优异高粱雄性不育系01-26A的组配降秆效应及其分子机理
2020-08-15邹剑秋王艳秋李金红朱凯
邹剑秋,王艳秋,李金红,朱凯
优异高粱雄性不育系01-26A的组配降秆效应及其分子机理
邹剑秋,王艳秋,李金红,朱凯
(辽宁省农业科学院高粱研究所,沈阳 110161)
【】高粱机械化生产是未来发展的必然方向,而合理的株型是机械化生产的基础与关键。育种过程中发现,矮秆雄性不育系01-26A具有和现有粒用高粱恢复系组配F1都能降低株高的独特优势,是一个极其难得的株型调控材料。因此,对其株高遗传效应及调控基因位点进行研究,旨在探明其株高矮化遗传机理和调控机制,以期应用遗传育种手段促进高粱株型优化提供理论依据。以具有矮化株高效应的01-26A和不具矮化株高效应的7050A高粱雄性不育系为试材,重点对其与7个(包括6个粒用和1个甜高粱)恢复系的杂种F1的株高及节数、穗柄下茎秆高度、穗柄长和穗长等相关参数的遗传效应进行分析,同时对调控株高基因的位点Dw、Dw和Dw(Dw暂未被克隆)进行测定与分析。高粱雄性不育系01-26A(A1细胞质)具有显著的矮化粒用高粱株高的效应,以其为母本组配的杂种F1株高较以高粱雄性不育系7050A(A2细胞质)为母本组配的杂种F1株高降幅为15.8%,绝对值一般不超过160 cm,而以其为母本组配的甜高粱杂种F1株高降低不明显,不具矮化效应;01-26A矮化株高遗传效应主要表现在杂种F1穗柄下茎秆高度明显缩短,茎秆中下部节间长度与株高变化相关性较高;01-26A杂种F1穗柄长降低是造成株高变矮的另一原因,其效应小于穗柄下茎秆高度,而穗长对株高变化的影响很小;通过基因位点的序列和类型分析,确定了01-26A矮化基因Dw—Dw的基因类型,并通过多个杂交组合的株高遗传数据分析,推断01-26A基因型很可能是dwdwDwDwdwdwdwdw,即三矮高粱不育系;另外,通过对株高调控基因研究分析,发现01-26A的dw和dw矮化基因可能对粒用高粱杂种F1株高影响效应更大,而Dw的存在是造成其与甜高粱杂种F1株高没有矮化的内在原因。高粱雄性不育系01-26A可能是具有dwdwDwDwdwdwdwdw基因的三矮高粱不育系,可通过降低杂种F1穗柄下茎秆高度(主效)和穗柄长(次效),实现高粱株高的矮化调控;但其与甜高粱杂交,可能由于Dw的存在,F1并未发现明显的矮化效应。
高粱;株高;遗传效应;基因位点
0 引言
【研究意义】高粱是中国重要的粮食、饲料作物和酿酒原料,是世界上仅次于小麦、玉米、水稻、大麦的第五大作物,并具有抗旱、耐涝、耐盐碱、耐瘠薄等多重抗性和强大的杂种优势[1-2]。随着生产水平的不断提高,高粱机械化生产成为未来高粱产业持续健康发展的必然方向,而现阶段中国适宜机械化栽培高粱资源较少,很多优良的高粱资源因组配的杂交种株高较高,影响了其利用效率和利用潜力,导致机械化专用品种选育相对落后,限制了高粱产业的进一步提升[3-5]。因此,开展高粱株高矮化遗传效应及调控基因位点研究,解析高粱株高的遗传机理,以便更加有针对性地应用高粱资源,对高粱株高进行科学合理地调控,促进高粱产业向机械化和规模化发展尤为重要。【前人研究进展】关于高粱株高的调控效应,学者们开展了大量研究,曾有报道气候条件、种植密度和理化调控等手段会对株高造成较大影响[6];而更多人从株高遗传方面开展了研究,有研究认为株高受茎秆节间长度、节数、穗柄长和穗长等因素影响[7-9];也有报道认为高粱的株高基因是所有已知的最有用的遗传因子之一,可使粒用高粱植株变矮,但对甜高粱株高的矮化效应并不明晰[10];ROSS等[11]研究表明高粱株高的调控基因主要有4个,即—。另外,卢庆善等[12]提出通过选择具有株高矮化基因的高粱资源与其他资源组配,可显著降低杂种F1的株高,并且确定了4对非连锁矮化基因控制高粱株高的遗传,即高秆与矮秆品种杂交,在杂种一代高秆对矮秆表现为部分显性,同时将株高分为5个等级:0-矮级、1-矮级、2-矮级、3-矮级、4-矮级。此外,学者在调控高粱株高的Dw—Dw4个基因作用效应方面开展了研究,指出Dw和Dw在调控茎秆节间长度方面效应明显,而作为在高粱上第一个被克隆的矮化基因,除对茎秆节间长度存在调控外,还对分蘖数、抗倒伏和产量性状具有显著影响[9]。Dw至今还没有被克隆,但曾在帚高粱中被检测到[13],虽没有深度报道,但分析可能对穗柄长具有影响,进而调控株高。