秋水仙素对菊苣多倍体的诱导
2020-09-24赵建华任国辉郑金双
赵建华 任国辉 郑金双

摘要:利用不同质量浓度秋水仙素溶液对菊苣生长点进行不同次数的诱变处理。通过观察气孔特征、测定DNA相对含量进行诱变植株筛选鉴定,确定其倍性水平。结果表明,0.2%秋水仙素处理4次,菊苣诱导率最高,为8.16%,获得8株菊苣同源四倍体植株,与二倍体对照相比,变异植株叶片增厚,叶色加深,叶片褶皱,下表皮气孔变大,保卫细胞内叶绿体数量增加,叶绿素含量增加,单位面积气孔数目减少。本试验共获得23株变异植株,其中22株同源四倍体,1株混倍体,构建了菊苣多倍体诱导技术体系,获得了菊苣多倍体新种质,丰富了菊苣遗传基础,为良种选育提供了材料。
关键词:菊苣;秋水仙素;诱导;同源四倍体
中图分类号:S567.23+9.035.2
文献标志码:A
文章编号:1002-1302(2020)16-0177-06
菊苣(Cichorium intybus L.)為菊科菊苣属多年生草本植物,营养价值高,抗逆性强。以干燥地上部分及根入药,味微苦、咸,具有清肝利胆、健胃消食、利尿消肿等功效[1]。菊苣叶片柔嫩多汁,营养物质丰富,研究表明,其叶丛期含粗蛋白22.87%,初花期含粗蛋白14.73%、粗脂肪2.1%、粗纤维30.6%,被称为绿色健康之王[2]。目前,国内关于菊苣的研究多集中于栽培技术[3-4]、活性成分测定及提取物的药理作用[5-6]等方面,而对于菊苣遗传育种方面的研究较少,张玉等利用60CO-r辐照菊苣种子发现,高剂量辐照对菊苣诱变效果明显[7],但诱变是正向还是逆向则需进一步在农艺性状方面作深入研究。
多倍体植物大多表现为营养器官的“巨大性”,具有生长健壮、抗逆性增强、果实增大以及营养物质含量提高等优点,因此多倍体诱变育种已成为植物育种的重要方式,目前已在大田作物、蔬菜作物、观赏植物、药用植物育种方面得到广泛应用。隗志松等研究发现,多倍体水稻植株高大粗壮,穗大粒大,稻米质优,抗逆性强[8]。茆吉健利用秋水仙素诱变处理获得四倍体萝卜,产量较二倍体增加26.92%,并且根内可溶性蛋白和可溶性糖等营养成分较二倍体植株显著增加[9]。宋莲等利用秋水仙素对墨兰×大花蕙兰的F1进行诱变处理,获得的多倍体植株观赏价值高和抗逆性强[10]。王凤宝等采用秋水仙素浸泡枸杞干种子,获得了对瘿螨免疫,高抗白粉病和根腐病,营养品质和药用成分(甜菜碱、枸杞多糖、黄酮类)显著高于二倍体对照的多倍体新种质[11]。
本试验采用不同质量浓度的秋水仙素溶液对子叶期菊苣进行不同次数的诱变处理,以期构建菊苣多倍体诱导技术新体系,丰富菊苣种质资源。
1 材料与方法
1.1 试验材料
菊苣(Cichorium intybus L.)种子,购于承德兴春和农业集团股份有限公司。
1.2 试验方法
本试验采用随机区组设计,2因素(不同秋水仙素浓度)3水平(不同处理次数)处理。
第1次诱变处理于2017年10月初进行。将菊苣种子直播于50孔育苗穴盘中,置于棚中育苗,待菊苣2张子叶完全展开后,将棉球置于两子叶中间的生长点处,利用不同质量浓度秋水仙素(0.05%、0.10%、0.20%)分别于06:00—07:00和17:00—18:00处理3、5、7次。以蒸馏水处理为对照,每个处理50株,2次重复,待所有处理结束后,摘去棉球,继续育苗。
