不同玉米芯基质预处理方式对猴头菌生长的影响
2020-09-01范博文张俊莹赵长江李佐同
范博文,张俊莹,赵长江,李佐同
(黑龙江八一农垦大学农学院,大庆 163319)
猴头菌(Hericium erinaceus)是我国珍贵的食药兼用真菌[1],属于猴菇目、猴菇科、猴菇属[2]。猴头菌味道鲜美,营养丰富,有“山珍猴头、海味燕窝”之称。在猴头菌的栽培过程中,木屑为常规的栽培基质。但随着生产规模扩大,以及人们对森林资源的重视。木屑资源紧张已成为限制食用菌产业发展的一大因素[3]。用玉米芯代替木屑栽培猴头菌,一方面可以缓解木屑资源紧张问题,另一方面可以有效的利用农业废弃物。内在菌丝胞外酶活性和外在基质营养组成是决定猴头菌分解能力及产量的两个重要因素[4-5]。赵占国[6]认为葡萄糖作为营养源最适合猴头菌菌丝生长。韩春华[7]研究发现培养基质中氮源的添加比例对食用菌产量具有一定的影响。并且氮源的添加比例可以改变食用菌子实体品质[8]。King 等[9]现在胞外酶系统中,纤维素酶、淀粉酶、木质素酶对分解基质起到重要作用。Turner,Claydon 等[10-11]在研究中发现,食用菌分解胞外酶活性和产量具有相关性。李永江等[12]发现,食用菌大多数重要的农艺性状如菌丝生长速度、酶活性、质量以及产量等都是数量性状,是由多个数量基因座位和环境控制。目前,对猴头菌代替木屑栽培的研究主要集中于基质添加种类和比例,而对代料基质预处理方式的研究较少。通过对玉米芯基质进行预处理,探究不同预处理方式对猴头菌生长的影响,为农业废弃物栽培猴头菌提供理论依据和技术指导。
1 材料与方法
1.1 试验材料
猴头菌(Hericium erinaceus)菌种为“RT22”。
1.2 试验方法
1.2.1 母种制备
将马铃薯去皮称200 g,切成1 cm3的正方体后。加入800 mL 蒸馏水中,煮至沸腾后保持30 min。用纱布将马铃薯滤汁滤出,加入20 g 葡萄糖和20 g 琼脂,定容至1 000 mL。装入试管中高压灭菌,试管灭菌后制成斜面培养基,接种猴头菌菌种。
1.2.2 原种制备
原种配方为:木屑80%、麦麸18%、石膏1%、蔗糖1%,物料含水量65%,原种配料装袋后高温灭菌,灭菌后接种猴头菌母钟。
1.2.3 玉米芯基质栽培种制备
玉米芯栽培种配方为:玉米芯80%、麦麸18%、石膏1%、蔗糖1%,物料含水量65%。按四个处理(表1)称取物料,装袋后高温灭菌,灭菌后接种猴头菌原种,接种后将菌棒放入相对湿度60%、温度25 ℃环境中培养,当菌丝长满菌袋后作为玉米芯基质栽培种。
1.2.4 出菇期管理
将猴头菌玉米芯基质栽培种放入相对湿度90%、温度18 ℃的环境中培养,每天早晚通风2 次,一次通风0.5 h。当猴头菌子实体菌刺约0.5 cm 时,采收子实体。

表1 玉米芯基质预处理方法Table1 Pretreatment of maize cob
1.3 指标测定
1.3.1 猴头菌生长指标测定
猴头菌菌棒长满袋后测定长满袋天数和菌丝生长速度指标;采摘猴头菌子实体后,测定子实体干重和生物学效率指标。
1.3.2 玉米废弃物培养料碳氮养分及胞外酶活性测定
玉米芯基质的碳氮比需分别测定玉米芯基质的总碳和全氮含量,总碳(TC)利用非分散型红外线气体分析仪测定[13];全氮采用凯氏定氮法[14]测定。羧甲基纤维素酶、β-葡萄糖苷酶、半纤维素酶、淀粉酶、滤纸纤维素酶、过氧化物酶、漆酶活力利用紫外可见分光光度计(日立)U3900 测定[15-19],羧甲基纤维素酶、β-葡萄糖苷酶、半纤维素酶在540 nm 波长处测定光密度值;淀粉酶在520 nm 波长处测定光密度值;滤纸纤维素酶在540 nm 波长处测定光密度值;漆酶在600 nm 波长处测定光密度值。
1.4 数据分析
通过Excel 2016 对数据进行整理,GraphPad Prism 5 对数据进行分析,文中数据均有3 次或3 次以上重复。
2 结果与分析
2.1 不同预处理方式对形态指标的影响
2.1.1 不同预处理方式对菌丝生长速率的影响
菌丝生长速度反应菌丝生长快慢及长势强弱。如图1 所示,在不同基质预处理中,B 处理的菌丝生长速度显著高于其他处理(P<0.05),与S 处理、F 处理、T 处理相比分别提高13.31%、9.58%、7.33%;T 处理的菌丝生长速度与F 处理差异不显著(P<0.05);F处理的菌丝生长速度与S 处理差异不显著(P<0.05);T 处理的菌丝生长速度显著高于S 处理(P<0.05),提高了5.57%。