同时,大量研究表明,株高矮化调控同时受一个或多个基因共同作用,高粱在株高调控中往往是由Dw—Dw协同完成[14]。【本研究切入点】迄今为止,在已收集到的全世界高粱资源中,虽然资源非常丰富,株高的幅度在55—655 cm,但在资源利用方面还存在很多问题,且先前的研究表明,从株高遗传研究角度而言,大多数高粱资源依然是1-矮基因型和2-矮基因型材料[15],3-矮基因型和4-矮基因型材料相对较少。另外,虽然前人在高粱株高调控方面开展了许多研究,但仍然缺少株高组配降秆效应的系统研究。【拟解决的关键问题】本研究利用育种过程中发现的具有和现有粒用高粱恢复系组配F1都能降低株高的矮秆雄性不育系01-26A为株型调控材料,并且选择不具矮化株高效应的不育系7050A作为对照,系统开展株高矮化遗传效应和调控基因位点研究。旨在探明01-26A矮化基因关键调控位点(包括Dw、Dw和Dw,而Dw至今还未被克隆),解析高粱株高的遗传机理,从遗传角度解决高粱株高矮化调控问题,促进高粱向机械化、规模化发展。
1 材料与方法
1.1 试验材料
1.1.1 2个高粱雄性不育系和7个高粱恢复系 2个高粱雄性不育系,包括具有矮化株高效应的01-26A和不具矮化株高效应的7050A,其中,01-26A具有一般配合力好、特殊配合力高、高抗丝黑穗病、抗蚜虫、抗叶斑病、抗旱、抗涝、耐瘠薄、活秆成熟等特点,最重要的是其具有和所有粒用高粱恢复系组配F1都能降低株高的特性。7个高粱恢复系,包括粒用高粱恢复系6个(LNR-4、NK1、0-01、3535、3550和BR92)和甜高粱恢复系1个(LTR168)。以上9个材料的系谱见表1。
1.1.2 84个杂交种 由2个雄性不育系和7个恢复系不完全双列杂交的14个杂交种F1;粒用杂交组合(F1)57个,其中,01-26A与恢复系杂交组合33个,7050A与与恢复系杂交组合24个;甜高粱F1杂交组合13个,其中,与01-26A组配7个,与7050A组配6个。
1.2 试验设计
试验于2016—2018年连续3年在辽宁省农业科学院研究基地进行,对2个不育系、7个恢复系及其不完全双列杂交形成的14个杂交组合进行种植。随机区组设计,6行区,行长3 m,行宽0.6 m,小区面积10.8 m2,3次重复。完全抽穗后测定株高和株高参数,其中株高参数包括穗柄下茎秆高、穗柄长、穗长、茎秆节数和节间长。
2019年对以上参试材料在辽宁省农业科学院人工气候室(温度:25℃,湿度:45%,光照12 h/黑暗12 h)进行幼苗培养,进行矮化基因提取与分析。
此外,2018年对01-26A与粒用恢复系杂交33个组合、7050A与粒用恢复系杂交24个组合、01-26A及7050A与甜高粱恢复系杂交13个组合的株高进行了测定,进一步验证01-26A的株高矮化效应。
1.3 测定项目与方法
1.3.1 株高及其参数测定 待高粱穗完全抽出后,采用伸缩性直尺测定株高,株高测定时去掉边行,每个高粱品种(品系)测定3株,求其平均值。同时,对2个不育系、7个恢复系(包括粒用高粱和甜高粱)及其不完全双列杂交形成的14个杂交组合进行株高参数测定,将每小区测定株高的3个植株去掉叶和叶鞘,分离穗、穗柄和穗柄下茎秆,分别用直尺测定其长度并求平均值,之后将3年结果求平均值。
1.3.2 株高基因Dw、Dw和Dw的检测 在人工气候室进行幼苗培养,六叶期取叶片进行矮化基因提取与分析。利用改良CTAB法提取三叶期高粱幼苗DNA,根据https://phytozome.jgi.doe.gov/pz/portal.html#! search?show=KEYWORD&method=Org_Sbicolor网站公布的调控株高的基因序列设计引物(表2),进行dw、dw和dw的扩增,扩增体系为Mix 10 μL primer(100 μmol·L-1)0.5 μL、DNA 0.5 μL和ddH2O 8.25 μL。扩增条件为94℃2 min;94℃30 s,55℃ 30 s,72℃ 45 (120) s,32个循环;72℃10 min。dw和dw的PCR产物送至大连TaKaRa公司测序,然后用DNAman软件进行序列分析。dw的PCR产物进行琼脂糖检测。

表1 2个高粱雄性不育系和7个高粱恢复系的系谱
1.4 数据分析
