植物-害螨-捕食螨中寄主植物表面毛和挥发物对捕食螨行为的影响
2020-08-26张燕南刘林鑫朱利君杨丽红
张燕南, 刘林鑫, 朱利君, 杨丽红*
(1.绵阳师范学院生命科学与技术学院,四川绵阳 621006;2.北川羌族自治县自然资源局, 四川绵阳 621006)
0 引言
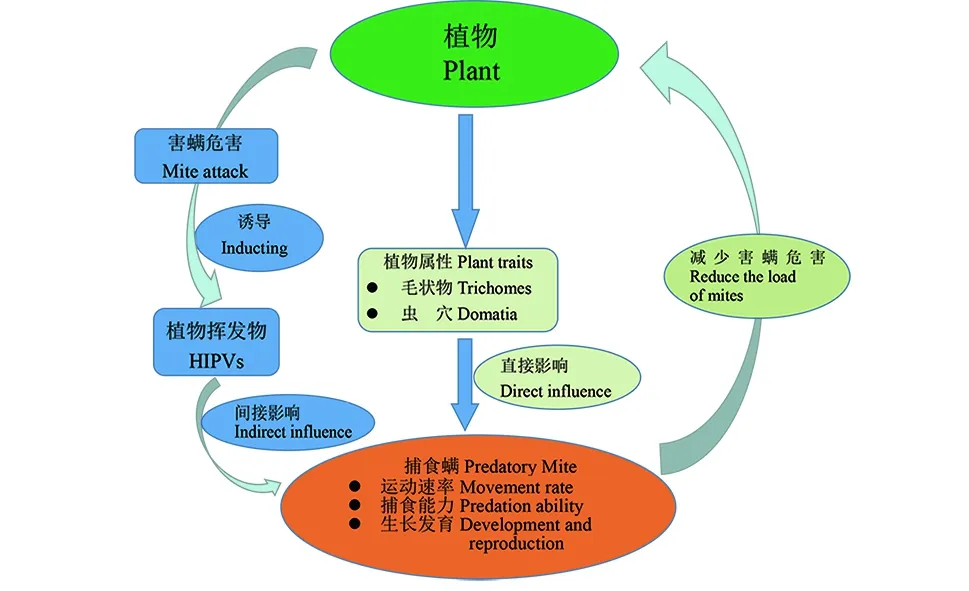
图1 寄主植物对捕食螨的直接影响和间接影响Fig.1 The indirect and direct effect of host plants on predatory mites (Acari: Phytoseiidae)
由于大田作物的多样性和复杂性,对于大田作物的生物防治具有一定的挑战性.在一些作物上生物防治似乎很容易实现对害虫的控制,而对于其它一些作物天敌对害虫的控制又显示的特别困难.这些自然天敌的效能在一定程度上取决于寄主植物防御性.植物的防御特性主要分为直接防御(又称为“constitutive defense”)和间接防御(又称为“inducible defense”)[1].直接防御中的物理防御包括一些物质的分泌[毒素(toxins)、消化抑制剂(digestion inhibitors)和驱虫剂(repellents)]和形态特征[体刺(spines)、蜡状物(waxes)、毛状物(trichomes)和虫穴(domatia )][2].钦俊德的专著详细论述了昆虫与植物的相互协同进化关系,其指出物种在漫长的生物进化过程中,因营养、繁殖、防卫、扩散等需要,“寄主植物-害螨-捕食性天敌”三者之间便相互选择、相互适应,从而形成了直接或间接的复杂互作关系[3].在三级营养生态系统中,一方面,植物直接或间接地为天敌提供食物、猎物和庇护所等资源,同时植物在受到昆虫危害后将会产生挥发性化合物来吸引天敌从而使植物产生直接的防御反应以抑制害螨种群[4-5].第二方面,作为化学信号传介媒体的植食性昆虫,其利用植物危害后散发的挥发性物(HIPVs)可以准确地找到寄主植物进行取食、产卵及繁殖等行为.以上两个过程对三营养关系进行有效的调控,构成一个平衡的生态系统[6-7](如图1).