第2次诱变处理于2018年3月中旬,第3次于10月初在智能日光温室中进行试验。处理方法同第1次处理,利用0.1%和0.2%秋水仙素溶液分别处理不同次数(4、6、8次),每个处理50株,3次重复。
1.3 多倍体植株的筛选与鉴定
1.3.1 气孔鉴定 当植株有完全展开的3张叶时,取二倍体和诱变植株完全展开的叶片,用镊子撕取下表皮,置于滴有1滴蒸馏水的载玻片上,盖片,赶出气泡,吸去多余水,于Olympus BX53光学显微镜下观察,20×镜下选取10个视野,观察气孔大小、单位面积气孔数目并记录,筛选气孔变异的植株为候选植株。
气孔变异植株比例=气孔鉴定变异株数/处理株数×100%。
1.3.2 DNA相对含量鉴定 利用流式细胞仪鉴定气孔以获得候选植株进行NDA相对含量鉴定,剪取二倍体和诱变植株完全展开的幼嫩叶片,放入装有500 μL裂解液的预冷培养皿中,刀片快速切碎,用260~560目的尼龙网过滤到离心管中,放置于冰中 5 min,加入100 μL PI染色液,于冰上避光染色 12 min,染色后移至上样管中,利用BD LSR FortessaTM流式细胞仪检测,确定其倍性水平,并对获得的变异植株编号,T表示同源四倍体(T-1至 T-22),H表示混倍体(H-1)。
根据秋水仙素处理后成活株数计算致死率,依据上述鉴定方法获得的变异株数计算诱导率。
致死率=死亡株数/处理幼苗总数×100%;
诱导率=变异株数/处理幼苗总数×100%
1.4 二倍体与变异植株的比较
1.4.1 叶片大小及厚度的测量 在植株有5张完全展开叶时,取二倍体与变异植株的完全展开叶,用直尺测量叶长和叶宽,再利用打孔器取中叶脉两侧的叶片,用游标卡尺测量10张小叶片的厚度。
1.4.2 气孔大小的比较 取二倍体和变异植株完全展开的叶片,利用气孔鉴定的制片方法,在显微镜下观察,40×镜下选取5个视野,测量气孔的长和宽,记录气孔保卫细胞中叶绿体数目。
1.4.3 叶绿素含量测定 称取0.2 g成熟叶片放入研钵中,加入95%乙醇研成匀浆,移至10 mL容量瓶中定容,3 000 r/min离心5 min,取上清液,加95%乙醇稀释,利用分光光度计于662、644 nm下比色,记录吸光度D值并计算叶绿素含量。
叶绿素含量(mg/L)=5.13×D662 nm+20.44×D644 nm;叶绿素相对含量(mg/g)=叶绿素浓度(mg/L)×叶绿素溶液体积(mL)/叶片质量(g)×1 000。
2.5 数据分析
利用Excel表格和DPS软件进行数据整理分析。
3 结果与分析
3.1 秋水仙素诱导条件筛选
由表1可知,在第1次诱变中,植株变异率最高为31.00%(0.20%秋水仙素处理7次),最低为13.00%(0.05%秋水仙素处理3次)。由于棚中高温低湿的条件不利于幼苗生长,导致幼苗死亡率极高,随着秋水仙素浓度的增加,致死率逐渐升高,0.20%秋水仙素处理7次致死率最高,为100%,0.05%秋水仙素处理3次致死率最低,为87.00%。经比较发现,0.05%秋水仙素处理生长点后植株形态变异数量最少;不同质量浓度处理3次时,植株形态变异数量均少于其他处理次数。因此,秋水仙素浓度为0.05%,处理次数为3次时,均不适合作为秋水仙素诱导菊苣多倍体的处理条件。