采用DPS7.05和Excel 2007软件对亲本和F1各参数进行统计分析,计算变异系数(coefficient of variation,)。变异系数(%)=/,为杂交F1数据的标准差,为杂交F1数据的平均值。
2 结果
2.1 株高及其参数遗传分析
2.1.1 株高遗传分析 参试的01-26A和7050A 2个高粱雄性不育系与6个粒用高粱恢复系组配的F1代杂交种株高存在显著差异(表3)。值得注意的是,01-26A与6个粒用高粱恢复系组配杂交种株高变幅为141.3—159.7 cm,变异系数()为13.6%—22.9%,而7050A组配的杂交种株高变幅为159.6—185.7 cm,为10.9%—23.7%,01-26A组配的粒用高粱F1杂交种株高的平均值比7050A组配的粒用高粱F1杂交种降低了15.8%。说明01-26A对杂种F1具有显著的矮化效应。F1株高与亲本关联分析发现,01-26A组配的粒用高粱F1杂交种株高基本在双亲之间或略高于双亲最高值,而7050A组配的粒用高粱F1杂交种则均表现为高于双亲最高值。

表2 PCR引物扩增序列

表3 不育系、恢复系及其F1株高遗传效应
**:<0.01;*:<0.05;ns:差异不显著;+:株高在这一范围;—:株高不在这一范围;PH:株高。下同
**:<0.01; *:<0.05; ns: difference is not significant; +: the plant height is in this range; —: the plant height is not in this range; PH: plant height. The same as below
与甜高粱恢复系组成的杂交F1中,01-26A和7050A与甜高粱恢复系LTR168的杂种F1株高增加明显,均明显高于双亲最高值,但二者差异很小。说明01-26A与甜高粱杂交株高遗传中,没有明显的株高矮化遗传效应。
2.1.2 穗柄下茎秆高度遗传分析 01-26A和7050A与粒用高粱恢复系组配的F1穗柄下茎秆高度存在显著差异(表4)。雄性不育系01-26A比7050A穗柄下茎秆高度低35.9%,它们分别与6个粒用高粱恢复系组配的杂交种F1穗柄下茎秆高度变平均值01-26A比7050A降低19.9%,差异达极显著水平,介于双亲之间。说明穗柄下茎秆高度在01-26A矮化株高效应中发挥着重要作用。在01-26A为母本时,除01-26A/NK1外,其他5个粒用高粱组合F1杂交组合穗柄下茎秆高度均介于双亲之间并且高粱两亲本的平均值;7050A与所有粒用组合杂种F1穗柄下茎秆高度均高于亲本最高值,与株高遗传趋势相一致。在与甜高粱杂交F1中,01-26A和7050A与甜高粱恢复系杂种F1穗柄下茎秆高度差异不显著,其变化趋势与株高遗传基本一致,进一步说明了穗柄下茎秆高度对株高变化的重要作用。
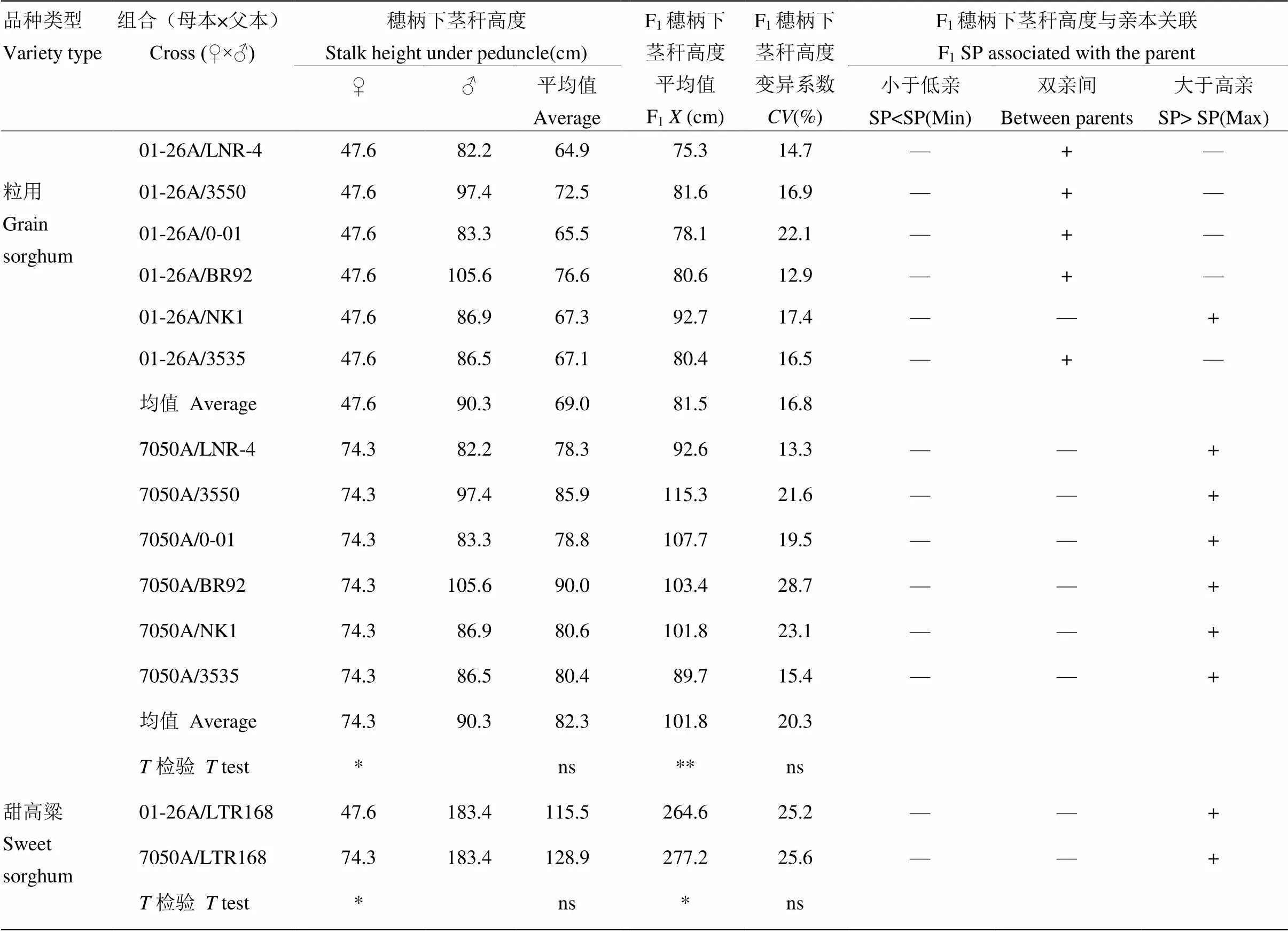
表4 不育系、恢复系及其F1穗柄下茎秆高遗传效应
+:穗柄下茎秆高度在这一范围;—:穗柄下茎秆高度不在这一范围;SP:穗柄下茎秆高度
+: the stalk height under the peduncle is in this range; —: The stalk height under the peduncle is not in this range; SP: the stalk height under the peduncle
2.1.3 穗柄长遗传分析 穗柄长作为株高的重要组成部分,在01-26A和7050A与粒用高粱恢复系组配的F1杂交种中变化活跃,01-26A作母本组配的F1穗柄长显著低于7050A(表5)。01-26A和7050A 2个母本差异不显著,而在与相同父本杂交F1中,01-26A作母本与粒用高粱组配的F1穗柄长比7050A的F1降低10.7%,且变异系数较小。说明穗柄长在01-26A矮化株高效应中发挥着重要作用,且效应值与株高相近。此外,在01-26A为母本时,与6个粒用高粱恢复系杂种F1组合穗柄下茎秆高度均高于亲本最高值,其遗传变化大于株高;7050A与所有粒用组合杂种F1穗柄下茎秆高度均高于亲本最高值,与株高遗传趋势相一致。另外,01-26A和7050A与甜高粱杂交F1中穗柄长差异不显著,可能是因为这两个不育系存在相同或相似的调控穗柄长度的基因。