图1 不同预处理方式对菌丝生长速率的影响Fig.1 Effect of different pretreatment methods on mycelium growth rate
2.1.2 不同预处理方式对菌丝长满袋天数的影响
菌丝长满袋天数反应菌丝长满培养基质的时间。如图2 所示,在不同基质预处理中,S 处理的菌袋长满袋天数为45.33 d,显著高于T 处理和B 处理(P<0.05),分别提高了6.25%、13.96%;S 处理的菌袋长满袋天数与F 处理差异不显著(P<0.05);F 处理的菌袋长满袋天数为43.56 d,与T 处理、B 处理差异不显著(P<0.05);T 处理的菌袋长满袋天数为42.67 d,与B 处理差异不显著(P<0.05)。

图2 不同预处理方式对菌丝长满袋天数的影响Fig.2 The effect of different pretreatment methods on the days of mycelium filling
2.1.3 不同预处理方式对子实体干重的影响
干重反应样品干物质重量。如图3 所示,在不同基质预处理中,F 处理和B 处理的子实体干重分别为21.96 g 和21.86 g,其中F 处理显著高于S 处理(P<0.05),提高19.68%;B 处理显著高于S 处理(P<0.05),提高19.15%;T 处理的子实体干重为19.14 g,与F 处理、B 处理相比差异不显著(P<0.05);S 处理的子实体干重为18.35 g,与T 处理相比差异不显著(P<0.05)。
2.1.4 不同预处理方式对子实体生物学效率的影响
生物学效率反应样品的转化率。如图4 所示,在不同基质预处理中,B 处理的生物学效率为0.51,显著高于T 处理(P<0.05),提高38.92%;F 处理的生物学效率为0.51,显著高于T 处理(P<0.05),提高39.52%;S 处理的生物学效率为0.42,与B 处理、F 处理差异不显著(P<0.05);T 处理的生物学效率为0.37,与S 处理差异不显著(P<0.05)。

图3 不同预处理方式对子实体干重的影响Fig.3 Effect of different pretreatment methods on dry weight of sub body
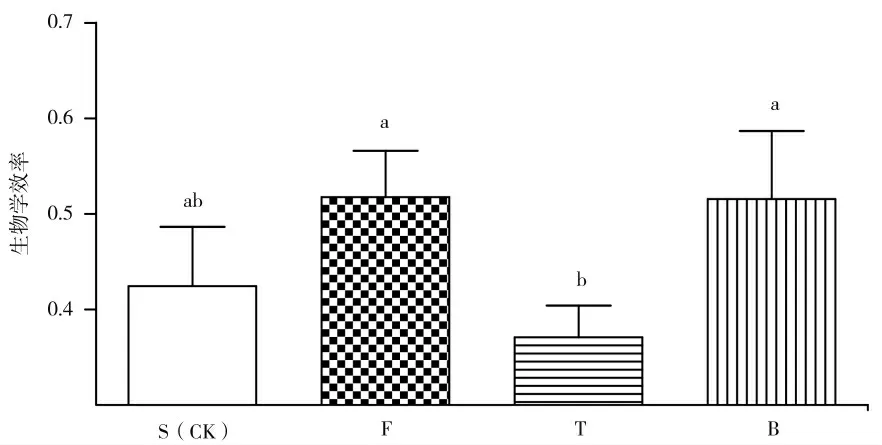
图4 不同预处理方式对生物学效率的影响Fig.4 Effects of different pretreatment methods on biological efficiency
2.2 不同预处理方式对胞外酶活性的影响
2.2.1 不同预处理方式对纤维酶活性的影响
纤维素酶表征了纤维素酶的活力。如图5 所示,不同玉米芯基质预处理中,猴头菌菌丝分泌羧甲基纤维素酶影响较大。其中,B 处理的羧甲基纤维素酶最高,显著高于S 处理(P<0.05),提高了44.2%,与F处理和T 处理差异不显著(P<0.05);F 处理与T 处理、S 处理差异不显著(P<0.05);T 处理和S 处理之间的差异并不显著(P<0.05)。不同基质预处理对滤纸纤维素酶有较大的影响。其中S 处理的滤纸纤维素酶最高,显著高于T 处理(P<0.05),提高了21.88%,与F 处理和B 处理差异不显著(P<0.05);F 处理与T 处理、B 处理差异不显著(P<0.05);T 处理和B 处理之间差异不显著(P<0.05)。不同基质预处理对猴头菌菌丝分泌β-葡萄糖苷酶影响较大。其中,T 处理的β-葡萄糖苷酶最高,显著高于F 处理(P<0.05),提高了33.42%,与S 处理和B 处理差异不显著(P<0.05);B处理与S 处理、F 处理差异不显著(P<0.05);S 处理和F 处理之间差异并不显著(P<0.05)。
2.2.2 不同预处理方式对淀粉酶和半纤维素酶活性的影响
淀粉酶是指一类能催化分解淀粉(包括糖原、糊精等)的糖苷键的酶之总称;半纤维素酶是能将构成植物细胞膜的多糖类(纤维素和果胶物质除外)加水分解的各种酶总称。如图6-A 所示,不同基质预处理对猴头菌菌丝分泌淀粉酶的影响较大。T 处理的淀粉酶含量0.14,F 处理的淀粉酶含量0.12。其中T 处理的淀粉酶含量最高,显著高于S 处理(P<0.05),相比提高了40%;S 处理的淀粉酶含量最低,但与F 处理和B处理相比差异不显著(P<0.05);F 处理和B 处理之间相比差异不显著(P<0.05)。如图6-B 所示,不同基质预处理对猴头菌菌丝分泌半纤维酶的影响较大。其中F 处理的半纤维酶含量为3.23,B 处理的半纤维酶含量为3.2,F 处理的半纤维酶含量显著高于T 处理(P<0.05);B 处理的半纤维酶含量显著高于T 处理(P<0.05);F 处理和B 处理之间相比差异不显著(P<0.05)。

