长江远端三角洲表层沉积物中底栖有孔虫分布及其对环境的指示
2020-08-26张静胡刚马国荣惠建国张明震张勇杜宝霞
张静,胡刚,马国荣,惠建国,张明震,张勇,杜宝霞
1.兰州大学地质科学与矿产资源学院,兰州 730000
2.青岛海洋地质研究所,青岛 266071
3.中国科学院西北生态环境资源研究院/甘肃省油气资源研究重点实验室,兰州 730000
长江远端三角洲是东海内陆架泥质沉积体的重要组成部分,位于台湾海峡北部出口处,黑潮支流、台湾暖流和闽浙沿岸流等不同性质和来源的水系汇集,强烈的季风交替以及复杂的地形等条件加剧了近岸海域的水文变化[1],使潮汐、上升流、锋面等现象聚集[2]。因此,了解和掌握该海域的水团分布及其变化,不仅对不同物质来源的沉积物输运和沉积分布具有重要影响,而且对深刻认识东海沿岸水文环境状况、合理开发及保护福建沿海海洋资源等方面都具有重要意义[3]。
关于东海水文环境变化的研究方法除了通过传统的水文理化性质(盐度、温度、溶解氧)数据分析外,近年来随着技术的发展,一些研究通过对沉积物本身及其中的稀土元素、重金属和石英等矿物指标进行分析讨论[4-6],以及利用生物指标对水文环境进行间接指示,如硅藻、浮游动物、有孔虫等[7-9]。海洋生物对海洋水体环境敏感,底栖有孔虫作为一种常见的海洋微体生物,由于其群落组合及分布主要受控于底层水体环境,因此可为研究海域水文特征和海洋沉积环境的演化提供可靠证据[10-11]。特别是对于沿岸浅海环境来说,有孔虫的属种特征、数量分布,群落组合能够反映不同的水文环境,对水团分布、海流状况等均有指示意义[12]。过去我国对东海海域及其近海进行了大量的地质调查研究,其中针对浙江沿岸、台湾海峡等地区的海域表层沉积物有孔虫调查,为第四纪海洋研究提供了大量的古生物证据[13-14]。以往的有孔虫研究虽然涉及过本文研究区,但主要位于台湾海峡中南区域和长江口附近,而长江远端三角洲及其沿岸的研究工作较薄弱,零星研究也仅限于对台湾海峡平潭岛附近小区域表层沉积物有孔虫分析与水团环境研究[15]。显然,目前的研究资料难以全面反映长江远端三角洲海域不同水团展布的复杂水文信息。因此,本文以1∶25万比例尺的海洋地质调查精度,对取自福建霞浦县20 km2海域分布的145件表层沉积物样品进行了底栖有孔虫定量分析,探讨了福建近岸底栖有孔虫组合的分布规律及其与水团分布之间的对应关系。
1 区域水文特征
研究区位于福建北部海域,是东海内陆架泥质沉积区南部,主要受到冬季以东北季风和夏季以东南季风的季节性交替为特征的东亚季风环流影响。海底近岸地形复杂,整体由近岸向外海倾斜,等深线走向为NE-SW,基本平行于海岸线[16]。
近岸港湾岛屿众多,海岸线曲折。研究区西部靠近闽江口,有大量闽江水和三沙湾水注入,其东部海域主要受台湾暖流(北向的黑潮分支及南海水)和闽浙沿岸流两种水团控制。台湾暖流终年向北,其底层水体几乎影响整个海域,以高温高盐为特征。闽浙沿岸流起源于长江口和杭州湾一带,由沿途入海径流加入靠近闽浙近岸的一支海流,受季风变化明显,具低温、低盐、高溶解氧特征[17]。夏季,受西南季风影响,闽浙沿岸流减弱,除闽江口有明显的东北向低盐水舌外,研究海域基本受台湾暖流水所控制,高盐水基本覆盖西侧沿岸20 m等深线以东区域[18]。冬季,闽浙沿岸流影响范围较大,在冬季强盛东北季风下,扩展范围可影响至广东南澳岛附近海域[17](图1a)。
福建北部海域处于近岸浅水的细粒沉积带,主要为粉砂质泥和泥质粉砂沉积[19],是东海内陆架泥质区的南端,沉积物大部分属于泥质粉砂沉积,呈带状沿海岸线分布,只有小部分为粉砂区[20-21](图1b)。
2 材料与方法
本研究材料取自于福建北部海域26°~27°N、120°~121.2°E,研究区域内基本按等间距设计了145个研究站位,对沉积物开展底栖有孔虫化石种群相关分析研究。所有样品按照微体古生物研究的常规实验方法处理:首先将样品置于培养皿内,在45 ℃恒温下烘干,称量20~50 g干样置于500 mL烧杯中,加入浓度为10%的H2O2溶液浸泡24 h,使样品充分分散。然后将分散好的样品用250目的标准铜筛进行冲洗,直至样品中泥质碎屑被冲尽为止。收集筛上部分置于45 ℃恒温烘干箱,最后取烘干样品备于体视镜下鉴定。原则上每个样品统计不低于200枚壳体。鉴定过程中,若样品中所含有孔虫壳体数目很大,统计过于费时,预先对样品进行缩分处理。底栖有孔虫属种鉴定和分类主要依据附近研究区域参考文献[19,22]。