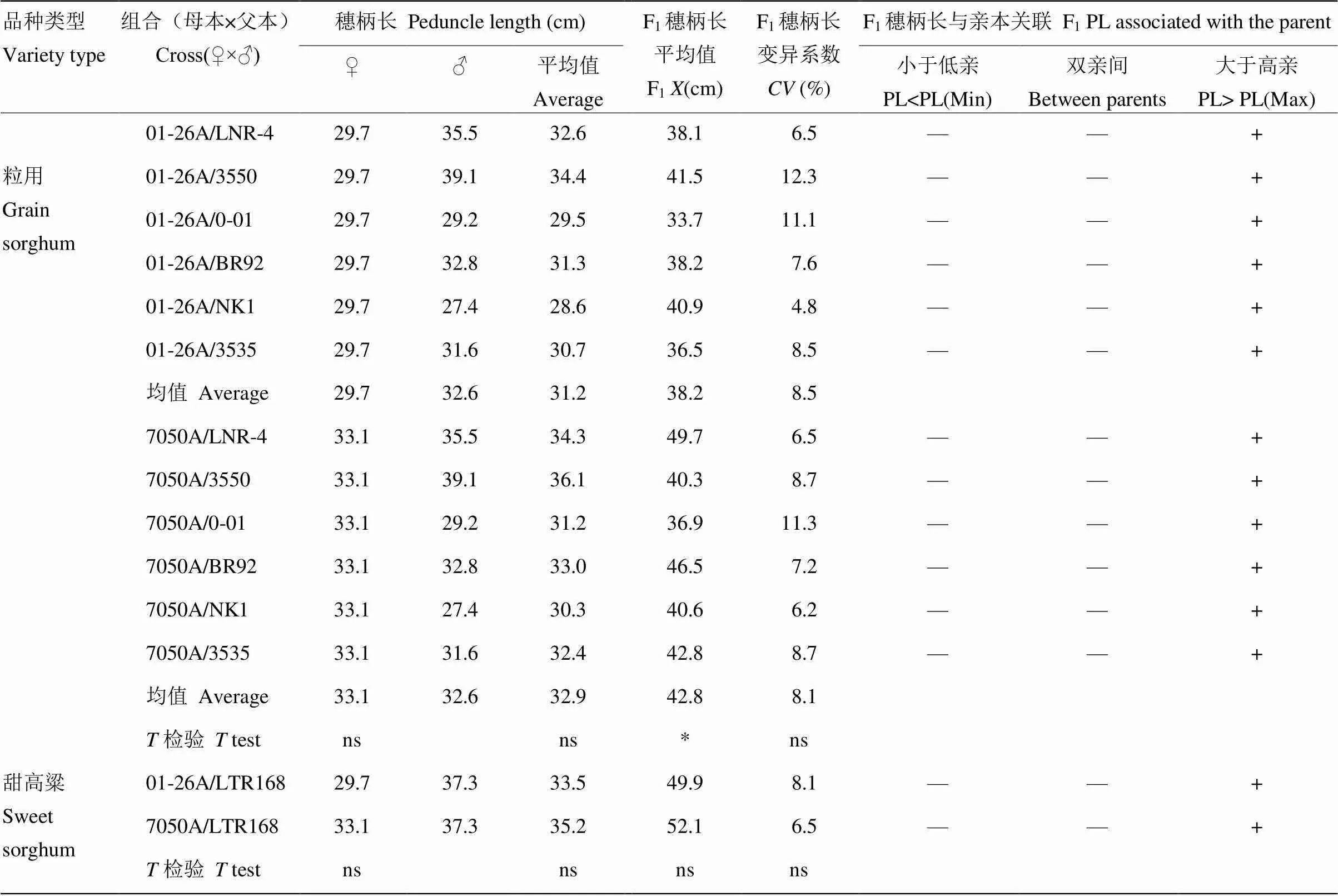
表5 不育系、恢复系及其F1穗柄长遗传效应
+:穗柄长在这一范围;—:穗柄长不在这一范围;PL:穗柄长
+: the peduncle length is in this range; —: the peduncle length is not in this range; PL: peduncle length
2.1.4 穗长遗传分析 01-26A和7050A与粒用高粱组配的F1穗长差异不显著(表6)。2个母本01-26A和7050A穗长差异不显著,父本相同,01-26A和7050A分别与6个粒用高粱恢复系组配杂交种F1中,01-26A作母本F1的穗长变幅为28.7—33.4 cm,平均变异系数()仅为3.2%,而7050A作母本的F1穗长变幅为26.7—32.2 cm,平均变异系数()为4.5%,说明01-26A作母本组配的杂种F1穗长较7050A相对更加整齐,但穗长的变化较穗柄下茎秆高度和穗柄长对杂种F1株高影响相对较小。
2.1.5 茎节参数对株高遗传的影响 不育系01-26A和7050A与粒用高粱组配的F1的节间长、茎节数与株高存在相关(表7)。分析发现,01-26A组配的多数F1中下层和中层节间长度与株高变化相关性较大,而与7050A组配的的F1则表现为中上层和上层节间长度与株高关联密切。所以,分析认为01-26A杂交组配的F1株高的矮化效应很可能是由于茎秆中下层和中层节间长度变短所致;而对于甜高粱,01-26A和7050A组配的F1均表现为中层和中上层节间长度与株高相关性较大,也和上述结果相吻合。此外,2个母本无论是与粒用高粱还是甜高粱恢复系组配的杂种F1均表现出节数与株高关联密切。

表6 不育系、恢复系及其F1穗长遗传效应
+:穗长在这一范围;—:穗长不在这一范围;HL:穗长 +: the head length is in this range; —: the head length is not in this range; HL: head length

表7 茎节相关参数对杂种F1株高遗传的影响
下层:下2节;中下层:下6节;中层:下10节;中上层:下14节;上层:下18节
lower: No.2 internode; lower middle: No.6 internode; middle: No.10 internode; upper middle: No.14 internode; upper: No.18 internode
2.2 组配F1杂交种株高矮化效应验证
为进一步验证01-26A的株高矮化效应,对01-26A与33个恢复系、7050A与24个恢复系的杂种F1亲本的株高进行了测定与分析(图1)。通过F1株高分类和出现频次分析发现,01-26A组配的杂种F1株高在100—160 cm,且多数品种株高在140—160 cm;而与7050A组配的杂种F1株高在140—190 cm,多数品种株高集中在160—190 cm。对这两个不育系与13个甜高粱恢复系组配分析发现01-26A与甜高粱组配的杂种F1株高主要都集中在340—380 cm,7050A与甜高粱组配的杂种F1代株高主要都集中在360—400 cm,与先前(2.1.1)的研究结果相吻合。通过验证进一步说明01-26A可显著降低杂种F1粒用高粱的株高,但对甜高粱没有矮化株高的效应。

图1 组配F1杂交种株高矮化效应验证
2.3 株高矮化基因位点研究
2.3.1 高粱Dw序列分析 为进一步分析01-26A对粒用高粱杂种F1株高的矮化效应和对甜高粱杂种F1株高无矮化效应的作用机制,对试验材料进行了矮化基因检测。分析发现雄性不育系01-26A和7050A、7个恢复系(6个粒用、1个甜高粱)以及它们组配的F1杂交种在Dw位点上存在差异(图2)。01-26A作为母本,在Dw基因组序列中第1 350位上碱基由A突变为T,造成Dw发生突变,即01-26A具有矮化基因dwdw,而与之组配的6个粒用高粱恢复系(LNR-4、NK1、0-01、3550、3535、BR92)除3550基因型发生突变(由A到T)为dw外(01-26A与3550杂种F1基因型dwdw,表型矮秆),其他5个品系基因型均为DwDw,与之组配的杂交种基因型均为杂合,即Dwdw,由于在杂种F1高粱高秆对矮秆表现为部分显性[12],因此,01-26A对F1的株高矮化起到了一定的作用,此结果也与株高表型(2.1.1和2.2)的研究结果相呼应。01-26A与甜高粱恢复系杂交F1基因型为Dwdw,株高表现为小幅下降。7050A作为母本,在基因组第1 350位上碱基未发生突变,所以其基因型为DwDw,不具矮化基因,除与3535杂种F1基因型Dwdw外,与其他恢复系(包括粒用和甜高粱)杂交F1的基因型均为DwDw,所组配组合表现为株高普遍高于01-26A组配的杂交组合,与表型结果相吻合。