近三十年来,许多学者已经总结出植物的毛状体和虫穴影响节肢动物的行为和性能[8],同时,众多的实验室也在三级营养关系相互作用的背景下对HIPV进行了研究,大多数学者普遍认识到HIPV的生态重要性,因为HIPV能够吸引捕食性天敌和寄生性天敌来防止害虫对植物攻击.然而,植物表面特性对捕食螨的影响仍存在很大争议,同时因捕食螨体型小且不能飞等特性,很多学者对HIPV在寄主植物-害螨-捕食螨三级营养关系大的生态背景下对捕食螨的影响也存在质疑.本综述重点描述寄主植物表面毛和HIPV在三级营养关系中对捕食螨具体生物学影响,在简要概述历史性问题和主要研究问题的基础上最终能利用植物表面毛和挥发物对害螨生物防治提供一些新的线索.
1 寄主植物表面毛对捕食螨直接影响
植物叶片表面有各种各样的结构(如毛状体和虫穴)增加了节肢动物所经历的微环境的复杂性.毛状体类似于毛发状结构,其密度、长度和类型取决于植物,更具体地说取决于它们产生的器官.毛状体包括很多不同类型,例如:非腺毛、腺毛、卷曲毛、直毛、单细胞毛、多细胞毛、钩毛和单细胞毛[9].在生物防治中,毛状体通过对害虫和天敌提供物理防御来承担生物屏障的角色[10].虫穴是植物表面体上的一种小型形态发生中空结构,可作为螨虫和其他节肢动物的庇护所.作为回报,生活在虫穴中的螨虫和其他节肢动物通过清除致病真菌的孢子和菌丝和其他有害节肢动物为植物提供了好处[1].植物表面因毛状物和虫穴的存在不但影响害螨行为影响,同时直接影响捕食螨行为,也为大田生物防治提供了一个防治方向.
1.1 寄主植物表面毛对捕食螨运动速率的影响
寄主植物表面特征可以阻碍或促进捕食者的运动,从而显著影响与宿主或猎物的接触率.有研究表明在高密度毛状物环境下,捕食螨运动速率受到毛状物负面影响.有研究对比了不同密度毛状物对瑞氏钝绥螨(Amblyseiusswirskii)运动速率,表明在塑料板上运动速率最高,玫瑰次之,而在菊花、非洲菊和番茄上运动速率没有差异[11].智力小智绥螨(Phytoseiuluspersimilis)在不同寄主植物表面结构上的运动速率也得到了相同验证[12].有研究表明,捕食螨在低密度毛状物的植物上的行走速度为0.10 ± 0.030 cm·s-1, 而捕食螨在表面毛状物密度不多的玫瑰上的行走速度为0.12 ± 0.015 cm·s-1[13],以上两个结果表明,寄主植物表面毛密度只有到达一定密度之后才能对捕食螨行为起负面作用.此外,毛状物的长短、软硬等均能影响捕食螨在植物表面的爬行速度[14].以上这点为大田释放捕食螨提供新的策略,要求在释放捕食螨时应考虑释放植物表面毛的特性从而增加对捕食螨的利用率.
1.2 寄主植物表面毛对捕食螨捕食能力和生长发育的影响
植物形态特征可以影响不同捕食者的生存、繁殖力、觅食和它们与猎物的相互作用以及对猎物的功能反应[15-18].捕食螨运动速率会受到植物表面毛密度的负面影响,这也就意味着增加了捕食螨寻找猎物时间以及降低捕食率,这一观点在胡瓜钝绥螨(Neoseiuluscucumeris)和智力小智绥螨(P.persimilis)上均有证实[12,18-19].
然而,也有研究表明,植物表面毛对捕食螨捕食能力能起到积极作用,一些螨类喜欢具有致密毛状物的植物种类[8, 12].瑞氏钝绥螨(A.swirskii)喜好植物表面具有簇状虫穴作为产卵点,这导致了植物虫穴和每片叶片的卵、幼螨和成螨数量成相关[20-21]. 这种现象可能是由于毛状物多的植物表面湿度比较大,而捕食螨产卵在湿度低的地点其成活率明显很低[13]而导致得.此外,表面毛少的植物不利于捕食螨附着,很容易从叶片上掉落下来.相反,植物表面毛多的植物会减少对捕食螨的这种负面影响,会增加捕食螨觅食成功率[12, 22].
