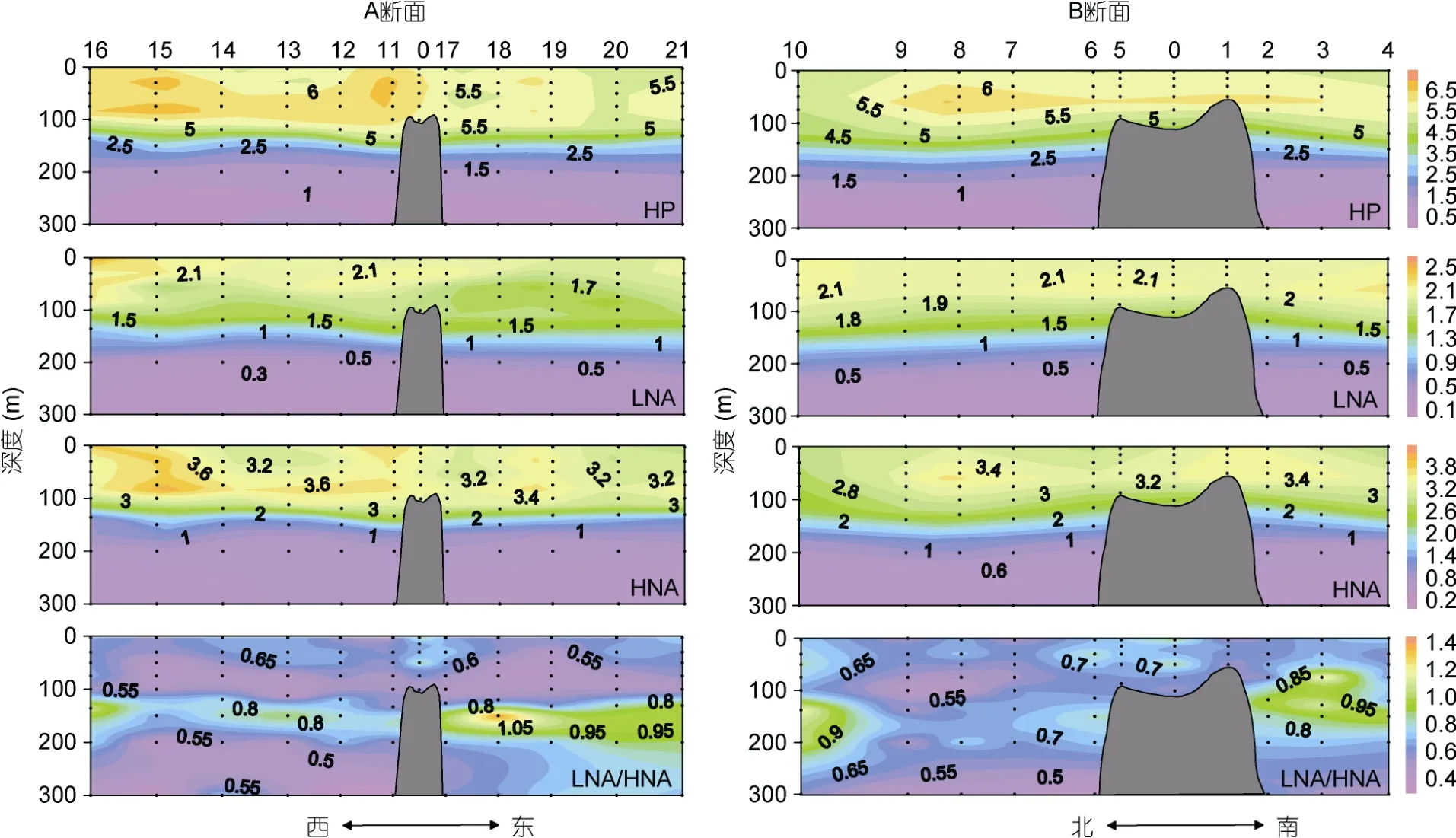
图4 M4 海山A、B 断面总异养原核生物(HP, ×105cells/mL)、低核酸含量异养原核生物(LNA, ×105cells/mL)、高核酸含量异养原核生物(HNA, ×105cells/mL)丰度, 低核酸含量与高核酸含量异养原核生物丰度比值(LNA/HNA)垂直分布 Fig.4 Vertical distributions in abundances of total heterotrophic prokaryotes (HP, ×105cells/mL), low nucleic acid content heterotrophic prokaryotes (LNA, ×105cells/mL), high nucleic acid content heterotrophic prokaryotes (HNA, ×105cells/mL), and the ratio of low and high nucleic acid content heterotrophic prokaryotes abundances (LNA/HNA) along Transects A and B in M4 Seamount
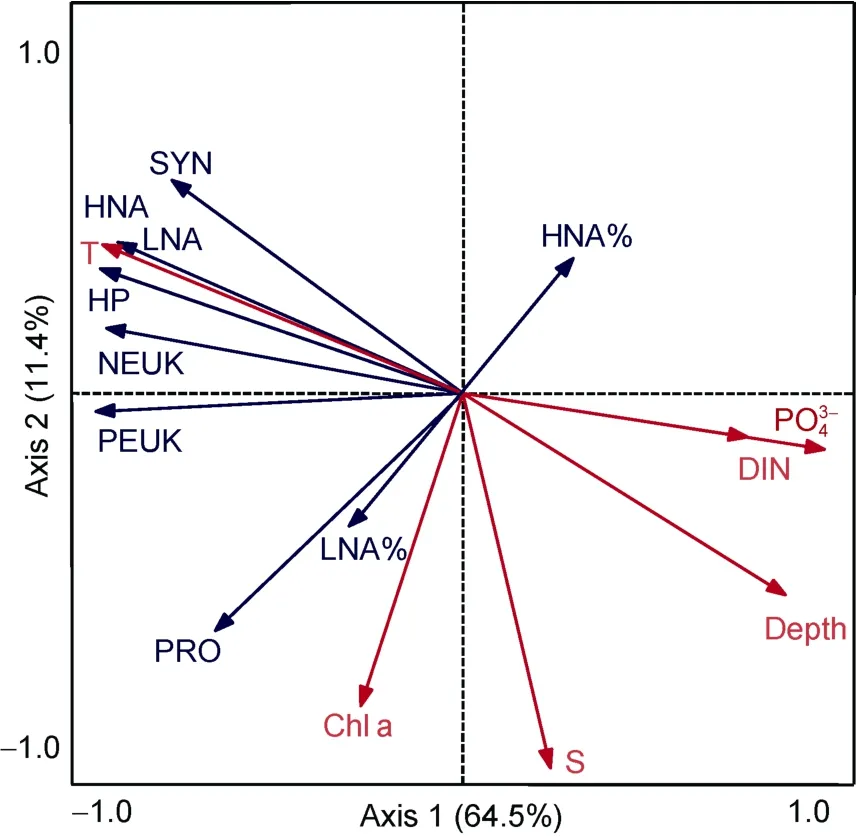
图5 300m 以浅超微型浮游生物与环境因子之间的冗余分析 Fig.5 Redundancy analysis between ultraplankton and environmental parameters in the upper 300m water column
2.3 超微型浮游生物各类群生物量及百分比
超微型浮游生物总生物量平均值为11.23±5.20μgC/L, 最高值为100m 水层的16.31μgC/L, 最低值为300m水层的1.73μgC/L, 其中异养原核生物的生物量远高于超微型自养浮游生物。