与第1次诱变处理相比发现,第2次诱变处理的植株形态变异的株数较多,变异率增加。形态鉴定结果表明,0.20%秋水仙素处理8次,变异率最高,为49.67%,0.10%秋水仙素处理4次,变异率最低,为35.33%。0.20%秋水仙素处理6次、8次致死率最高,均为31.67%;0.10%秋水仙素处理4次致死率最低,为13.67%。DNA相对含量鉴定共获得16株变异植株,诱导率最高为5.33%(0.20%秋水仙素处理4次),诱导率最低为1.67%(0.10%秋水仙素处理6次、0.20%秋水仙素处理8次)。移栽至智能日光温室后,高温低湿的条件导致病虫害发生严重,变异植株死亡率极高。
3.2 秋水仙素对菊苣诱变效果的影响
由表2可知,不同质量浓度的秋水仙素和不同次数处理之间相比较,随着浓度和处理次数的增加,诱导率和致死率均呈增加趋势。气孔鉴定的结果表明,0.2%秋水仙素处理4次和6次变异率较高,分别为8.00%和6.67%,与其他处理相比差异显著。0.1%秋水仙素处理6次致死率最低,为1.00%;0.2%秋水仙素处理8次致死率最高,为10.26%。DNA相对含量鉴定结果表明,0.2%秋水仙素处理4次和6次诱导率较高,分别为8.16%(8株)和7.14%(7株),与其他处理相比差异显著;0.1%秋水仙素处理4次诱导率最低,为0.99%(1株)。
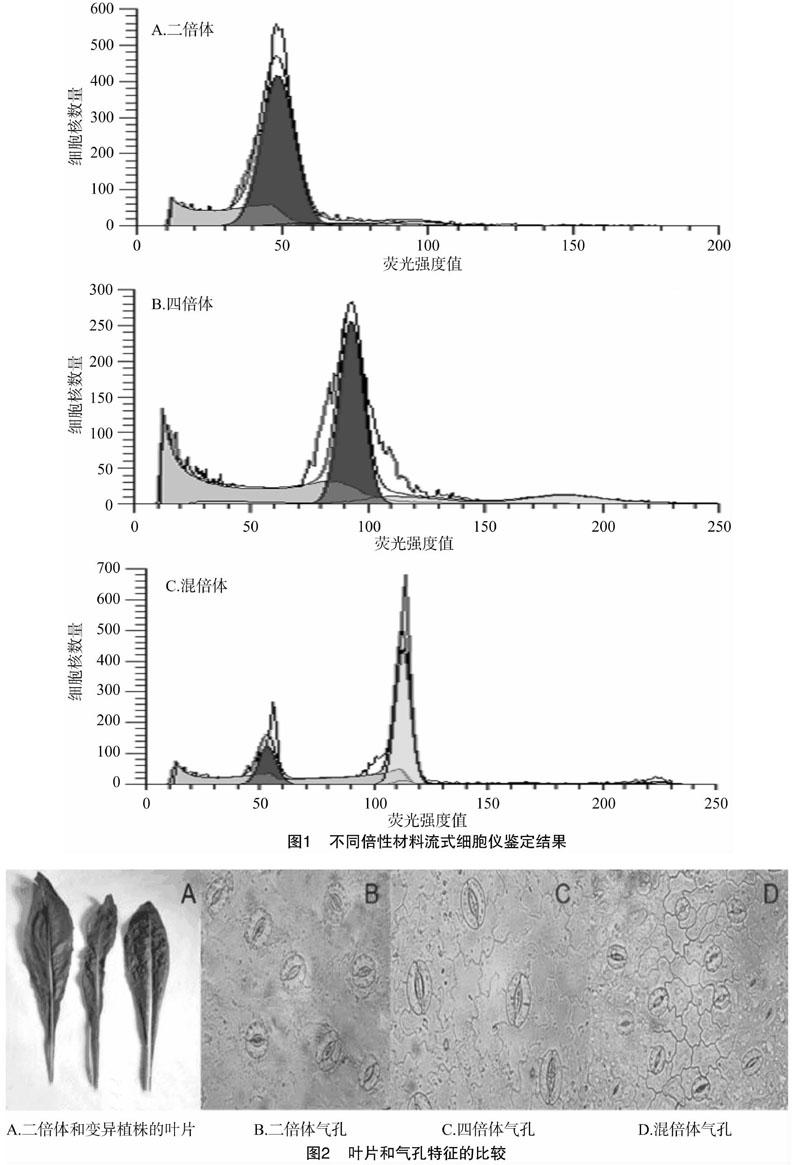
利用流式细胞仪对不同倍性的菊苣进行鉴定(图1),横轴为荧光强度值,纵坐标为细胞核数量,不同倍性菊苣的荧光强度峰值不同。二倍体荧光强度值为50时,细胞核数量达到最大值;四倍体荧光强度值为100时,细胞核数量达到最大值;混倍体在荧光强度值为50和100时,均出现峰值。
3.3 二倍体与多倍体菊苣的比较
3.3.1 叶片特征、气孔特征及叶绿素含量比较 变异植株的叶色加深,叶片褶皱(图2-A);四倍体气孔增大,单位面积气孔数目减少,混倍体气孔大小不一(图2-B、图2-C、图2-D)。由表3可知,变异植株的叶长和叶宽均显著低于二倍体植株。所有四倍体植株的叶厚均大于二倍体植株,其中 T-1、T-2、T-5、T-6、T-7、T-13、T-19、T-20和H-1与对照相比差异不显著。所有变异植株的叶绿素含量均显著高于二倍体植株,不同变异植株间叶绿素含量差异较大,其中T-6、T-7、T-10、T-12、T-16和 H-1葉绿素含量较高。
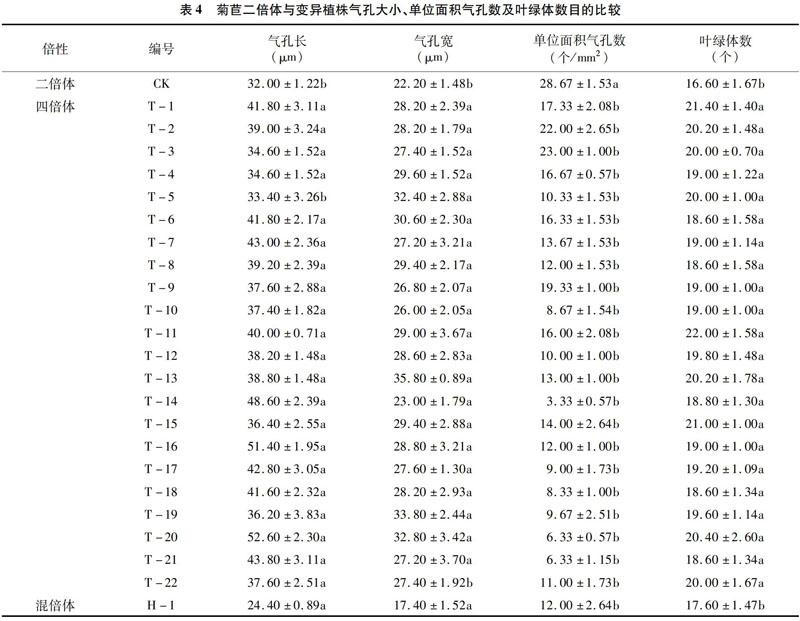
3.3.2 菊苣气孔大小、单位面积气孔数目及叶绿体数目的比较 由表4可知,变异植株(除F5、H-1)的气孔长度显著大于二倍体植株,其中T-5与二倍体差异不显著;变异植株气孔宽度均小于二倍体植株,其中T-22与二倍体差异不显著。变异植株叶片下表皮单位面积气孔数目与二倍体相比减少19.78%~88.39%,与二倍体植株存在显著差异。四倍体植株叶片下表皮保卫细胞内叶绿体数目显著的多于二倍体植株,增加了12.05%~32.53%,但混倍体植株与二倍体植株差异不显著。