2.3.2 高粱Dw序列分析 由图3可以看出,在Dw基因组第549位上,01-26A碱基序列GA并未缺失(GA缺失为dw突变的特征位点),说明01-26A基因型为DwDw,而其他所有参试恢复系组配F1杂交种(粒用和甜高粱)除恢复系0-01发生基因缺失杂合(导致杂种F101-26A/0-01为Dwdw外,其他F1杂交种基因型均为DwDw。7050A在Dw位点的基因型与01-26A相同(DwDw),其组配的F1杂交种与01-26A一致,所以Dw在这两个不育系杂交F1组合株高矮化调控中没有起到明显作用。

图2 高粱Dw1位点分析

**:基因缺失;··:杂合 **: gene deletion; ··: heterozygosity
2.3.3 高粱Dw位点分析 矮化基因Dw在第五外显子插入882 bp的重复序列,使其变成dw,基因片段大小由1 450 bp变为2 330 bp。由图4、表8可以看出,01-26A(第17泳道)和7050A(第8泳道)在Dw位点基因型均为dwdw,而参试恢复系LNR-4、3550和BR92为野生型(DwDw),其他恢复系均为突变型(dwdw)。从Dw位点角度分析发现01-26A与粒用杂交组合01-26A/3550、01-26A/BR92、01-26A/LNR-4及甜高粱杂交组合01-26A/LTR168基因型均为Dwdw,此研究结果与先前(2.1.1和2.2)株高表型的研究结果基本吻合;而7050A与粒用杂交组合7050A/NK1、7050A/3535和7050A/3550基因型均为dwdw,其他组合基因型为Dwdw,也与基因株高表型变化(2.1.1和2.2)基本一致。综上所述,01-26A的dwdw在对杂种F1株高矮化调控中发挥了重要作用。而01-26A和7050A与甜高粱恢复系杂交组配均为Dwdw,也进一步解释了2个母本组配的甜高粱杂交种均表现为高秆的内在原因。
2.3.4 矮化基因型及平均株高 对参试的2个雄性不育系、7个恢复系和其组配形成的14个杂交种的基因型和平均株高进行分析(表9)。发现01-26A和7050A的基因型相同,均为DwDw;01-26A的和矮化基因型分别为dwdw和dwdw。而7050A的Dw基因型为DwDw,基因型为dwdw,其组配的粒用杂交组合的平均株高总体高于01-26A组配的杂交种。此外,尚未被克隆,没有检测,但其基因型很可能也与F1株高密切相关。
3 讨论
株高作为高粱群体构成和机械化生产的重要指标备受人们的关注,而通过遗传手段矮化株高是调控株高最有效、最稳定的手段之一[16]。本研究通过将01-26A与粒用高粱和甜高粱恢复系杂交,对亲本和组配F1杂交种的株高进行分析,发现该材料可显著降低粒用杂种F1的株高,而对甜高粱杂种F1株高没有显著的矮化效应。此研究结果与管延安等[17]对普通高粱与甜高粱杂交组合株高主基因多基因模型遗传效应研究得出的株高遗传变化趋势基本一致,李延玲等[18]对高粱株型性状数量遗传分析也得出类似的结论。此外,Olson等[19]研究认为,株高是高粱株型调节的关键农艺性状,通过遗传改良株高是高粱矮化育种的必然方向,与本研究结果相吻合。

表8 Dw3检测序号及对应品系/组合

表9 矮化基因型及其平均株高
因为w4尚未被克隆,没有检测,故表中未作标记 Becausew4has not been cloned yet, there is no detection, so the table is not marked in table

1、2、3、7、9、10、12、13、14、20、21和23泳道片段大小为1263 bp,其他片段大小为2145 bp