超微型自养浮游生物的生物量范围在 0.05— 6.02μgC/L 之间, 平均值为2.69±1.92μgC/L。130m 以浅生物量随深度增加逐渐升高, 130m 以深生物量随深度增加逐渐降低(图6)。超微型自养浮游生物各类群有不同的垂直分布模式, 生物量的主要贡献者在不同水层发生变化。75m 以浅随深度增加SYN 生物量逐渐降低, PRO 逐渐升高, PEUK 和NEUK 生物量分布比较均匀; NEUK 在超微型自养浮游生物中所占百分比最高为 58.62%±8.53%, 其次是 PEUK 为20.22%±2.68%, SYN 和PRO 所占百分比较低分别为8.75%±2.19%和12.41%±13.28%。100—150m (DCM)水层中SYN 生物量极低, PRO 和PEUK 明显升高, NEUK 略有降低; PRO 的生物量超过NEUK, 在超微型自养浮游生物中占优势(53.83%±6.32%)。DCM 以深(200—300m)各类群生物量都明显下降, NEUK 是生物量的主要贡献者(46.18%±7.82%)。
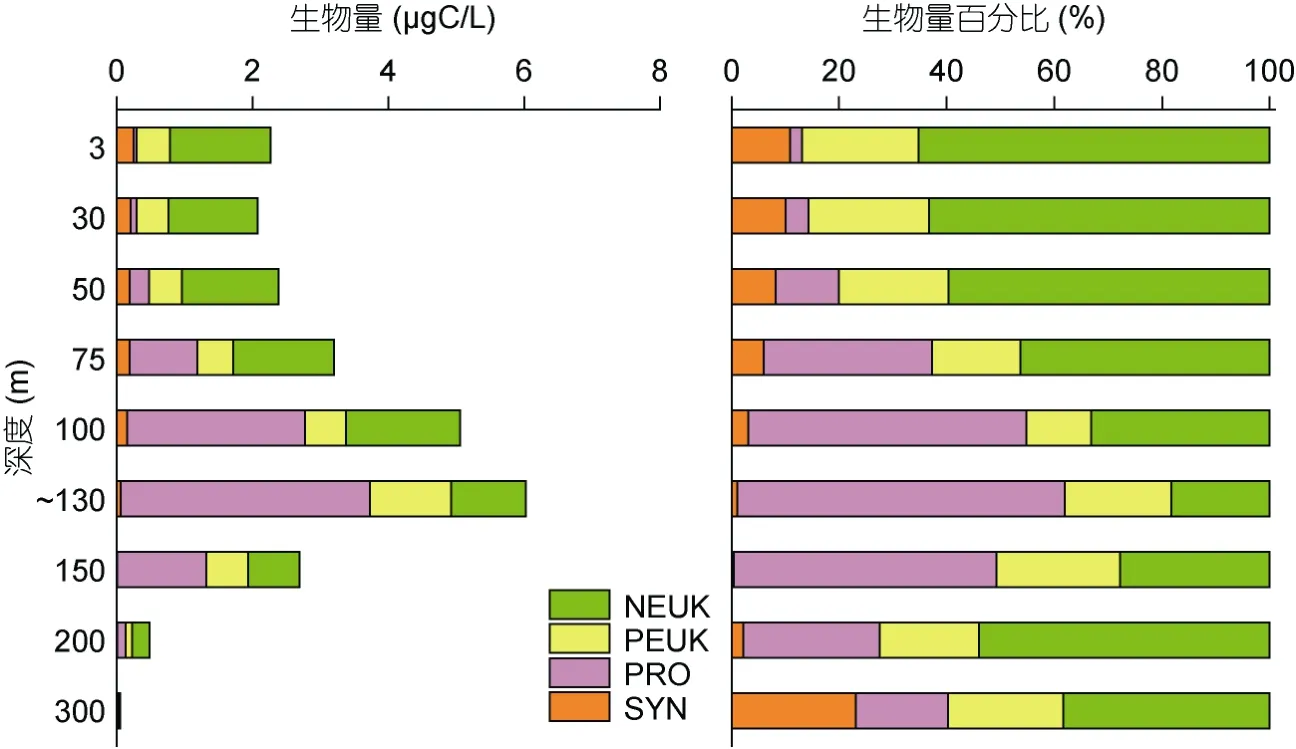
图6 M4 海山超微型自养浮游生物的生物量及百分比的垂直分布 Fig.6 Vertical distributions of autotrophic ultraplankton biomass and percentage in M4 Seamount
HP 生物量范围为1.68—11.25μgC/L, 平均值为8.54±3.28μgC/L。100m 以浅HP 生物量相对较高, 100m 以深生物量随深度增加逐渐降低。在HP 中LNA 和 HNA 生物量平均值分别为 3.29±1.22μgC/L 和5.25±2.32μgC/L, 二者的垂直分布与总 HP 一致。LNA 在总异养原核生物中所占百分比范围在34.18%(300m)—46.59%(150m), 平均值为38.95±3.98%; HNA 百分比范围在53.41%(150m)—65.82%(300m), 平均值为61.05±3.98%, 总体来说LNA 所占的比例低于HNA(图7)。