利用国际上广泛适用于生物群落结构分析的PAST软件处理分析数据。首先把部分具有相似生态环境意义的同属不同种底栖有孔虫合并到属,剔除稀有种(只在个别样品中出现的种),并选择至少在3个样品中含量大于2%的属种,最后得到32个种(属)的百分含量数据,同时对其进行因子分析;综合划分底栖有孔虫生态组合;最后分析各个底栖有孔虫组合的分布环境,深入了解控制底栖有孔虫组合分布的水团环境。

图1 研究区域水文(a)与取样位置(b)[23]Fig.1 Location maps showing water dynamics (a) and sampling sites (b)[23]
3 结果
3.1 底栖有孔虫分布
本研究共鉴定统计底栖有孔虫37属52种(图2—3)。底栖有孔虫全群中占2%以上的优势种共14个,分别是Ammonia beccariivars.(11.69%)、Elphidium advenum(11.52% )、Bolivina robusta(8.52%)、Elphidium hispidulum(6.19%)、Rotalinoides compressiuscula(5.02% ) 、Ammonia pauciloculata(5.51% ) 、Fursenkoina schreibersiana(4.80% ) 、Bulimina marginata(4.42% ) 、Bolivina spathulata(3.77%)、Hanzawaia nipponica(3.76%)、Rotalidium annectens(2.86%)、Heterolepa dutemplei(2.52%)、Buliminasp.(2.39%)、Floriluscf.atlanticus(2.01%)。
研究区底栖有孔虫丰度一定程度上符合随着离岸距离增大而增加的一般规律[19],总分布趋势呈现从西北到东南,从近岸到外海递增的趋势。研究区内145个站位样品间的底栖有孔虫丰度(每克干样中的个体数)变化较大,每克干样中底栖有孔虫最高可达1 626枚,平均每个样丰度为110枚/g,丰度高于100枚/g的站位有28个,在50~100枚/g范围的有20个,小于50枚/g的站位占绝大多数,共97 个(图4a)。
研究区底栖有孔虫的简单分异度为4—34种,其分布与丰度格局基本相似,自西北到东南递增,其分布的控制因素主要以水深为主,另外分异度在研究区出现两个高值区,一个是在研究区的中部区域,第2个是在研究区的西南部区域(图4b)。
本研究区位于东海内陆架泥质沉积区,主要以玻璃壳体类型为主,最多占100%,最少占44.9%,其平均含量占底栖有孔虫总量的90.6%。其次是瓷质壳类型也较为常见,最多占29.3%,平均7.7%;胶结壳质底栖有孔虫极少出现,在101个站位中缺失。在145个站位中共发现玻璃质壳底栖有孔虫30属42种,其中143个站位玻璃质壳含量在50%以上,玻璃质壳体百分含量在研究区分布较平稳。水深小于40 m区域,玻璃质壳体含量变化较大,绝大多数站位含量在75%以上,部分站位达85%~100%。研究区水深40~60 m海域玻璃质壳体含量基本大于85%,而水深大于60 m的深水区域玻璃质壳体含量变化不大,基本在90%以上。研究区共发现瓷质壳类底栖有孔虫4属7种,平均占底栖有孔虫总量的7.7%。其中134个站位瓷质壳类含量在20%以下,11个站位含量可达20%以上,主要分布于水深小于40 m的近岸浅海;在东南部水深大于40 m的海域瓷质壳含量急剧减少(图5)。

图2 研究区部分有孔虫(图版1)(比例尺=100 μm。 1Textularia sagittula(Defrance),2 Martinottiella communis(d' Orbigny),3 Martinottiella sp.,4 Sigmoilopsis asperula(Karrer),5 Quinqueloculina seminula(Linné),6-7 Quinqueloculina akneriana rotunda(Gerke),8-10 Quinqueloculina lamarckiana d'Orbigny,11 Lagena doveyensis Haynes,12 Lagena spicata Cushman et McCulloch,13 Dentalina decepta(Bagg),14 Fissurina marginata(Montagu),15-16 Bolivina robusta Brady,17-18 Bulimina marginata d' Orbigny,19 Virgulopsis orientalis Ho et Hu,20 Uvigerina asperula Czjek,21 Uvigerina aculeata d' Orbigny,22 Rosalina sp.,23 Pseudorotalia indopacifica(Thalmann),24-25 Rotalinoides compressiuscula(Brady))Fig.2 Some foraminifera species in the research area(Plate 1)