本研究通过遗传分析认为01-26A矮化株高遗传效应主要表现在杂种F1代穗柄下茎秆高度明显降低,同时穗柄长降低也是造成株高变矮的重要原因,穗长对株高变化的影响很小。此研究结果与MULTANI[20]对高粱株高调控的研究结果相一致;Ordonio等[21]以粒用高粱对高粱叶夹角、株高、穗长、平均茎节长度遗传分析也得出类似的结论。有研究认为,小麦株高与其构成因素呈极显著遗传正相关,株高构成因素对株高的作用大小依次为倒一节>倒二节>倒三节>倒五节>倒四节>穗的结论[22],此结果与本研究结果基本吻合,但影响的节间位置略有差异,可能是由于作物不同,植株高度差异和分蘖数不同所致。
前人已经有过报道,高粱株高的调控基因主要有、、和,株高矮化是这4个基因以及其他未知基因共同作用的结果[23-25]。虽然本研究中通过对01-26A矮化基因型的检测确定其基因型为Dwdw,对没有检测,但鉴于其与30余个恢复系杂交F1均为株高降低的表型数据分析,尤其是穗柄长度的分析(较7050A组配的杂交组合显著降低),可推断01-26A位点很可能为,即01-26A的基因型为dwDwDwdwdwdwdw,即3-矮高粱雄性不育系。而7050A很可能是DwDwDwdwdwdwdw的2-矮不育系。此推断与前人通过隐性等位基因在Milo系中鉴定出和,而在Kafir背景中鉴定了的隐性等位基因的论断基本一致[26-28]。本研究还发现,01-26A的和在其杂种F1株高矮化调控方面发挥了重要作用,可促使茎秆中下部节间变短从而降低株高。此结果与Jia等[29]研究并提出的和具有降低株高的效应,但作用效应值更大的结果相吻合;但与THURBER等[30]研究并指出的在高粱株高调控中起着关键作用,中部茎秆影响程度最大,茎秆受影响程度较大的结果略有差异,可能是选用矮化材料差异所致,也可能是几个基因互作差异所致。截至目前,虽然还没有被克隆,报道也很少,但普遍认为影响高粱穗柄长[31]。
控制高粱株高的基因除Dw—Dw外,还可能受其他基因或其他内在因素的调控,尚需进一步研究;同时本研究采用的高粱恢复系类型有限,通过01-26A与更多矮化基因型和更多种类恢复系杂交组配进行株高矮化效应分析还有待于进一步研究。
4 结论
高粱雄性不育系01-26A确定含有dwDwDwdwdw3对基因,同时通过其大量株高矮化效应验证,推断其很可能具有,即为基因型dwDwDwdwdwdwdw的三矮高粱不育系。01-26A具有和大多数粒用高粱恢复系组配都能矮化杂种F1株高的遗传效应,可主要通过降低杂种F1中下部节间长度和穗柄长度降低F1株高,实现对高粱株高的矮化调控,但其与甜高粱恢复系杂交未发现其具有明显矮化效应。
[1] MORRIS G P,RAMU P, DESHPANDE S P, HashCT, ShahT, UpadhyayaHD. Population genomic and genome-wide association studies of agroclimatic traits in sorghum., 2013, 110(2): 453-458.
[2] 王劲松, 焦晓燕, 董二伟, 王彬, 武爱莲, 韩雄, 郭珺, 王立革. 不同灌溉时期和施氮量对高粱水分利用及产量的影响. 山西农业科学, 2016, 44(6): 777-783.
Wang J s, Jiao X y, Dong E W, Wang B, Wu A l, Han X, Guo J, Wang L g. Effects of different irrigation periods and nitrogen application rates on sorghum water use and yield ., 2016, 44(6): 777-783. (in Chinese)
[3] 王平, 丛玲, 朱振兴, 张丽霞, 张曦. 高粱化基因Dw/dw对株高及其它农艺性状的影响. 辽宁农业科学, 2019(5): 12-15.
Wang P, Cong L, Zhu Z x, Zhang L x, Zhang X. Effects of sorghum dwarfing geneDw/dwon plant height and other agronomic traits., 2019(5): 12-15. (in Chinese)
[4] 李嵩博, 唐朝臣, 陈峰, 谢光辉. 中国粒用高粱改良品种的产量和品质性状时空变化. 中国农业科学, 2018, 51(2): 246-256.