3 讨论
3.1 M4 海山超微型自养浮游生物生物量的分布及影响因素
本研究中超微型浮游生物各类群的丰度与以往大洋中报道的结果具有可比性。M4 海山SYN、PRO、PEUK、NEUK、HP 丰度值与热带和亚热带西太平洋、热带西太平洋 Y3 和 M2 海山、亚热带北太平洋ALOHA 站、大西洋北部环流等海域检测值相近, 略低于北太平洋副热带环流西部和赤道太平洋海域(表1)。
超微型自养浮游生物中生物量的主要贡献者因海域等不同而发生变化。在低纬度海域PRO 和PEUK对总生物量贡献较大, SYN 和NEUK 相对较小。Painter 等(2014)在亚热带北大西洋东部(26.2°N, 31.1°W)研究发现100m 以浅PRO 是生物量的主要贡献者, 其次是PEUK; 在100—150m 水层PRO 的贡献有所下降, 但仍高于 PEUK; 在 150—200m 水层PEUK 生物量的贡献超过PRO。在亚热带北太平洋ALOHA 站位(22°45′N, 158°W), 表面混合层(0—50m)、DCM 层(114±18m)、DCM 层以深(120—175m)PRO 生物量的贡献都要高于其他自养类群(Campbellet al, 1997)。然而, 一些研究发现在高纬度海域NEUK 对总生物量的贡献也较高。Tarran 等(2001)在东北大西洋(36°42′—59°18′N, 19°12′—20°12′W)100m 以浅研究发现NEUK 的生物量贡献最高, 可达61.20%。Van Oostende 等(2017)在靠近亚北极(54°N, 20°W)的北大西洋发现NEUK 对微微型和微型自养浮游生物的贡献高达86%。本研究结果显示在热带西太平洋M4海山(10.3—10.9°N, 139.9—140.4°E), NEUK 在75m以浅(58.62%)和200—300m 水层(46.18%)也是生物量的主要贡献者, 100—150m 水层生物量的主要贡献者是PRO。
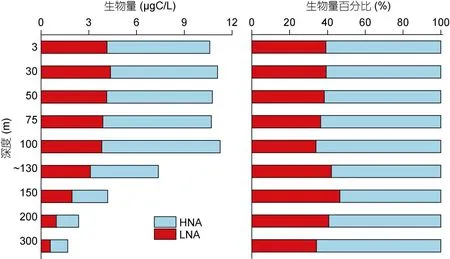
