环境因素对胶州湾栽培龙须菜的影响及生长预测模型构建❋
2020-08-03张敬宇隋正红
张敬宇, 周 伟, 郭 鹏, 隋正红❋❋
(1. 中国海洋大学海洋生物遗传学与育种教育部重点实验室, 山东 青岛 266003; 2. 江苏海洋大学,江苏省海洋生物资源与环境重点实验室, 江苏 连云港 222005;3. 江苏海洋大学,江苏省海洋生物产业技术协同创新中心, 江苏 连云港 222005; 4. 江苏海洋大学,江苏省海洋生物技术重点实验室, 江苏 连云港 222005)
龙须菜属于红藻门(Rhodophyta)、红藻纲(Rhodophyceae)、真红藻亚纲(Florideae)、杉藻目(Gigartinales)、江蓠科(Gracilariaceae)。它的生长速度快,琼胶含量高,养殖过程中还能吸收N、P等富营养元素,解决海水的富营养化现象[1]。2018年江蓠养殖产量位列中国海藻产量第二,仅次于海带(Saccharinajaponica)[2]。龙须菜作为江蓠栽培产业化中最成功的物种,是重要的产琼胶海藻[3]。福建、山东、广东作为江蓠养殖大省,占全国养殖面积95%以上[2]。山东作为养殖三大省中的唯一北方省份,在龙须菜南北轮养模式中起到了重要的作用。山东栽培海域覆盖黄海和渤海,青岛胶州湾频临黄海,且作为山东省龙须菜的栽培区具有较悠久的历史,本次实验选择青岛胶州湾常规栽培海域作为实验位点。
近年来越来越多的研究开始关注龙须菜与环境因素之间的关系。Li等[4]利用筏式栽培技术在青岛海域对野生龙须菜进行实地栽培实验,全年跟踪调查藻体生物量及海水温度变化,阐述了龙须菜生物量随季节发生变化。Yang等[5]在青岛胶州湾展开了相应的龙须菜栽培优化实验,重点比较了不同栽培方式(水平挂养和垂直挂养)及栽培水层对龙须菜生长的影响,发现水平挂养是最为优异的栽培方式,垂直挂养中水层对龙须菜生长影响较为明显。Zhou等[6]建立了龙须菜与黑鮶(Sebastodesfuscescens)共养体系,对5组处理体系中营养盐的变化进行定时检测,数据显示藻体生长吸收大量营养盐,生物量的变化与营养盐之间是存在关系的。陈伟洲等[7]利用07-1、07-2和981三种品系进行海区实地栽培,采集了6周的实验数据,初步研究显示龙须菜生物量与海水温度、透明度呈正相关性,与海水盐度呈现一定负相关性。状东红等[8]研究了龙须菜养殖与广东深澳湾环境因子的变化特征,结果显示龙须菜养殖能够有效的吸收海水中的氨态氮和无机磷。
目前有关龙须菜海区栽培的影响因素研究相对较少,少数研究虽然关注了环境因素的影响,但取样时间点较少、未能系统的针对龙须菜生长进行分析。实验室环境与海区实际栽培之间存在较大的差异,在实验室培养过程中,龙须菜的生长环境较为稳定,无法真实地反映实际生产中环境因素与龙须菜生长状况之间的关系。传统的农业养殖中,根据养殖户的养殖经验进行粗放式的管理,夹苗时间单一,水层固定等,忽略了不同季节环境因子,如温度和光照的剧烈变化及耦合作用等对龙须菜生长的影响。因此,及时掌握龙须菜挂养层海水温度、光照、盐度、 pH 及营养盐等生态因子的变化规律,研究海区栽培状态下龙须菜生长与生态因子变化的响应规律,显得尤为重要。
本研究采用养殖最常用的方法,即通过藻段夹苗、水平挂养,采用“全浮筏式”栽培,对栽培状态下两种龙须菜品系鲁龙1号和981的生长与生态因子进行实时监测,构建龙须菜生态动力学模型。研究结果对于认识自然海区栽培龙须菜的生长对生态因子的响应规律具有重要意义,并可进一步为龙须菜栽培实践和管理提供科学依据。
1 材料方法
1.1 研究位点及材料处理
本次研究位点胶州湾位于中国黄海中部,面积近400 km2(见图1)。
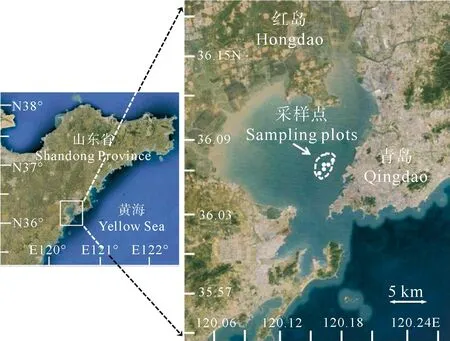
(白色曲线框内显示为栽培区域;白色箭头指示为采样点。The cultivation area is shown in the white curve box; sampling locations are shown by the white arrow.)