3.2 底栖有孔虫优势种分布
大量研究表明,底栖有孔虫优势种和特征种可以用来指示环境[19]。研究区共选取了6类底栖有孔虫优势种和特征种,它们各自的分布特征显著。A.beccariivars.是研究区内占全群比例最大的种,主要分布在研究区西部,对滨岸浅海具有优越的指示作用[19];E.advenum作为近岸浅水种[19],在研究区中部大量分布;B.robusta为典型的中陆架水团代表种[19],在研究区中东部具有较高丰度;E.hispidulum主要分布在研究区中部,为内陆架沿岸水常见分子[19];A.pauciloculata为浙江沿岸水代表种[24],主要分布在研究区中西部;B.marginata同样在中陆架深水区丰富,其丰度可以作为高有机质通量的指标[25],,其分布高值与B.robusta分布较一致;H.nipponica属于典型的喜暖水种[24],一定程度上可以反映海峡暖流的影响程度,其分布范围与暖水种Hetero.dutemplei较为一致(图6)。

图3 研究区部分有孔虫(图版2)(比例尺=100 μm。1-4 Ammonia beccarii(Linné)vars.,5-8 Rotalidium annectens(Parker & Jones),9 Cancris sp.,10 Cribrinonion porisuturalis S.Y.Zheng,11-12 Elphidium advenum(Cushman),13-14 Fursenkoina schreibersiana(Czjzek),15-16 Florilus cf.atlanticus(Cushman),17-18 Hanzawaia nipponica Asano,19 Gyroidina sp.,20 Heterolepa dutemplei(d' Orbigny))Fig.3 Some foraminifera species in the research area(Plate 2)
3.3 底栖有孔虫组合
为了更准确地查明研究区底栖有孔虫分布与海洋环境之间的关系,利用国际上广泛适用于生物群落结构分析的PAST软件,对筛选后的底栖有孔虫属种数据进行因子分析(表1)。因子分析结果识别出4个对总方差值贡献大于2%的底栖有孔虫因子组合,它们总共解释了总方差的83.25%。
主因子1解释了总方差的64.51%,该因子中具有高因子荷载正值的代表属种是A.beccariivars.,Quinqueloculinaspp.,其 次 是R.compressiuscula,A.pauciloculata。高值区主要分布在研究区西部水深小于40 m的近岸区域。主因子2解释了总方差的9.88%,在该因子中具有最高因子载荷正值的代表属种是B.robusta,其次是因子载荷绝对值大于1的属种,包括H.nipponica,H.mantaensis,B.spathulata,B.marginata,U.asperula,U.aculeata,U.schwageri,主要分布在研究区东南部水深大于50 m区域。主因子3解释了总方差的5.91%,在该因子中具有高因子载荷负值的代表属种E.hispidulum,F.schreibersiana,E.advenum,R.annectens,主要分布在研究区西部水深小于40 m区域,与主因子1分布区交错。主因子4解释了总方差的2.88%,E.advenum和A.pauciloculata为高因子载荷正值代表,而H.nipponica,H.mantaensis,R.compressiuscula为高因子载荷负值代表,其负值代表的环境与上述高因子载荷正值的属种所代表的环境呈负相关,高值区主要集中在主因子1和主因子3分布区之间的过渡区域(图7)。
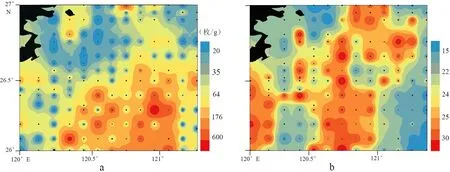
图4 底栖有孔虫丰度(a)及简单分异度图(b)Fig.4 Abundance (a) and simple diversity (b) of benthic foraminifera of the study area