图7 M4 海山低核酸含量和高核酸含量异养原核生物的生物量及百分比的垂直分布 Fig.7 Vertical distributions of low and high nucleic acid content heterotrophic prokaryotes in biomass and percentage in M4 Seamount
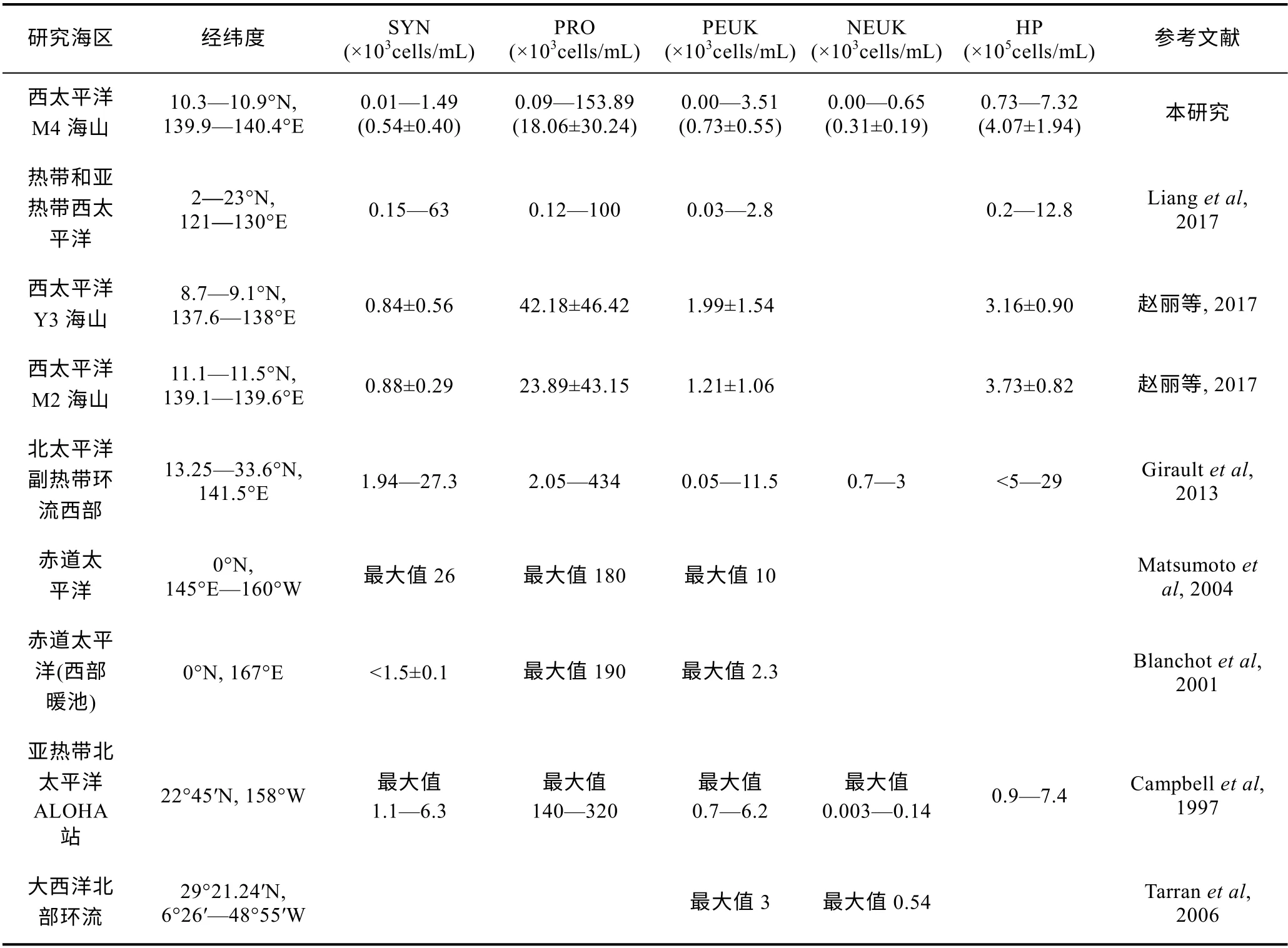
表1 不同海区超微型浮游生物丰度的比较 Tab.1 Comparison in abundance of ultraplankton in different regions of the global oceans
温度、光照和营养盐等环境因子是影响PRO 和NEUK 生物量分布的重要因素。M4 海山位于寡营养的热带海域, 光照较强, 表层水温达到30°C; 水体长期存在物理分层, 上层营养盐含量较低。由于PRO的光系统 II 的热敏感性, 其适宜的生长温度高于15°C; 在表层水中强光照对PRO 的生长有抑制作用(Otero-Ferreret al, 2018)。因此, 温度适宜, 光照较弱且营养盐含量相对较高的DCM 层更适合PRO 的生长。NEUK 生物多样性较高, 能够适应不同的光照、温度等环境条件(Marieet al, 2010)。此外, NEUK 细胞体积较大, 碳含量高。这两方面因素共同导致NEUK生物量在表层和深层占优势。RDA 研究结果显示超微型自养浮游生物(SYN, PEUK, NEUK)与营养盐呈负相关关系, 一是由于超微型自养浮游生物体积小, 在寡营养环境中更有竞争优势; 二是深层海水中的营养盐含量较高, 但温度低、光照弱不利于超微自养浮游生物的生长, 温度和光照是深层海域中主要的限制因素(Agawinet al, 2000)。
3.2 LNA 和HNA 生物量比例的变化及影响因素