图5 不同预处理方式对纤维素酶活性的影响Fig.5 Effect of treatments on cellulase activity of H.erinaceus
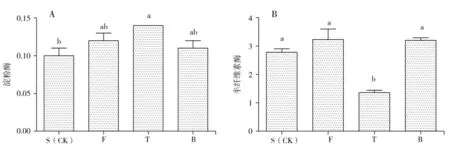
图6 不同预处理方式对淀粉酶(A)和半纤维素酶(B)活性的影响Fig.6 Effect of different pretreatment methods on the activity of amylase and hemicellulase
2.2.3 不同预处理方式对木质素酶活性的影响
木质素酶对木质素具有很强的降解能力。如图7-A 所示,不同基质预处理对猴头菌菌丝分泌过氧化物酶的影响较大。F 处理的过氧化物酶含量为0.14,B 处理的过氧化物酶含量为0.12,T 处理的过氧化物酶含量为0.09。F 处理的过氧化物酶含量最高,显著高于S 处理和T 处理(P<0.05),分别提高了86.67%和33.34%;S 处理和T 处理之间的过氧化物酶含量差异不显著;B 处理和F 处理的过氧化物酶含量差异不显著。如图7-B 所示,不同基质预处理对猴头菌菌丝分泌漆酶的影响较大。B 处理的漆酶含量为18.91,F 处理的漆酶含量为17.87,S 处理的漆酶含量为17.50。B 处理的漆酶含量最高,显著高于S 处理和T 处理(P<0.05),分别提高了8.05%和12.63%;S 处理和T 处理之间的漆酶含量差异不显著;B 处理和F处理的漆酶含量差异不显著。

图7 不同预处理方式对木质素酶活性的影响Fig.7 Effects of different pretreatment methods on the activity of lignin enzyme
2.2.4 胞外酶活性与菌丝生长速度及子实体干重相关性分析
通过相关性分析将七种胞外酶活性同猴头菌菌丝生长速度和子实体干重进行分析(表2),菌丝生长速度与羧甲基纤维素酶、β-葡萄糖苷酶、半纤维素酶、淀粉酶、过氧化物酶、漆酶呈正相关,与滤纸纤维素酶呈负相关。其中,菌丝生长速度与羧甲基纤维素酶的相关度最高,为0.858;与漆酶的活性次之,为0.675;与过氧化物酶的活性为0.418。子实体干重与羧甲基纤维素酶、滤纸纤维素酶、半纤维素酶、淀粉酶、过氧化物酶、漆酶呈正相关,与β-葡萄糖苷酶呈负相关。其中,子实体干重与过氧化物酶相关度最高,为0.965;与羧甲基纤维素酶的活性次之,为0.908;与漆酶的活性为0.733。七种胞外酶与菌丝生长速度没有显著的相关性,但是七种胞外酶中的过氧化物酶与子实体干重显著相关。在菌丝生长速度和子实体干重中,过氧化物酶、漆酶和羧甲基纤维素酶的相关性较高。说明过氧化物酶、漆酶和羧甲基纤维素酶对猴头菌菌丝生长速度和子实体干重影响较大。在三种纤维素酶中,菌丝生长速度和子实体干重并不都与其呈正相关,说明猴头菌生长是通过三种纤维素酶相互调节来影响的。