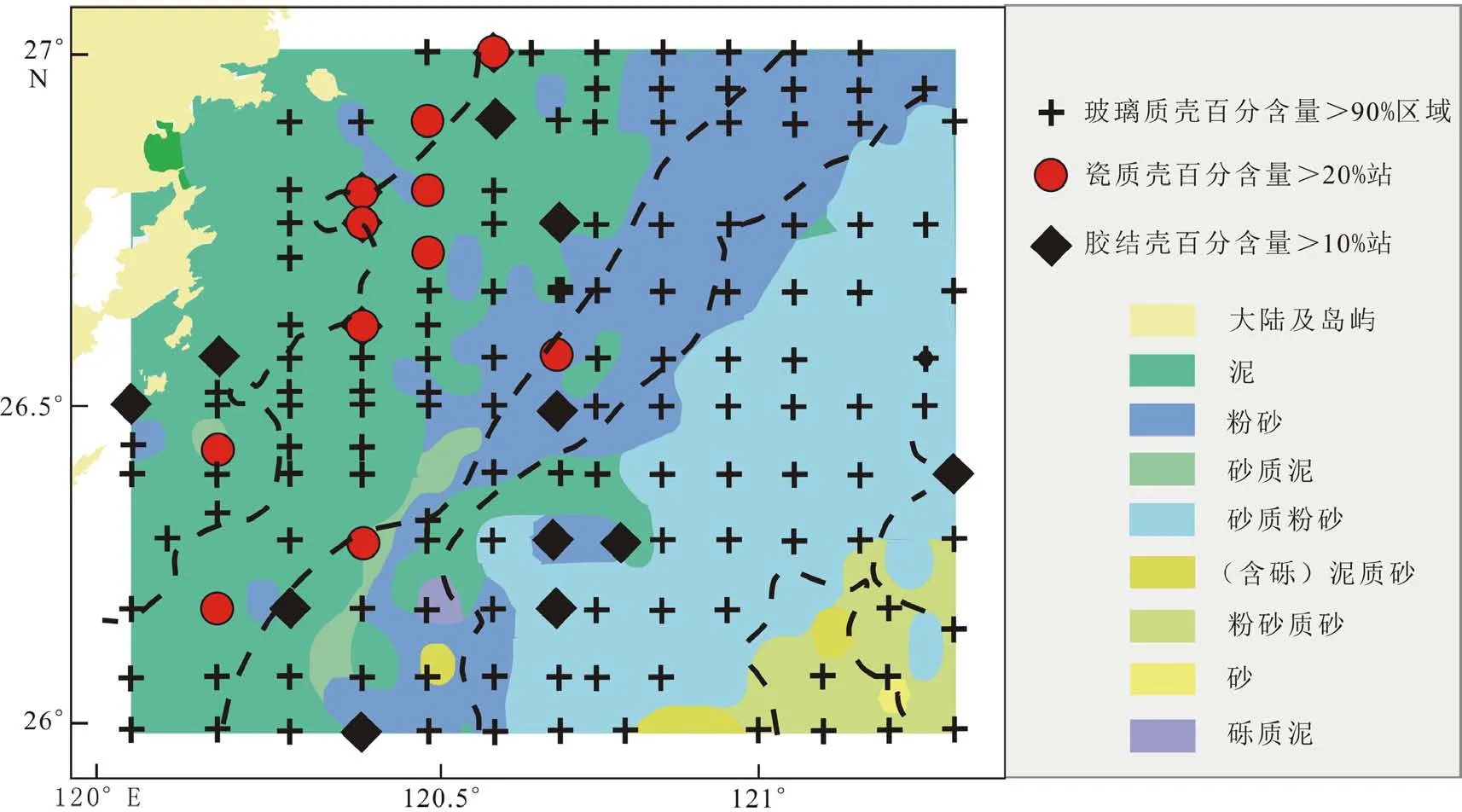
图5 底栖有孔虫三大类壳体分布Fig.5 Distribution of the three types of foraminiferal shells
4 讨论
4.1 底栖有孔虫分布的影响因素
底栖有孔虫的分布受多种环境变量(水深、温度、盐度、营养盐)共同控制[11]。其中水深因素导致的底栖有孔虫分带现象在东海非常明显[19],滨岸指示种A.beccariivars.、浅水种E.advenum、深水种Uvigerinaspp.在研究区内依次沿30、40、60 m等深线分布,说明研究区内水深对底栖有孔虫种群的分布有明显的分异作用;除水深外,盐度对底栖有孔虫分布也有重要影响,东海海区作为盐度正常或接近正常的海域,平均盐度为33%,既不包括潮上带,又无碳酸盐补偿面以下的深海,三大类底栖有孔虫比例一般以玻璃质壳为主,其次是胶结壳质,而且胶结质壳一般在砂质沉积区含量较高,在泥质沉积区含量偏少,为0~5%[19]。研究区西部受闽江冲淡水影响,其底栖有孔虫分布以广温广盐种A.beccariivars.和Quinqueloculina属为主,明显受到低盐水体的控制,其次为数不多的胶结壳质底栖有孔虫也在研究区西北区域分布,与前人在东海的研究分布符合,指示盐度对研究区内底栖有孔虫的分布有影响;其次,有机质通量也是制约底栖有孔虫分带的重要因素,Uvigerinaspp.、B.marginata的丰度可以作为高有机质通量的指标[25].,研究区中部的泥质区沉积物主要来自于长江物质的影响,长江流域带来的巨大水沙通量及有机质可能是造成其丰度高的主要原因[25];最后,调查区的沉积物类型主要为黏土和粉砂质黏土组成的泥,其分布面积约占研究区面积的一半,主要分布在研究区西北近岸区域,有粉砂零星分布在泥质区,另有一条带状泥分布在粉砂和砂质粉砂间[26]。底质类型对底栖有孔虫分布有一定影响,与底栖有孔虫分布关系具有一定的相关性。
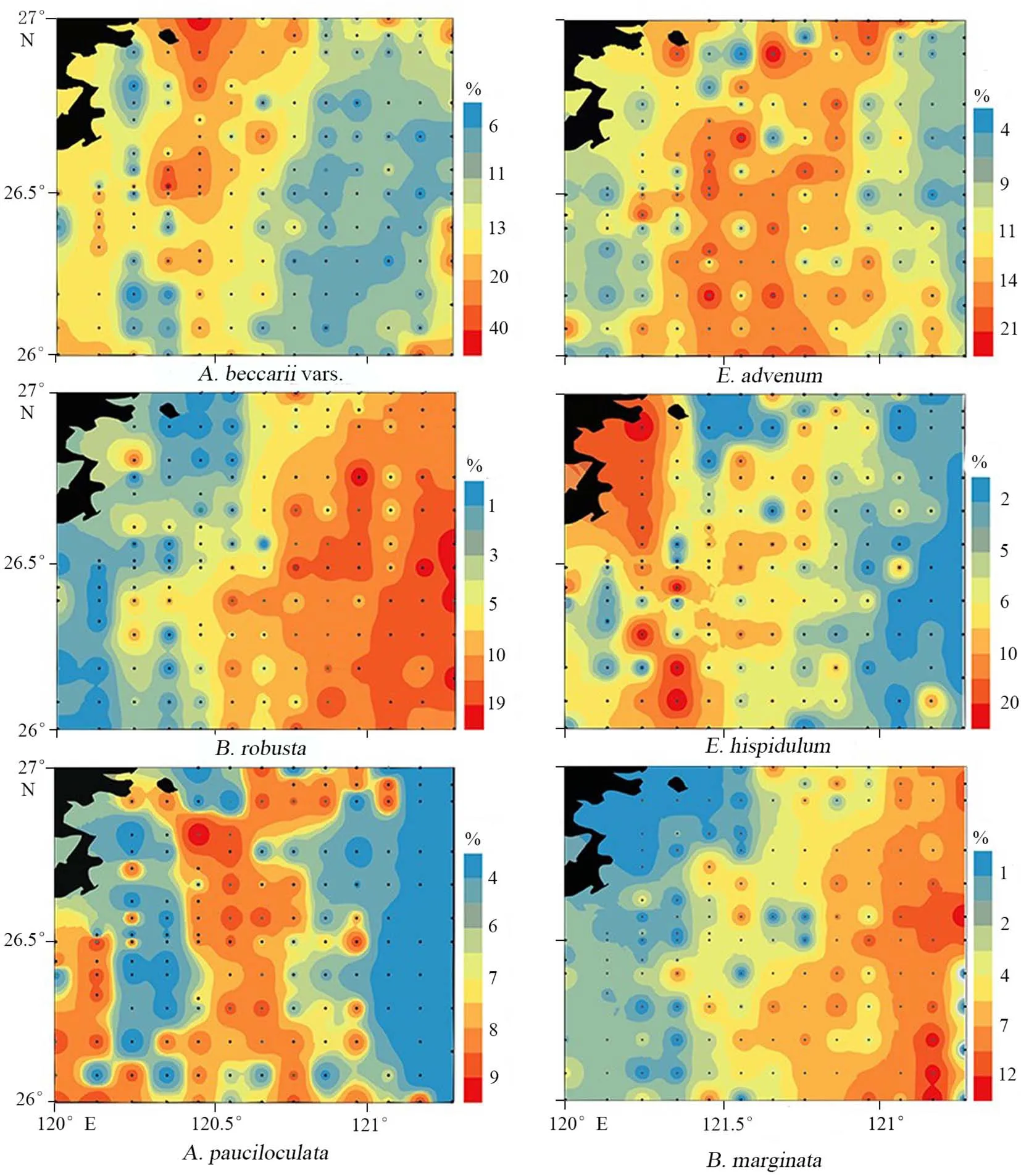
图6 底栖有孔虫优势种相对百分含量分布Fig.6 Relative abundance(%)of dominant benthic foraminifera
4.2 底栖有孔虫组合特征对区域环境的指示
控制底栖有孔虫分布的种种因素归结到一点就是水团的综合特征决定,它决定了研究区沉积物输运与分布、沉积动力条件和有机质输运过程,从而决定了底栖有孔虫在海洋中的分布格局和组合特征,因此不同的有孔虫组合对水体环境具有重要指示意义[17]。本文通过因子分析结果,综合划分不同底栖有孔虫组合,讨论研究区不同水系的区域展布及规模。
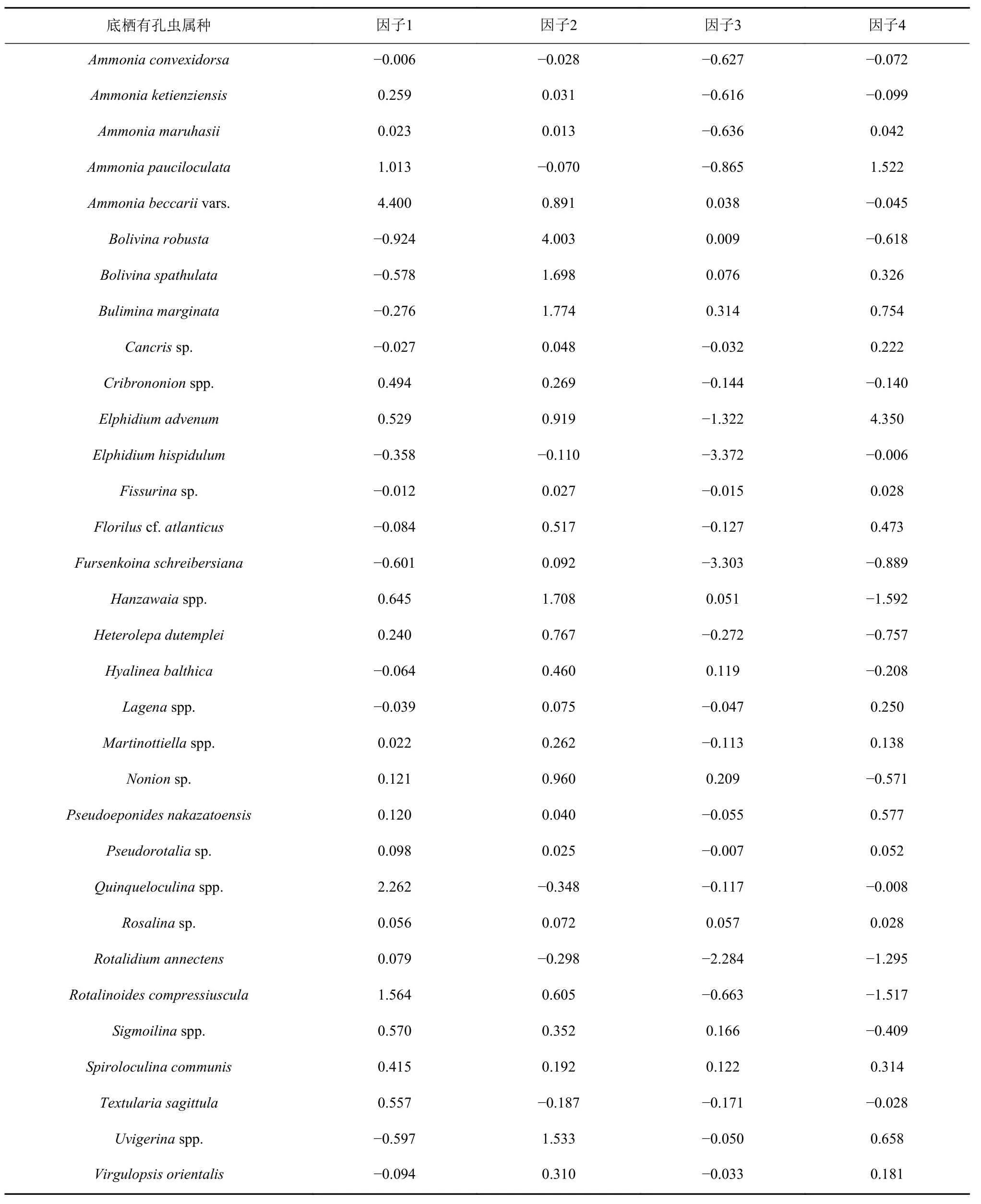
表1 研究区底栖有孔虫因子分析结果Table 1 Factor analysis results of benthic foraminifera
组合Ⅰ(Ammonia beccariivars.-Quinqueloculinaspp.)对应主因子1,以滨岸浅海指示种A.beccariivars.为代表,广泛分布于我国沿海近岸沉积,是典型的近岸浅水型底栖有孔虫类型,对滨岸浅海具有明显的指示作用[19,23]。其次Quinqueloculina属主要出现于水深小于50 m、砂质底质以及水动力相对稳定的海域,一般认为在高盐环境中丰度较高,但也有一些研究认为其对低盐环境也有一定的忍耐性[27]。实验研究表明,温度对物种组成的影响大于盐度,Quinqueloculina属在适宜温度时,盐度与其含量有明显相关关系,与此同时研究中发现Ammonia对低温的忍耐力大于Quinqueloculina,两者在低温下呈现一定的竞争关系,盐度较高时,Quinqueloculina属明显占优势[28]。A.pauciloculata在水深20~30 m分布最多,为浙江沿岸水代表种。A.compressiuscula在东海40~120 m区间最多见,为典型的中陆架代表种[19]。组合Ⅰ(A.beccariivars.-Quinqueloculinaspp.)在黄渤海常见,作为近岸浅水环境代表,受控于水团、盐度、深度以及底质的共同作用[29-30],并且在研究区A.beccariivars.作为优势种百分含量明显大于Quinqueloculina属,指示了低温低盐的沿岸浅海环境。

图7 研究区内底栖有孔虫4个主因子载荷的平面分布Fig.7 Distribution of four factor loadings of benthic foraminifera in the study area