变异植株之间单位面积气孔数目差异不显著,其中T-10、T-14、T-17、T-18、T-20、T-21和H-1的单位面积气孔数目;T-2和T-3单位面积气孔数目较多。
4 结论与讨论
秋水仙素是人工诱导植物多倍体常用的药剂,在细胞有丝分裂期间通过抑制纺锤体的形成从而实现染色体的数目加倍[12-13],但高浓度的秋水仙素可以致使材料的生长受到抑制或死亡。殷婷婷利用不同浓度秋水仙素和通过不同处理时间以诱导油菜种子,发现0.4%秋水仙素处理48 h时死亡率最高为63%[14]。彭绿春等利用不同浓度秋水仙素和不同时间浸泡腋花杜鹃无菌苗,发现秋水仙素浓度为0.25%处理72 h死亡率最高为60%[15]。孙嘉曼等采用不同浓度秋水仙素对野生蕉愈伤组织进行不同时间诱变处理,以0.2%浓度处理15 d时致死率最高,为43.62%[16]。本试验发现秋水仙素浓度越高,处理次数越多,幼苗致死率越高,其中0.2%秋水仙素处理8次时,致死率最高,为10.26%。
筛选最佳的秋水仙素浓度和处理时间,是获得多倍体植物的关键因素。孔维一等采用秋水仙素处理一串红幼苗茎尖,发现0.1%秋水仙素处理2 d时诱变率最高,为13.3%[17]。吴照云等利用秋水仙素处理胭脂萝卜幼苗生长点,表明以2 g/L秋水仙素诱变率最大,为33.3%[18]。邓云等采用秋水仙素处理二倍体不结球白菜,表明以0.1 mol/L秋水仙素处理6次效果最好,诱导率达17.5%[19]。本研究以不同质量浓度秋水仙素处理不同次数来诱导菊苣幼苗生长点,结果表明,0.2%秋水仙素处理4次菊苣诱变效果最好,诱导率为8.16%。
同源四倍体植株叶片厚度、气孔大小、叶绿素含量、保卫细胞内叶绿体数均高于二倍体植株,而气孔密度低于二倍体植株[20]。漆慧娟比较不同倍性的甜叶菊发现,四倍体植株气孔密度比二倍体减少49.16%,保卫细胞内叶绿体数较二倍体增多78.49%,气孔长和宽比二倍体分别增加72.25%和69.25%,叶绿素含量也显著高于二倍体[21]。钟程等比较不同倍性水平贵州野生淡黄花百合,发现四倍体植株气孔比二倍体明显增大,单位面积气孔数目减少,且气孔长宽及单位面积气孔数均达到极显著水平[22]。吴顺等比较不同倍性水平的紫花丹参,发现多倍体叶绿素含量比二倍体提高近75%[23]。本研究中,菊苣变异植株叶片下表皮单位面积气孔数较二倍体减少11.61%~55.22%,叶绿素含量比二倍体提高近23.46%,四倍体保卫细胞内叶绿体数较二倍体增加84.56%,叶片厚度及气孔大小均与二倍体存在显著差异。
本试验得到的变异植株,其叶长和叶宽显著低于二倍体,其原因可能是秋水仙素的毒害作用,使菊苣F1植株生长缓慢,叶片褶皱,叶面积减小,若收获四倍体植株的种子后继续种植,其叶面积可能增大。
参考文献:
[1]国家药典委员会. 中国人民共和国药典(一部)[M]. 北京:中国医药科技出版社,2015:310.