表2 胞外酶与菌丝生长速度及子实体干重之间的相关性Table 2 Correlation between extracellular enzyme and mycelial growth rate and dry weight of fruiting body
3 讨论
食用菌通过从基质中吸收的营养来用于自身的生长发育。对栽培基质进行预处理,有利于菌丝对营养物质的吸收,更好地提高子实体的产量和品质。以往对玉米芯基质代替木屑栽培食用菌研究主要集中于添加玉米芯的比例,研究直接对玉米芯材料本身进行处理,以提高玉米芯基质的栽培效率。通过试验发现,不同玉米芯预处理方法对猴头菌菌丝生长速度、长满袋天数、干重和生物学效率都有较大影响。预处理改变了玉米芯基质的理化性质,将玉米芯中大分子的营养物质变成各种小分子的营养物质,更有利于猴头菌吸收和利用。T 处理和S 处理的原理都是对玉米芯进行预湿,使其吸收水分后变得松软,但二者的程度又不同。F 处理和B 处理是通过微生物对玉米芯进行一定的降解,但二者微生物的来源又不同。郭亚萍等[20]研究发现,双孢蘑菇堆肥过程优势类群为芽孢杆菌属(Bacillus)、类芽孢杆菌属(Paenibacillus)。猜测直接发酵处理和菌剂发酵处理过程中,这两个菌属也起到了主要的作用。在四种不同方法处理玉米芯基质中,都能检测到纤维素酶、半纤维素酶、淀粉酶和木质素酶。在七种酶中,不同处理方法的酶活性差异性较大。但纤维素酶活性一直处于较高水平,猜测与纤维素酶活性相关的cel1、cel2、cel3基因起到了关键的作用[21]。玉米芯基质中小分子营养种类和含量变化,也会对酶活性有一定的影响。有研究表明用碱性物质预处理稻草后将其作为栽培食用菌的基质,发现食用菌对稻草的分解能力得到提升[22]。用微生物预处理小麦秸秆后,小麦秸秆的还原糖含量发生了变化[23]。
通过相关性分析将七种胞外酶活性同猴头菌菌丝生长速度和子实体干重进行分析,菌丝生长速度与羧甲基纤维素酶、β-葡萄糖苷酶、过氧化物酶、漆酶的活性相关性较高;子实体干重与羧甲基纤维素酶、淀粉酶、过氧化物酶、漆酶的活性相关性较高。在猴头菌在营养生长时期和生殖生长时期,虽然胞外酶含量发生了变化,但羧甲基纤维素酶、过氧化物酶和漆酶的相关性一直都很高。因此,在兼顾子实体干重、菌丝生长速度的同时,提出羧甲基纤维素酶、过氧化物酶、漆酶可以用于连锁分析培养料的养分状况与食用菌生长状况。
从碳氮比的角度分析(如表3),进过预处理后玉米芯基质的碳氮比有较大的差异。碳源营养和氮源营养比例对猴头菌菌丝和子实体生长起决定性作用。碳氮比较高,子实体形成困难;碳氮比较低,子实体原基生长困难。在毛木耳(Auricularia polytricha)的研究中发现,碳氮比较低时,随着碳氮比升高,毛木耳干重也会增加。但到达一定值后,随着碳氮比升高,毛木耳干物质干重反而降低[24]。秀珍菇(Pleurotus geesteranus)的菌丝和子实体对碳氮比环境的要求较宽泛,当碳氮比在10~20∶1 时,秀珍菇菌丝生长最适合;当碳氮比在39.21~59.32∶1 时,秀珍菇子实体生长最适合[25-26]。在试验中,处理S 和处理T 的碳氮相对较高,处理F 和处理B 的碳氮比相对较低。猜测微生物预处理可以降低基质的碳氮比。碳氮比相对较低的处理F 和处理B,产量和菌丝生长速度都较高。因此,当玉米芯基质碳氮比在32~41∶1 时,更适合猴头菌的生长。

表3 不同玉米芯基质预处理后的碳氮比Table 3 Effect of maize cob on carbon and nitrogen ratio
4 结论
在供试的四种不同的玉米芯基质预处理中,菌剂发酵处理可以提高菌丝生长速度、子实体干重、生物学效率;缩短菌丝长满袋天数。在七种胞外酶中,羧甲基纤维素酶、滤纸纤维素酶、β-葡萄糖苷酶的活性、半纤维素酶、漆酶的活性较高;淀粉酶、过氧化物酶的活性较低。通过相关性分析,过氧化物酶、羧甲基纤维素酶和漆酶三种胞外酶与菌丝生长速度及猴头菌产量相关性较大。子实体产量最高的处理玉米芯基质碳氮比在32~41∶1 之间。