根据流式细胞术的绿色荧光和侧向散射光通常可以将水生生态系统中的异养原核生物分为2 或3 个类群(Gasolet al, 1999; Giraultet al, 2015)。本研究中, 我们检测到2 个具有相似的侧向散射光, 不同荧光强度的类群: LNA 和HNA。从淡水到咸水、寡营养到富营养等多种环境中都有 LNA 和 HNA 的报道(Nishimuraet al, 2005; Otero-Ferreret al, 2018)。LNA和HNA 的生态效应有差异。HNA 细胞体积较大, 通常被认为是生长活跃的细菌, 能够对捕食压力和营养盐变化做出迅速反映。而最初的研究认为LNA 可能是不活跃或者已经死亡的细菌(Gasolet al, 1999), 后来研究表明在一些寡营养环境中, LNA 也具有代谢活性, 是异养原核生物的重要组成部分, 对碳生物量有重要贡献(Zubkovet al, 2001; Jochemet al, 2004)。海洋LNA 和HNA 的类群组成也有显著差异, HNA 主要由红杆菌、拟杆菌和SAR116 组成, 而其他群体如SAR11 和SAR86 是LNA 主要组成部分(Vila-Costaet al, 2012)。目前对这两个类群的生态作用分析研究有限, 有待于深入研究。
根据以往报道, 不同海域中异养原核生物的优势类群不同。HNA 通常在高生产力的近岸海域占优势, 如南澳大利亚大陆架上升流区、地中海沿岸、加利西亚沿岸上升流区(Gasolet al, 1999; Patersonet al, 2012; Otero-Ferreret al, 2018)。LNA 通常在北太平洋、大西洋中部等寡营养大洋环境中占优势(Liet al, 1995; Giraultet al, 2015)。本研究中, 各站位HNA 生物量百分比平均值为61.05%, 远高于LNA, 与近岸研究结果比较相似(表2), 但在DCM 层内, LNA 生物量的比例明显升高, 尤其是在海山附近可高达58.64%。这种DCM 层内LNA 占优势的情况在地中海西北部也有出现。Scharek 等(2007)在地中海西北部表层和DCM 层中LNA 和HNA 的生长率和被摄食率研究发现, 表层HNA 的生长率和被摄食率均显著高于LNA, 而DCM 层LNA 的生长率和被摄食率高于HNA, 但差异并不显著; HNA 是表层水体中活性较高的组成部分, LNA 在DCM 层较为活跃。