[2]池惠武,郭 璞,王运琦,等. 菊苣的价值及研究现状分析[J].南方农业,2018,12(5):9-10.
[3]敖志东. 京冀地区菜用粉用菊苣高产栽培技术[J]. 农业与技术,2016,36(14):128-129.
[4]任冰如,陈 剑,梁呈元,等. 南京地区菊苣品种“沃姆”引种栽培初报[J]. 中国野生植物资源,2018,37(3):76-79.
[5]热阳古·阿布拉,夏 娜,姑丽各娜·买买提依明,等. HPLC法测定菊苣中芦丁、槲皮素和山柰酚含量[J]. 食品工业,2019,40(3):267-270.
[6]韩 畅,党 婷,王 茹,等. 菊苣根、茎的乙醇提取物对金黄色葡萄球菌及粪肠球菌抑菌差异性研究[J]. 中国药理学通报,2019,35(4):540-545.
[7]张 玉,白史且,李达旭,等. 60Co-γ辐射对菊苣种子发芽及幼苗生理的影响[J]. 草地学报,2013,21(1):147-151.
[8]隗志松,舒园园. 优良多倍体水稻资源的创建与现有多倍体材料改良的方法及途径[J]. 现代农业科技,2017(13):25-26.
[9]茆吉健. 不结球白菜、萝卜同源四倍体新种质的创制[D]. 南京:南京农业大学,2016.
[10]宋 莲,杨俊旭,刘 丹,等. 墨兰‘绿墨素×大花蕙兰‘世界和平F1代多倍体诱导初报[J]. 广西植物,2018,38(2):188-194.
[11]王凤宝,付金锋,董立峰,等. 多倍体菜用枸杞新品种天精3号选育[J]. 核农学报,2013,27(6):723- 730.
[12]Dhooge E,Van Laere K,Eeckhautt,et al. Mitotic chrimosome doubling of plant tissues in vitro[J]. Plant Cell Tissue and Organ Culture,2011,104(3):359-373.
[13]白英豪,张晓丽,李明军. 秋水仙素诱导丹参多倍体研究进展[J]. 江苏农业科学,2018,46(5):18-20.
[14]殷婷婷. 油菜多倍体诱导技术[J]. 江西农业,2018(10):14-16.
[15]彭绿春,陶俊锋,段修安,等. 腋花杜鹃多倍体诱导和鉴定[J]. 核农学报,2018,32(2):257-265.
[16]孙嘉曼,卢 江,韦绍龙,等. 不同浓度秋水仙素对二倍体野生蕉的多倍体诱导[J]. 西南农业学报,2017,30(9):2097-2100.
[17]孔维一,甘 颖,陈洪伟,等. 一串红体细胞多倍体诱导及其子代表型研究[J]. 北京农学院学报,2018,33(2):87-91.
[18]吳照云,陈发波,姚启伦,等. 涪陵胭脂萝卜多倍体诱导研究[J]. 种子,2017,36(3):38-41.
[19]邓 云,张蜀宁,孙敏红,等. 采用秋水仙碱创制优质、抗热同源四倍体不结球白菜[J]. 武汉植物学研究,2006,24(2):159-162.
[20]Abdoli M,Moieni A,Badi H N. Morphological,physiological,cytological and phytochemical studies in diploid and colchicine-induced tetraploid plants of Echinacea purpurea (L.)[J]. Acta Physiologiae Plantarm,2013,35(7):2075-2083.
[21]漆慧娟. 甜叶菊多倍体诱导及其生物学特性的研究[D]. 杭州:浙江农林大学,2014.
[22]钟 程,田 鑫,刘伦沛,等. 秋水仙素诱导贵州野生淡黄花百合的多倍体[J]. 贵州农业科学,2015,43(8):9-11.
[23]吴 顺,葛倩雯,刘肖坷,等. 紫花丹参多倍体的初步诱导[J]. 中药材,2016,39(3):479-481.