捕食功能于1959年被Holling用来评价捕食性天敌的捕食能力的重要研究手段,其描述捕食性天敌对猎物捕食的捕食量随着猎物密度变化而变化,在研究捕食者-猎物二者系统分析中具有非常重要的参考价值[23].捕食功能反应受到很多非生物因素的影响,如:温度、湿度等,同时也受到很多生物因素的影响,如:猎物类型、捕食者雌雄、寄主植物种类等[24-26].Podisusnigrispinus在辣椒和茄子上对甜菜夜蛾(Spodopteraexigua)的捕食类型属于Holling II型,而在番茄上的捕食类型属于Holling III[27]. 14斑点瓢虫分别在落芒草(Ricegrass)和小麦草(Wheatgrass)对俄罗斯小麦蚜虫的捕食类型分别为Holling II型和Holling III型[28].但植绥螨科的捕食类型基本均属于Holling II型[29,17].
此外,毛状物对捕食螨生长发育也起到积极作用.在自然毛状物密度较少的葡萄品种上人为增加簇状的聚酯纤维,捕食螨的生殖率会明显增加[30].加州新小绥螨(N.californicus)喜好在虫穴多的地点产卵[31].其它研究也证明,植物表面毛状物增加以后能增加捕食螨的分布和产卵量[16, 24,32].研究者利用毛状物对捕食螨生长发育的影响提出了“植物银行”的策略.研究发现加州新小绥螨(N.californicus)在羊毛状的人造毛状物上产卵量极高[33].加州新小绥螨(N.californicus)喜欢在自然状态的毛状物及所选5种人造毛状物(羊毛,蚕丝,尼龙,纤维胶,聚酯)上产卵[22].毛状物对捕食螨生长发育影响的特点为我们在田间释放捕食螨提供了新的方向.在大田,对于捕食螨不喜好的植物毛状物(比如玫瑰),我们就可增加“植物银行”来克服这一困难,从而加大对捕食螨的利用率.
2 植物挥发物在三级营养关系中对捕食螨的影响
植物、害虫和天敌之间的三级关系相互作用是所有陆地生态系统的组成部分.害虫诱导的植物挥发物(HIPVs)在这些相互作用中起着关键作用,因为它们可以吸引捕食者和寄生性天敌到达害虫危害的植物,从而降低害虫进一步危害[34-35].对于专一性天敌来说,这种能力也许是与生俱来的,但对于广谱性的天敌必须学会区分不同的HIPV混合物,帮助他们专注于最有利可图的线索[34].害虫诱导植物产生的挥发物(HIPVs)对天敌行为的影响已经在很多系统中均有记录,比如对异色瓢虫(Harmoniaaxyridis)和精美蚁形金小蜂(Theocolaxelegans)[36-37]等天敌均有研究.植物挥发物对捕食螨在不同作物、害螨系统中很早就有研究[38,39].植物-害螨系统中8个科的11种植物,叶螨科和瘿螨科的4种害螨和9种捕食螨之间有相互关系[40].在此基础上我们补充了近三十年以来其它害螨诱导植物释放挥发物对捕食螨行为影响的研究(表1).这些研究表明,被害螨危害的植物释放挥发物(HIPVs)能明显对捕食螨具有引诱作用.同时,已有研究表明,害螨诱导植物释放挥发物对捕食螨的吸引并不是对害螨诱导的单个植物挥发物的吸引的结果,它们可以在气味混合物中识别这些化合物[41].