组合Ⅰ主要位于研究区西部,该区域靠近闽江入海口,处于受低温低盐冲淡水注入的偏低盐度环境。这些研究指示的水体条件和闽浙沿岸流水团性质十分相似,因此,推测该区域为主要受闽浙沿岸流水团环境的影响。另外,在研究区西南部发现了少量在深水环境才能出现的Gyroidinasp.和Uvigerinaspp.[14],并聚集在闽江河口区出现分异度高值,在台湾海峡平潭岛附近的研究中,其浅海域出现了底流搬 运 的 少 量Gyroidinasp.、Globobuliminaspp.和Uvigerinaspp.[15]。因此,研究区浅水区出现的少量深水底栖有孔虫可能是受台湾暖流与闽浙沿岸流季节变化造成的上升流带来的深水种,同时闽江口作为强潮河口[30],可能受潮流影响将一些有孔虫属种携入河口,形成外来分子较多的有孔虫群落。
组合Ⅱ(B.robusta-B.marginata-Hanzawaiaspp.)(主因子2)以典型的中陆架水团代表种B.robusta为主,其在东海和台湾海峡多集中在中陆架40~120 m水深区[14,19],为低氧沉积中优势种[31]。H.nipponica在东海主要分布于中外陆架的现代沉积中,属于典型的喜暖水种,一定程度上可以反映海峡暖流的影响程度[19]。B.spathulata和B.marginata在东海常见于陆架区,Bulimina属在中陆架深水区丰富;Uvigerina属在中外陆架少量出现,是典型的深水种,在东海海槽常见[19],并且研究发现列式壳Uvigerina和Bulimina属可指示底层水环境,当其含量高时,指示底层水低氧富营养环境;反之,则指示高氧高有机质通量环境[32]。因此组合Ⅱ为生活在中陆架水深约40~120 m的低氧富营养环境的底栖有孔虫组合,且该区域浮游有孔虫含量在10%以上,最高达45%。沉积物中底栖有孔虫以玻璃质壳为主,瓷质壳和胶结壳含量非常少。该组合中底栖有孔虫丰度、分异度较高,这些特征与内陆架到中陆架正常海相环境的底栖有孔虫分布特征相吻合,而台湾暖流在水深50 m左右形成切变锋[2],阻碍了陆源碎屑沉积物及有机质向海输运的通道,起到“水障”作用[25]。综上所述,组合Ⅱ分布区域为受台湾暖流影响的中陆架较深水环境。
同样分布在研究区水深小于40 m区域的组合Ⅲ(Elphidium hispidulum-Fursenkoina schreibersiana)(主因子3),以E.hispidulum和F.schreibersiana为主,其因子仅解释总方差的5.91%,可能指示了研究区内短期的水文现象。E.hispidulum在东海分布于整个陆架,尤以内陆架居多,为内陆架沿岸水常见分子,也是适应高盐环境的暖水种[19]。Fursenkoina属在浅水50~60 m低氧区丰富,对最小含氧带(OMZ)的深度分布有显著的指示意义[33]。因此,该组合指示低氧高盐的底层沉积环境。
季节性缺氧在世界上许多封闭性海湾、河口和海岸普遍存在,通常认为低物理能、高径流量的海洋环境更容易发生低氧[34]。我国长江口外,珠江口附近的一些近岸海域及东海等海区多发季节性底层低氧现象[35],并有研究发现夏季长江口缺氧区主要出现在有锋面和上升流现象的海域附近[36]。这些低氧区的出现是由于淡咸水形成强烈盐度锋而造成的水体层化底层缺氧,并且较高程度地受季节控制[37]。低温高盐的台湾暖流深层水在6—8月主要盘踞在海峡中、北部海域的底层,并靠近海峡中线偏西一侧北上[38],受地形抬升和季风等因素影响,一年四季均存在上升流现象[2],并在夏季出现特性明显不同的两种或几种水体之间的狭窄过渡带(锋面)[39]。浙江沿岸水和台湾暖流之间,台湾暖流上层水和深层水之间均能够形成峰面,它们的位置会随季节发生变动[40]。在夏季,受西南季风、大量闽江入海水及台湾暖流增强的影响,盐度差异较大的上下水体分层,阻碍了表层富氧水与底层水的混合,底层水供氧不及时,海底沉积物可能处于缺氧甚至厌氧环境。因此,推测研究区低氧区的存在可能与夏季闽江水和外海水混合发生水体层化有关,而Fursenkoina属的分布一定程度上可能反映了夏季增强的台湾暖流深层水在浅海的展布范围。组合III的特征指示了夏季闽浙沿岸受海峡暖流高盐水入侵的沿岸浅水环境。