Li S b, Tang C c, Chen F, Xie G h. The spatio-temporal changes of yield and quality traits of improved grain sorghum varieties in China., 2018, 51(2): 246-256. (in Chinese)
[5] 杨慧勇, 赵文博, 王花云, 张福耀. 高粱丝黑穗病菌4号生理小种抗性基因的定位. 中国农业科学, 2015, 48(8): 1484-1491.
Yang H y, Zhao W b, Wang H y, Zhang F y. Location of the resistance gene of sorghum silk smut No. 4 physiological race., 2015, 48(8): 1484-1491. (in Chinese)
[6] 肖继兵, 刘志, 孔凡信, 辛宗绪, 吴宏生. 种植方式和密度对高粱群体结构和产量的影响. 中国农业科学, 2018, 51(22): 4264-4276.
Xiao J b, Liu Z, Kong F x, Xin Z x, Wu H s. Effects of planting methods and density on sorghum population structure and yield., 2018, 51(22): 4264-4276. (in Chinese)
[7] HIGGINS R H, THURBER C S, ASSARANURAK I, BROWN P J. Multiparental mapping of plant height and flowering time QTL in partially isogenic sorghum families., 2014, 4(9): 1593-602.
[8] 黄瑞冬, 高悦, 周宇飞, 吴奇, 张姣, 尚培培, 张壮, 高铭悦, 韩熠, 许文娟. 矮秆高粱辽杂35光合特性与产量构成因素. 中国农业科学, 2017, 50(5): 822-829.
Huang R d, Gao Y, Zhou Y f, Wu Q, Zhang J, Shang P p, Zhang Z, Gao M y, Han Y, Xu W j. Photosynthetic characteristics and yield components of dwarf sorghum Liaoza 35., 2017, 50(5): 822-829. (in Chinese)
[9] 王劲松, 焦晓燕, 丁玉川, 董二伟, 白文斌, 王立革, 武爱莲. 粒用高粱养分吸收、产量及品质对氮磷钾营养的响应. 作物学报, 2015, 41(8): 1269-1278.
Wang J s, Jiao X y, Ding Y c, Dong E w, Bai W b, Wang L g, Wu A l. Response of nutrient absorption, yield and quality of grain sorghum to nitrogen, phosphorus and potassium nutrition. ., 2015, 41(8): 1269-1278. (in Chinese)
[10] MURPHY R L, KLEIN R R, MORISHIGE D T, BRADY J A, ROONEY W L, MILLER F R. Coincident light and clock regulation of pseudo response regulator protein 37 (PRR37) controls photoperiodic flowering in sorghum., 2011, 108(39): 16469-16474.
[11] ROSS W M, KOFOID K D, Identification of granules for harvesting with Dw1 dominant gene., 1979, 19(3): 339-344.
[12] 卢庆善, 孙毅. 杂交高粱遗传改良. 北京: 中国农业科学技术出版社, 2005: 37-38.
LU Q S, SUN Y.. Beijing: China Agricultural Science and Technology Press, 2005: 37-38. (in Chinese)
[13] YAMAGUCHI M, FUJIMOTO H, HIRANO K, ARAKI-NAKAMURAS, OHMAE-SHINOHARA K, FUJII A. SorghumDw, an agronomically important gene for lodging resistance, encodes a novel protein involved in cell proliferation., 2016, 6(1): 28366.
[14] LI X, LI X, Fridman E, Tesso T T, Yu J. Dissecting repulsion linkage in the dwarfing geneDwregion for sorghum plant height provides insights into heterosis., 2015, 112(38): 11823.
[15] BROWN P, ROONEY W L, FRANKS C, KRESOVICH S. Efficient mapping of plant height quantitative trait loci in a sorghum association population with introgressed dwarfing genes., 2008, 18(1): 629-637.
[16] 李欣禹, 罗峰, 刘惠芬, 裴忠有, 高建明, 姜亦巍, 孙守钧. 不同类型高粱与环境的相互作用关系. 江苏农业科学, 2017, 45(4): 59-63.
LI X Y, LUO F, LIU H F, PEI Z Y, GAO J M, JIANG Y W, Sun S Q. The interaction between different types of sorghum and the environment., 2017, 45(4): 59-63. (in Chinese)
[17] 管延安, 张华文, 樊庆琪, 杨延兵, 秦岭, 王海莲, 王洪刚. 普通高粱与甜高粱杂交组合株高、糖度的主基因多基因模型遗传效应分析. 核农学报, 2012, 26 (1): 36-42.
GUAN Y A, ZHANG H W, FAN Q Q, YANG Y B, QIN L, WANG H L, WANG H G. Genetic effects analysis of the main gene polygenic model of plant height and sugar content of common sorghum and sweet sorghum hybrids., 2012, 26(1): 36-42. (in Chinese)
[18] 李延玲, 白晓倩, 于澎湃, 高建明, 裴忠有, 罗峰, 孙守钧. 高粱株型性状数量遗传分析. 华北农学报, 2018, 33(1): 143-149.
LI Y L, BAI X Q, YU P P, GAO J M, PEI Z Y, LUO F, SUN S J. Quantitative genetic analysis of plant type traits in sorghum., 2018, 33(1): 143-149. (in Chinese)
[19] OLSON S N, RITTER K, ROONEY W, KEMANIAN A, MCCARL B A, ZHANG Y. High biomass yield energy sorghum: Developing a genetic model for C4 grass bioenergy crops., 2012(1): 640-655.
[20] MULTANI D S. Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants., 2003, 302(5642): 81-84.
[21] ORDONIO R L, ITO Y, HATAKEYAMA A, OHMAE-SHINOHARA K, KASUGA S, TOKUNAGA T. Gibberellin deficiency pleiotropically induces culm bending in sorghum: an insight into sorghum semi-dwarf breeding., 2014, 4(1): 5287.