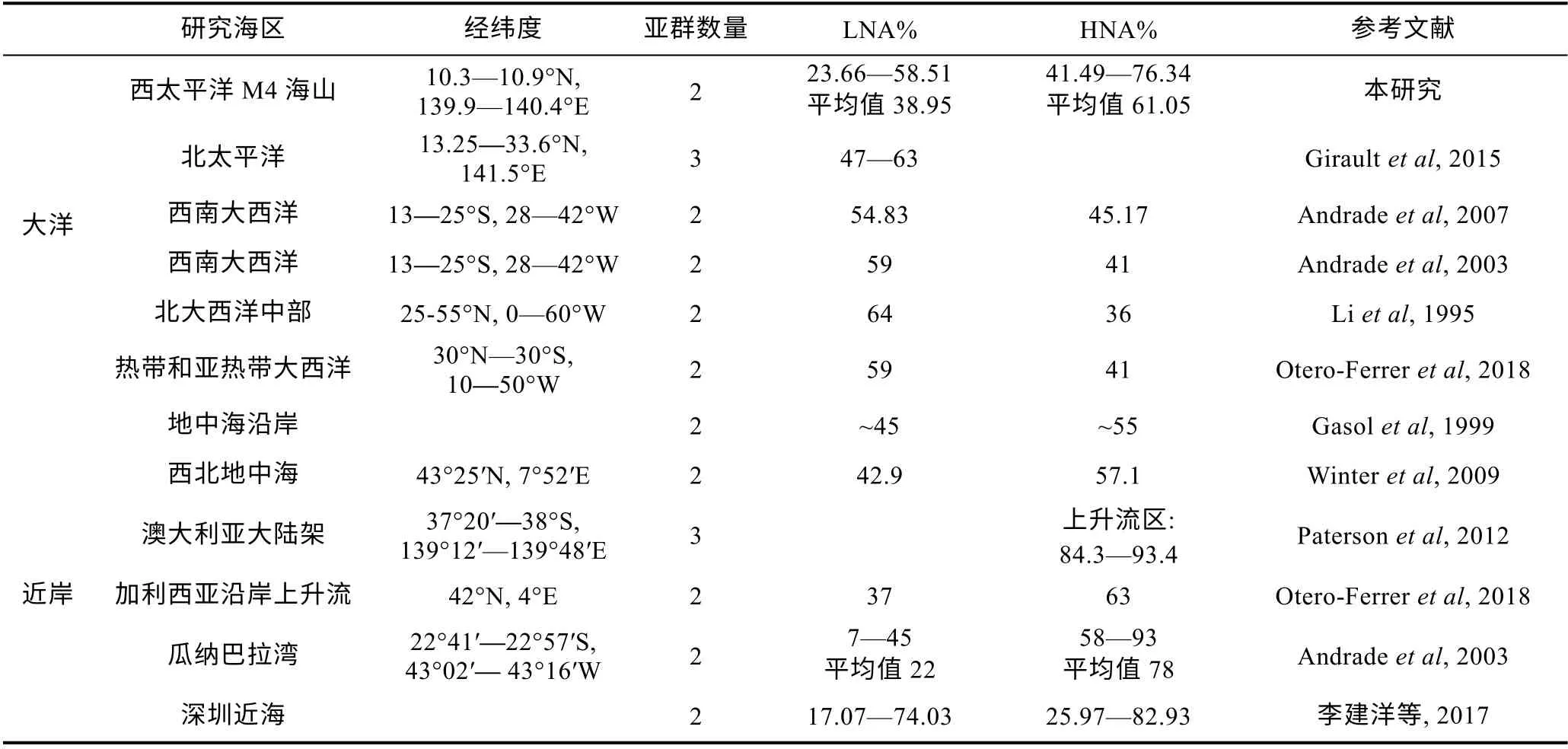
表2 不同海区低核酸含量和高核酸含量异养原核生物占总异养原核生物百分比的比较 Tab.2 Comparison in percentage of low and high nucleic acid contents of heterotrophic prokaryotes in different regions of the global oceans
异养原核生物的分布受环境和生物两方面因素的影响。M4 海山是一座典型的浅海山, 山顶距海面约57m。已有研究表明浅海山与海流的相互作用会产生水文动态变化, 产生内波、上升流、涡旋、泰勒柱等, 如在东北太平洋Cobb 海山、西北太平洋Minami-Kasuga海山、西南印度洋六座海山(Geninet al, 1985; Doweret al, 1992; Readet al, 2017)。虽然目前没有关于M4 海山环流的直接数据, 但Ma 等(2019)在M4 海山(即该文中的C4 海山)观察到了营养盐等值线在山顶附近有明显的抬升, 说明营养盐存在一个自下而上的抬升过程; 同时与远离M4 海山的参考站位相比, 海山区营养盐的浓度较高。这些现象表明M4 海山可能存在将低温高营养盐的深层水带到浅层的局部扰动或上升流。水文条件的变化会对LNA、HNA 生长产生影响。Andrade等(2007)在西南大西洋(13—25°S, 28—42°W)19°S 以南的区域发现LNA 占优势, 并认为这与该地区存在涡旋和上升流有关。因此在M4 海山上升流的存在可能是海山附近LNA 生物量比例升高的原因之一。此外, 病毒的侵染、裂解也会对异养原核生物亚群的比例产生影响。已有实验证明HNA 更容易受到病毒的侵染、裂解(Bonilla-Findjet al, 2009)。在M4 海山区浮游病毒的丰度在海山站显著高于大洋站(Zhaoet al, 2020), 可能会导致海山站HNA 死亡率较高, 从而表现为LNA生物量的比例升高。