组合Ⅳ(Elphidium advenum-Ammonia pauciloculata)(主因子4)处于组合Ⅰ和组合Ⅱ过渡区域。其中E.advenum作为近岸浅水种,可指示高氧环境[29]。A.pauciloculata主要分布于水深20~30 m处,为浙江沿岸水的代表种[19]。其正值代表的环境为低温低盐的较浅水内陆架环境。其负值代表的是高温高盐的较深水团环境。该组合指示混合水团过渡环境。该组合的分布区域主要集中于闽浙沿岸流影响较大的组合Ⅰ和明显受海峡暖流影响的组合Ⅱ之间的区域,且组合内底栖有孔虫类型有混合性质,其正值受闽浙沿岸流影响,其负值受海峡暖流影响,表明了该组合的过渡性质,其底栖丰度、分异度都比周边区域偏高,在东海、台湾海峡的水团分析中都曾识别到台湾暖流与沿岸冲淡水的季节性混合水团[41]。推测研究区内组合Ⅳ指示暂时性存在的季节性水团,且大部分时候台湾暖流与闽浙沿岸流的分界明显,可能与闽浙沿岸流和海峡暖流相遇形成混合水团有关。
以上研究表明,不同的底栖有孔虫的组合对应不同的水团环境,水团对其组合分布有明显的制约作用。其中,组合Ⅲ的分布明显反映了水团的溶解氧含量对底栖有孔虫组合分布的影响。以往对闽江的研究并未在其下游和河口发现明显缺氧现象,闽江作为强潮河流,夏季与外海水团混合较好,一般不会形成明显低氧区,但之前有研究报道Fursenkoina fusiformis对污染环境有很强的适应能力[42],对沉积物中重金属调查也显示闽江河口有较多的人类活动污染[5],推测人为活动对福建北部海域出现的相对低氧区有较大影响。研究区海域底栖生物组合复杂,可以指示其水团的消长变化,研究区主要受闽浙沿岸流、台湾暖流水团控制,这些水团的控制区域明显,对研究近岸水团变性、上升流现象和人类活动对环境影响等有重要意义。
5 结论
(1)对福建北部海域网格式等间距表层样品底栖有孔虫分析结果表明,该区底栖有孔虫丰度和分异度变化较大。底栖有孔虫组成以玻璃质壳体为主,其平均含量占底栖有孔虫总量的90.6%。其次是瓷质壳类型,平均含量为7.7%;胶结质壳底栖有孔虫仅在少数站位出现。
(2)研究区底栖有孔虫丰度一定程度上符合随着离岸距离增大而增加的一般规律,总分布趋势呈现从西北到东南、从近岸到外海递增的趋势。
(3)研究区海域的底栖有孔虫组合明显受到水团制约,对底栖有孔虫类型进行因子分析表明,组合Ⅰ(A.beccariivars.-Quinqueloculinaspp.)指示受闽浙沿岸水团影响的环境;组合Ⅱ(B.robusta-B.marginata-Hanzawaiaspp.)指示台湾暖流水团影响下的环境;组合Ⅲ(E.hispidulum-F.schreibersiana)可能与夏季闽江水与外海水混合发生水体层化有关;组合Ⅳ以E.advenum-A.pauciloculata为主,分布于组合Ⅱ与组合Ⅲ的过渡区域,指示受季节性闽浙沿岸水与台湾暖流混合水团共同影响的环境。
参考文献(References)
[1]伍伯瑜.台湾海峡及其邻近水域的流型和水文特征[J].海洋通报,1983,2(4):1-8.[WU Boyu.The current pattern and hydrologic character in the Taiwan Straits and its adjacent waters [J].Marine Science Bulletin,1983,2(4): 1-8.]
[2]潘玉球,曹欣中,许建平.浙江沿岸上升流锋区特征及其成因的初步探讨[J].海洋湖沼通报,1982(3):1-7.[PAN Yuqiu,CAO Xinzhong,XU Jianping.A preliminary investigation of the cause and characteristics of the upwelling front zone off Zhejiang [J].Transactions of Oceanology and Limnology,1982(3): 1-7.]
[3]曾刚.福建近海沿岸水及其水文状况[J].海洋通报 ,1986,5(3):32-37.[ZENG Gang.The coastal water and its hydrographic states in the nearshore waters along Fujian province [J].Marine Science Bulletin,1986,5(3): 32-37.]
[4]Li C S,Shi X F,Kao S J,et al.Rare earth elements in fine-grained sediments of major rivers from the high-standing island of Taiwan [J].Journal of Asian Earth Sciences,2013,69: 39-47.