[22] 姚金保, 任丽娟, 张平平, 杨学明, 马鸿翔, 姚国才, 张鹏, 周淼平. 小麦株高及其构成因素的遗传及相关性分析. 麦类作物学报, 2011, 31(4): 604-610.
YAO J B, REN L J, ZHANG P P, YANG X M, MA H X, YAO G C, ZHANG P, ZHOU M P. Analysis of genetics and correlation of plant height and its component factors in wheat., 2011, 31(4): 604-610. (in Chinese)
[23] MIZUNO H, KAWAHIGASHI H, KAWAHARA Y, KANAMORI H, MATSUMOTO T. Global transcriptome analysis reveals distinct expression among duplicated genes during sorghum-interaction., 2012, 12(1): 121.
[24] BROWN P J, PATERSON A H. Bridging classical and molecular genetics of sorghum plant stature and maturity., 2013, 11(1): 333-345.
[25] KLEIN R R, MULLET J E, JORDAN D R, MILLER F R, ROONEY W L, MENZ M A. The effect of tropical sorghum conversion and inbred development on genome diversity as revealed by high resolution genotyping., 2008, 48(1): 12-26.
[26] CASSADY A J, Klein P E, HILLEY J L, SAHRAEIAN S M E, SHARMA A, MULLET J E.Effect of a single height () gene of sorghum on grain yield, grain yield components, and test weight., 1965, 5(5): 385-388.
[27] MORISHIGE D T, KLEIN P E, HILLEY J L, SAHRAEIAN S M E, SHARMA A, MULLET J E. Digital genotyping of sorghum-A diverse plant species with a large repeat-rich genome., 2013,14 (1): 448.
[28] WU E, LENDERTS B, GLASSMAN K, BEREZOWSKA-KANIEWSKA M, ZHAO Z Y. Optimized-mediated sorghum transformation protocol and molecular data of transgenic sorghum plants., 2014, 50(1): 9-18.
[29] JIA Q, ZHANG J, WESTCOTT S, ZHANG X Q, BELLGARD M, LANCE R. GA-20 oxidase as a candidate for the semi dwarf gene sdw1 denso in barley., 2009, 9(1): 255-262.
[30] THURBER C S, MA J M, HIGGINS R H, BROWN P J. Retrospective genomic analysis of sorghum adaptation to temperate- zone grain production., 2013, 14(6): 68.
[31] YANG S, WEERS B D, MORISHIGE D T, MULLET J E. CONSTANS is a photoperiod regulated activator of flowering in sorghum., 2014, 14 (1): 148.
Dwarfing effect and molecular mechanism of an elite sorghum Male sterile line 01-26A in its hybrids
ZOU Jianqiu, WANG Yanqiu, LI Jinhong, ZHU Kai
(Sorghum research institute, Liaoning Academy of Agricultural Sciences, Shenyang 110161)
【】The mechanized production of sorghum is the inevitable direction of future development, and the ideal plant type is the basis and key factor for mechanized production. The dwarf male sterile line 01-26A was found a unique effect in reducing the plant height in F1generation when crossed with available grain sorghum restorer lines. Therefore, a study was conducted to determine the genetic mechanism and regulation mechanism of plant height dwarfing by its genetic effects and regulatory gene loci. 【】This study used sorghum male sterile line 01-26A (A1cytoplasm) with dwarf plant height effect and sorghum male sterile line 7050A (A2cytoplasm) without dwarf plant height effect as the test material, focusing on 7 restorer lines, including 6 grain restorer lines and 1 sweet sorghum restorer line, and their cross F1generation hybrids, the genetic effects of plant height, number of nodes, the total internode length under peduncle, the peduncle length and the head length were analyzed, the gene loci oftorelated with the plant height were also measured and analyzed.was not included because it had not been cloned.【】the male sterile line 01-26A had a significant dwarf effect on the plant height of grain sorghum, and its hybrids is 15.8% lower than that of the 7050A, generally, the absolute value of plant height did not exceed 160 cm. The plant height of F1generation derived from 01-26A with sweet sorghum restorer line had not been obviously reduced, so it had not dwarf effect on sweet sorghum hybrid. The genetic dwarf effect of 01-26A was mainly manifested in the shortening of the internode length under peduncle, and the internode length under peduncle has more correlated with the plant height variation. The peduncle length reduction of F1crossed by 01-26A with restorer lines is another reason of plant becoming shorter, but the effect was less than that of the internode length under peduncle. While the head length had much little effect on plant height variation. The dwarf genotype (to) of 01-26A was determined by PCR and sequencing ofDwtoDwgenes. And combined the analysis of plant height genetic data of multiple cross combinations, the plant height genotype of 01-26A was deduced to bedwdwDwDwdwdwdwdw,a 3-dwarf sorghum sterile line. In addition, by the analysis of plant height regulation genes, we found that theandof 01-26A may had a greater effect on the plant height of grain sorghum, while the presence of DW2was the immanent cause for not dwarfing on sweet sorghum F1.【】 01-26A was likely to be a 3-dwarf sorghum male sterile line with the genotype ofdwDwDwdwdwdwdw. It could achieve dwarf regulation of its F1by reducing the internode length (main effect) and the peduncle length (secondary effect). However, 01-26A, had not been found obvious dwarfing effect when crossed with sweet sorghum, which may be due to the presence of.
sorghum; plant height; genetic effect; gene locus

10.3864/j.issn.0578-1752.2020.14.006
2019-07-31;
2019-09-22
国家重点研发计划(2019YFD1001704/2019YFD1001700)、国家现代农业产业技术体系(CARS-06-13.5-A11)、辽宁省中央引导地方项目(2018416023)
邹剑秋,E-mail:jianqiuzou@126.com。通信作者朱凯,E-mail:zhukai72@163.com
(责任编辑 李莉)