3.3 M4 海山对超微型浮游生物的影响
M4 海山超微型自养浮游生物和异养原核生物各类群均没有发现丰度和生物量有明显升高或等值线波动现象, 即没有观察到明显的“海山效应”; 而海山影响了异养原核生物两个类群生物量的比例, LNA 的比例在海山附近升高。以往研究表明, 少数情况下在海山区能够观察到高叶绿素浓度、高浮游植物丰度等现象, 但这些现象也只是短暂的或季节性的(Rogers, 2018)。Genin 等(1985)在Minami-Kasuga 海山进行三次调查, 只有第一次在海山上方观察到了冷堆、叶绿素浓度增加以及高浮游动物生物量。此外, 海山并非对所有的浮游生物都能产生影响。Mendonça 等(2012)对东北大西洋Seine 和Sedlo 海山的微生物类群调查发现: 在Seine 海山的水体中, 只在春季观察到大的自养生物(除PRO 外)在山顶和海山站升高; 在Sedlo海山的水体中, 尽管微微型浮游生物的生物量较低, 但在夏季山顶和海山区有其峰值存在。海山区浮游生物的分布受采样时间、山顶距海面的深度、山体的形状、洋流的强度和稳定性、周围水体的温度、盐度以及营养盐等多种因素的影响。因此, 海山对超微型浮游生物的分布影响需要更多的调查进一步验证。
4 结论
本研究发现M4 海山超微型自养浮游生物的生物量的主要贡献者随水深发生变化。NEUK 在75m 以浅和 200—300m 水层是主要贡献者, PRO 在100—150m 水层是主要贡献者。超微型自养浮游生物各类群分布没有明显的“海山效应”。在异养原核生物中HNA 生物量所占百分比高于LNA, 然而海山附近DCM 层内LNA 生物量比例明显升高。海山的存在会对异养原核生物两个类群生物量的比例产生影响。目前海山区超微型浮游生物的研究比较少, 海山对超微型浮游生物的影响还需要更深入系统的研究。