随着HIPVs对捕食螨行为的影响,近年来,各位学者对植物组织释放单一化合物 (VOCs)对捕食螨行为影响产生了大量兴趣.已有研究证明,除植物被害虫危害后产生挥发物对捕食螨具有引诱作用外,机械损伤的植物释放出的化合物对捕食性天敌也具有引诱作用[42-43].害螨诱导的植物挥发性有机化合物是一个复杂的化学-感觉信息包,很难预测捕食螨是被吸引还是被排斥[44-45].然而,在气相色谱-质谱联用(GC/MS)色谱图的帮助下,我们可以分析特定的化学成分,并能检测出每种化合物对捕食螨行为的影响.在受二斑叶螨危害诱导的大豆叶的挥发性混合物中,四种挥发性化合物(芳樟醇、水杨酸甲酯、(E)-b-辛烯和(E)-4,8-二甲基-1,3,7-壬三烯)对智力小智绥螨(P.persimilis)具有吸引作用[46].水杨酸甲酯和芳樟醇能够吸引加州新小绥螨(N.californicus),而健康植株释放的三种化合物:(Z)-3-己烯-1-醇、(Z)-3-己烯乙酸酯和(E)-2-己烯醛均在二斑叶螨危害及机械损伤危害后释放化合物中也有发现,且对捕食螨也具有吸引作用[43].同时,此项研究还显示,水杨酸甲酯对捕食螨具有很强的吸引作用,它的吸引能力和植物被二斑叶螨危害后释放化合物对捕食螨吸引能力等同.研究表明,被神泽氏叶螨(Tetranychuskanzawai)危害的茶树叶片释放的挥发物:(E)-β-罗勒烯,4,8-二甲基-1,3,7-nonatriene (DMNT)和(E,E)-α-farnesene对捕食螨-Neoseiuluswomersleyi均具有很好的引诱作用.同样的,大豆叶片被二斑叶螨危害后,诱导产生化合物:DMNT, 水杨酸甲酯, β-石竹烯和 (E,E)-4,8,12-三甲基-1,3,7,11-tridecatetraene[47].己醛、壬醛、乙酸辛酯、庚醛和月桂烯等化合物明显对巴氏新小绥螨(N.barkeri)具有引诱作用.以上这些研究都显示捕食螨虽体型小,不易飞翔但对一些化合物很敏感,可利用其作为觅食、定位的重要信息线索[48].
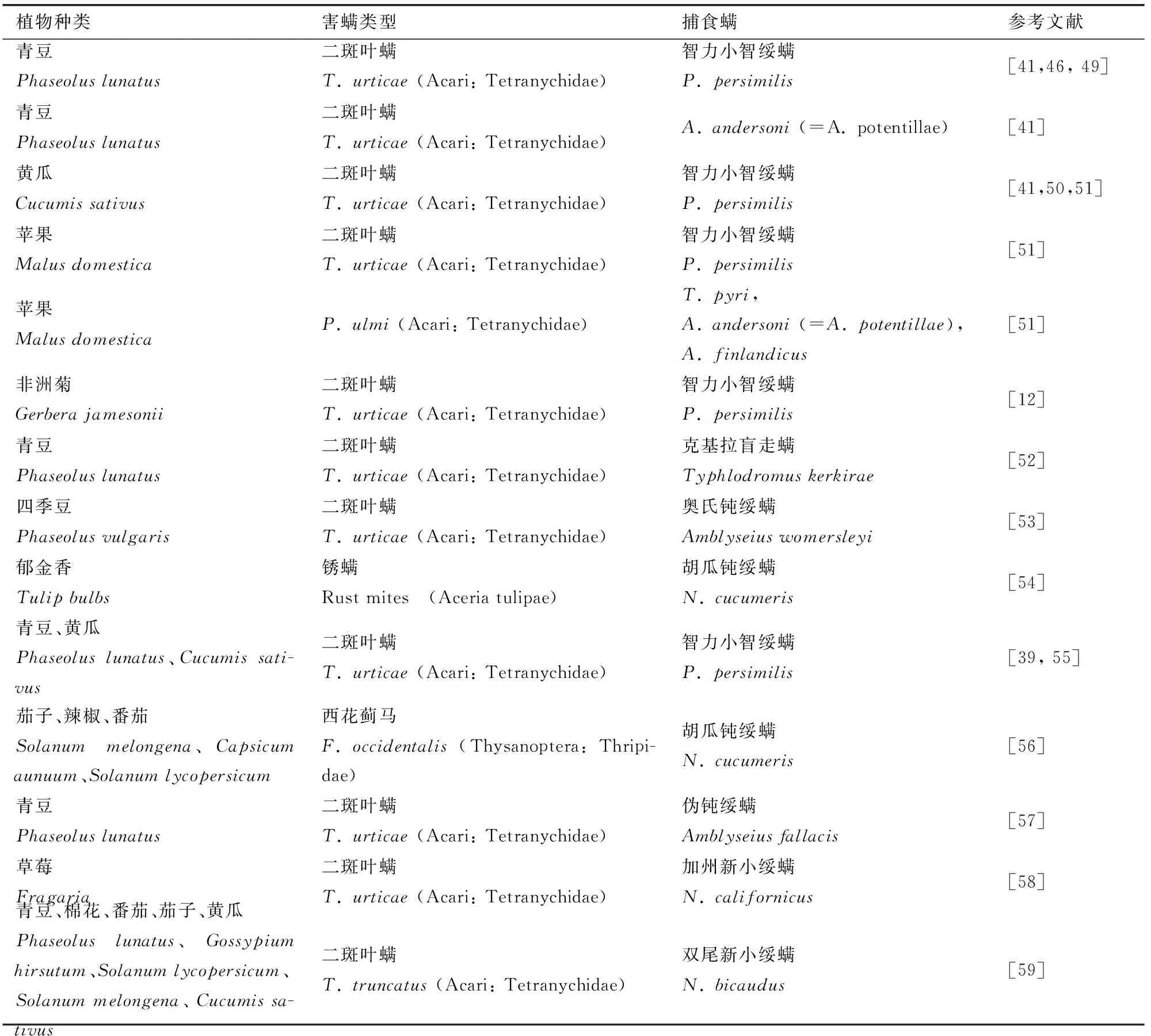