[5]Liu S F,Shi X F,Liu Y G,et al.Concentration distribution and assessment of heavy metals in sediments of mud area from inner continental shelf of the East China Sea [J].Environmental Earth Sciences,2011,64(2): 567-579.
[6]Qiao S Q,Yang Z S,Liu J P,et al.Records of late-Holocene East Asian winter monsoon in the East China Sea: Key grain-size component of quartz versus bulk sediments [J].Quaternary International,2011,230(1-2): 106-114.
[7]沈林南,吴祥恩,李超,等.福建三沙湾表层沉积硅藻分布特征及其与环境因子的关系[J].应用海洋学学报,2014,33(2):212-221.[SHEN Linnan,WU Xiang’en,LI Chao,et al.Distribution characteristics of diatom in surface sediment and its relation with environment factors in Sansha Bay of Fujian [J].Journal of Applied Oceanography,2014,33(2): 212-221.]
[8]田丰歌,徐兆礼.福建中部近海浮游动物数量分布与水团变化的关系[J].生态学报,2012,32(4):1097-1104.[TIAN Fengge,XU Zhaoli.Relating the distribution of zooplankton abundance in the coastal waters of central Fujian Province to the seasonal variation of water masses [J].Acta Ecologica Sinica,2012,32(4): 1097-1104.]
[9]Li X Y,Jian Z M,Shi X F,et al.A Holocene record of millennial-scale climate changes in the mud area on the inner shelf of the East China Sea [J].Quaternary International,2015,384: 22-27.
[10]汪品先,闵秋宝,卞云华,等.东海表层沉积中有孔虫、介形虫组合分布的初步研究[J].同济大学学报,1979(2):90-108.[WANG Pinxian,MIN Qiubao,BIAN Yunhua,et al.A preliminary study of Foramiriferal and Ostracod assemblages' distribution in bottom sediments of the East China Sea [J].Journal of Tongji University,1979(2): 90-108.]
[11]郑执中,郑守仪,傅钊先.东海底栖有孔虫区系的初步分析[J].科学通报,1979,24(19):903-906.[ZHENG Zhizhong,ZHENG Shouyi,FU Zhaoxian.Preliminary analysis of benthic foraminifera in the East China Sea [J].Chinese Science Bulletin,1979,24(19): 903-906.]
[12]张江勇,汪品先.深海研究中的底栖有孔虫: 回顾与展望[J].地球科学进展,2004,19(4):545-551.[ZHANG Jiangyong,WANG Pinxian.Benthic foraminifera in deep-sea research: retrospect and prospect [J].Advances in Earth Science,2004,19(4): 545-551.]
[13]陈荣华.浙江海岸带表层沉积物中有孔虫的分布及其影响因子[J].东海海洋,1990,8(3):35-48.[CHEN Ronghua.Distribution and influencing factors of foraminifera in surface sediments of coastal zone of Zhejiang Province [J].Donghai Marine Science,1990,8(3): 35-48.]
[14]方惠瑛.台湾海峡表层沉积物中底栖有孔虫分布特征[J].台湾海峡,1998(1):43-49.[FANG Huiying.Benthic foraminifera in surface sediments of Taiwan Strait [J].Journal of Oceanography in Taiwan Strait,1998(1): 43-49.]
[15]周洋,陈芳,孙桂华,等.台湾海峡西北部平潭岛海域表层沉积物中底栖有孔虫分布及其环境控制因素[J].微体古生物学报,2014,34(1):54-63.[ZHOU Yang,CHEN Fang,SUN Guihua,et al.Benthic foraminiferal distribution in surface sediments of the northwest Taiwan Strait near the Pingtan Island and its environmental controlling factors [J].Acta Micropalaeontologica Sinica,2014,34(1): 54-63.]
[16]陈峰.福建近岸海底地形的初步研究[J].台湾海峡,1982(2):83-90.[CHEN Feng.A preliminary study of underwater morphology along the Fujian coast [J].Journal of Oceanography in Taiwan Strait,1982(2): 83-90.]
[17]王翠,郭晓峰,方婧,等.闽浙沿岸流扩展范围的季节特征及其对典型海湾的影响[J].应用海洋学学报,2018,37(1):1-8.[WANG Cui,GUO Xiaofeng,FANG Jing,et al.Characteristics of seasonal spatial expansion of Fujian and Zhejiang Coastal Current and their bay effects [J].Journal of Applied Oceanography,2018,37(1): 1-8.]
[18]Jan S,Wang J,Chern C S,et al.Seasonal variation of the circulation in the Taiwan Strait [J].Journal of Marine Systems,2002,35(3-4): 249-268.
[19]汪品先,章纪军,赵泉鸿,等.东海底质中的有孔虫和介形虫[M].北京: 海洋出版社,1988.[WANG Pinxian,ZHNG Jijun,ZHAO Quanhong,et al.Foraminifera and Ostracods in the Sediments of the East China Sea[M].Beijing: China Ocean Press,1988.]
[20]石学法,刘升发,乔淑卿,等.东海闽浙沿岸泥质区沉积特征与古环境记录[J].海洋地质与第四纪地质,2010,30(4):19-30.[SHI Xuefa,LIU Shengfa,QIAO Shuqing,et al.Depositional features ang palaeoenvironmental records of the mud deposits in Min-Zhe coastal mud area,East China sea [J].Marine Geology and Quaternary Geology,2010,30(4): 19-30.]
[21]Li G X,Li P,Liu Y,et al.Sedimentary system response to the globalsea level change in the East China Seas since the last glacial maximum [J].Earth Science Reviews,2014,139: 390-405.
[22]Lei Y L,Li T G.Atlas of Benthic Foraminifera from China Seas[M].Berlin,Heidelberg: Springer,2016.
[23]丛静艺,袁忠鹏,胡刚,等.长江远端三角洲多源沉积分异作用及其动力机制[J].沉积学报,2020,38(3):528-537.[CONG Jingyi,YUAN Zhongpeng,HU Gang,et al.Sedimentary differentiation and hydrodynamic environment of multi-sourced sediment in the Changjiang Distal Delta [J].Acta Sedimentologica Sinica,2020,38(3): 528-537.]
[24]史光辉.东海陆架泥质区底栖有孔虫记录及其环境意义[D].中国海洋大学硕士学位论文,2014.[SHI Guanghui.Benthic foraminifera response to changes in paleoenviornments of Mud area on the East China Sea shelf[D].Master Dissertation of Ocean University of China,2014.]