表1 三级营养关系中植物被害螨危害后释放挥发物对捕食螨吸引作用研究Tab.1 Tritrophic systems for which evidence for carnivore attraction by herbivore-infested plants has been demonstrated
3 总结与展望
以上研究均表明寄主植物挥发物和表面毛均能影响捕食螨行为,因此,释放策略必须将这些属性视为生物控制实践中必不可少的和相互作用的组成部分.根据本文介绍,我们认为有必要将植物属性作为生物控制实践的一个基本和相互作用的组成部分来考虑.首先,增加植物毛状物对捕食螨起积极或者负面影响.一方面,植物毛状体可以降低捕食螨和害虫附着在植物表面的能力,同时影响捕食螨爬行速率从而降低捕食者的效率.另一方面,寄主植物毛状物又可作为捕食螨的产卵地点,同时可作为捕食螨避难所而免受其它微生物危害.因此,寄主植物毛状物对捕食螨影响具有复杂性,毛状物长度、类型及硬度均是考虑因素,因此建立毛状物长度和捕食螨爬行速率之间模型关系可为捕食螨生物防治提供很好的理论基础.其次,植物挥发物对捕食螨行为具有导向作用,从复杂挥发物中筛选出对捕食螨具有明显引诱作用的单一化合物是现如今生态学上的一个热点和难点.在理论层面,已有大量学者研究出对固定捕食螨具有引诱作用的化合物,但在大田实际应用中化合物对捕食螨引诱仍是空白.导致这一现象主要因为捕食螨体型小,不具飞行能力,这就为研究挥发物对捕食螨引诱带来困惑和困难.此外,单一在实验室研究的单一化合物一般价格昂贵,不易收获,这就为大田实际应用方面带来很大困难.研究出操作简单,价格实惠的单一化合物将是新的研究方向.同时,HIPVs在该领域的应用的研究通常集中在它们吸引天敌的能力上,但研究HIPVs对害螨引诱可增加捕食螨对害螨集中捕食效果,这也为捕食螨大田释放提供一个方向.
总之,我们应该充分利用植物对捕食性螨的影响(直接和间接),加强大田捕食性螨的释放策略.其它的策略,如“植物银行”和使用补充食物来源为进一步利用植物对捕食螨的影响提供了创新的机会,最终提高捕食螨的成本效益.由于许多作物在其发育的所有阶段不能保证随时提供植物资源和产卵地点,因此利用植物提供产卵地和植物资源来克服这些问题是一个好的选择.