[25]Jian Z M,Wang L J,Kienast M,et al.Benthic foraminiferal paleoceanography of the South China Sea over the last 40,000 years [J].Marine Geology,1999,156(1-4): 159-186.
[26]类彦立,李铁刚.奥茅卷转虫 Ammonia aomoriensis (Asano,1951)与毕克卷转虫 Ammonia beccarii (Linnaeus,1758)(有孔虫)的分类学以及在黄东海分布的温盐深特征比较研究[J].微体古生物学报,2015,32(01):1-19.[LEI Yanli,LI Tiegang.Ammonia aomoriensis (asano,1951) and Ammonia beccarii (linnaeus,1758) (foraminifera) :comparisons on theirtaxonomy and ecological distributions correlated to temperature,salinity and depth in the yellow SEA [J].Acta Micropalaeontologica Sinica,2015,32(01): 1-19.]
[27]Murray J W.Ecology and Palaeoecology of Benthic Foraminifera[M].Harlow: Longman Scientific and Technical,1991.
[28]Dong S S,Lei Y L,Li T G,et al.Responses of benthic foraminifera to changes of temperature and salinity: Results from a laboratory culture experiment [J].Science China Earth Sciences,2019,62(2): 459-472.
[29]李小艳,石学法,程振波,等.渤海莱州湾表层沉积物中底栖有孔虫分布特征及其环境意义[J].微体古生物学报,2010,27(1):38-44.[LI Xiaoyan,SHI Xuefa,CHENG Zhenbo,et al.Distribution of benthic foraminifera in surface sediments of the Laizhou Bay,Bohai Sea and its environmental significance [J].Acta Micropalaeontologica Sinica,2010,27(1): 38-44.]
[30]王海霞,赵全民,李铁刚,等.辽东湾表层沉积物中底栖有孔虫分布及其与沉积环境的关系[J].海洋地质与第四纪地质,2011,31(2):87-94.[WANG Haixia,ZHAO Quanmin,LI Tiegang,et al.Distribution of benthic foraminifera in surface sediments of the Liaodong Bay and its bearing on sedimentary environments [J].Marine Geology and Quaternary Geology,2011,31(2): 87-94.]
[31]Zhao B C,Yan X X,Wang Z H,et al.Sedimentary evolution of the Yangtze River mouth (East China Sea) over the past 19,000 years,with emphasis on the Holocene variations in coastal currents [J].Palaeogeography,Palaeoclimatology,Palaeoecology,2018,490: 431-449.
[32]Kaiho K.Benthic foraminiferal dissolved-oxygen index and dissolvedoxygen levels in the modern ocean [J].Geology,1994,22(8): 719-722.
[33]Mazumder A,Nigam R.Bathymetric preference of four major genera of rectilinear benthic foraminifera within oxygen minimum zone in Arabian Sea off central west coast of India [J].Journal of Earth System Science,2014,123(3): 633-639.
[34]刘海霞,李道季,高磊,等.长江口夏季低氧区形成及加剧的成因分析[J].海洋科学进展,2012,30(2):186-197.[LIU Haixia,LI Daoji,GAO Lei,et al.Study on main influencing factors of formation and deterioration of summer hypoxia off the Yangtze River Estuary [J].Advances in Marine Science,2012,30(2): 186-197.]
[35]王巧宁,颜天 ,周名江.近岸和河口低氧成因及其影响的研究进展[J].海洋环境科学,2012,31(5):775-778.[WANG Qiaoning,YAN Tian,ZHOU Mingjiang.Research progress on cause of hypoxia and its influence in coastal and estuary region [J].Marine Environmental Science,2012,31(5): 775-778.]
[36]韦钦胜,王保栋,于志刚,等.夏季长江口外缺氧频发的机制及酸化问题初探[J].中国科学: 地球科学,2017,60(2):360-381.[WEI Qinsheng,WANG Baodong,YU Zhigang,et al.Mechanisms leading to the frequent occurrences of hypoxia and a preliminary analysis of the associated acidification off the Changjiang estuary in summer [J].Science China Earth Sciences,2017,60(2): 360-381.]
[37]叶丰,黄小平.近岸海域缺氧现状、成因及其生态效应[J].海洋湖沼通报,2010(3):91-99.[YE Feng,HUANG Xiaoping.The status,causes,and ecological effects of coastal hypoxia [J].Transactions of Oceanology and Limnology,2010(3): 91-99.]
[38]翁学传,张启龙,颜廷壮,等.台湾海峡中、北部海域春、夏季水团分析[J].海洋与湖沼,1992,23(3):235-244.[WENG Xuechuan,ZHANG Qilong,YAN Tingzhuang,et al.Analysis of water masses in the middle and northern Taiwan strait in spring and summer [J].Oceanologia et Limnologia Sinica,1992,23(3): 235-244.]
[39]许金电,黄奖,邱云,等.浙闽沿岸水的空间结构特征及生消过程[J].热带海洋学报,2015,34(1):1-7.[XU Jindian,HUANG Jiang,QIU Yun,et al.Spatial structure characteristics of Zhejiang and Fujian coastal water and their evolution [J].Journal of Tropical Oceanography,2015,34(1): 1-7.]
[40]潘玉球,徐端蓉,许建平.浙江沿岸上升流区的锋面结构、变化及其原因[J].海洋学报,1985,7(4):401-411.[PAN Yuqiu,XU Ruirong,XU Jianping.Preliminary study on the characteristics and causes of upwelling front along the coast of Zhejiang province [J].Acta Oceanologica Sinica,1985,7(4): 401-411.]
[41]喻祖祥,苏育嵩,俞光耀,等.长江口及济州岛附近海域变性水团的初步分析[J].山东海洋学院学报,1984,14(3):1-12.[YU Zuxiang,SU Yusong,YU Guangyao,et al.A preliminary analysis of modified water masses in the sea area near the Yangtze River mouth and the Chijudo island [J].Journal of Shandong College of Oceanology,1984,14(3): 1-12.]
[42]桂峰,王柳柱,赵晟,等.舟山港表层沉积物中底栖有孔虫组合特征初步研究[J].微体古生物学报,2013,30(4):415-422.[GUI Feng,WANG Liuzhu,ZHAO Shen,et al.Benthic foraminiferal characteristics in surface sediments of the Zhoushan harbour,Zhejiang,China [J].Acta Micropalaeontologica Sinica,2013,30(4): 415-422.]