2015 年5~11月,在青岛胶州湾龙须菜栽培区设实验站点(35°05′N~36°07′N,120°16′E~120°18′E)。站点内所设龙须菜栽培架为单筏式全浮筏架,全长100 m,双架间距6 m,架身每隔5 m悬黑色可再生浮球1个(或泡沫浮漂),浮球漂浮在水体表面,距离主筏架20~30 cm,筏架方向与水流方向较为一致,生态环境优良,环境较为稳定。
实验用龙须菜“鲁龙1号”来自福建省连江罗源湾金牌渔业科技有限公司, 实验用龙须菜“981”来自福建省莆田市南日岛栽培区,两种品系的龙须菜均为栽培常用的二倍体。实地夹苗,夹苗用苗绳为聚乙烯材质,单绳长6 m两端预留各0.5 m,有效夹苗长度为5 m,夹苗起始量为每根苗绳2 kg龙须菜。夹苗后均匀分至各站点,水平挂养至筏架上,控制苗绳距离一致(距离水面约0.5 m)。在苗绳重量大于15 kg时进行分苗,防止苗绳断裂。
1.2 藻体生物量及日平均生长速率测定
采用便携式电子称(Model:WH-A03,China),称量空苗绳重量及起始夹苗后绳苗重量,二者差值为实际起始夹苗量W0;每站点每2周(t)测定10根单苗绳上藻体重量(去苗绳重),记录为Wt,直至实验结束。根据以下公式,计算龙须菜日平均生长速率(Specific growth rate, SGR)[13]:
SGR(%·d-1)=(lnWt-lnW0)×t-1×100% 。
其中:W0起始夹苗鲜重;Wt培养t天后藻体鲜重;t培养时间。
1.3 环境因子监测
环境因子的监测与海区栽培实验同步进行,以保证龙须菜生物量的变化情况与环境因素之间一一对应。具体操作如下:
温度和光照:温光测定采用高精度HOBO系列光照温度记录仪(Model: UA-002-64/Onset, USA),实验期间每30 min采集一次数据,实验结束后统一导出数据。
盐度:海水相对盐度测定采用手持式折光仪(LS10T, Suwei, China)测定,每2周测定1次,重复3次。
pH:海水pH测定采用pH计(STARTER 3100, OHAUS, China),每2周测定1次,重复3次。
营养盐:在龙须菜栽培水层,每站点每2周采集3次水样。采集完成后,经0.22 μm滤膜过滤后,即刻测定NH3-N、NO3-N、PO4-P和DIC(海水溶解无机碳)浓度,具体流程参考“Palintest Test Instructions”(YSI,USA):利用YPM152和YAT1707配制NH4-N测量液;利用YAP163配制NO3-N测量液;利用YAP177配制PO4-P测量液;利用YAP188配制总碱度测量液(需结合测定pH,计算DIC)[11],采用YSI便携式分光光度计(YSI 9500, USA)测定上述测量液,记录浓度读数即可,表示单位为mg·L-1。
1.4 显微观察方法
实验数据采集时,收集少量材料(<5 g)于采样袋中带回观察。使用双目体式显微镜(SZ2-ILST, OLYMPUS, Japan)观测龙须菜的表皮形态。使用手术刀将龙须菜表皮解剖下来,置于载玻片上,滴加少量海水后用盖玻片轻压封片置于正置显微镜(BX53, OLYMPUS, Japan)中观察龙须菜表皮的显微形态。
1.5 数据统计及处理
数据分析利用软件SPSS 21.0(IBM,USA),分析中显著性水平p<0.05。栽培龙须菜藻体生长速率与生态因子相关性分析采用Pearson相关性分析;对模型整体的评价采用F检验;回归模型的系数采用t检验;对模型贡献程度采用Shapley值法进行计算;模型构建过程中产生的数据只保留小数点后三位。
本研究中,海水温度与光照强度是指HOBO仪器每隔30 min记录的海水的实际温度与实测的光照强度。其中,日均温度的计算过程如下:
式中,T1至T48指24 h记录的48组数据。以本公式计算出的Taver指代日均气温。
文中分析使用的光照强度值是由HOBO仪器所记录的一天之内48组光照强度的平均值,以此指代这一天的光照强度。文中光周期用来表示光照时长,计算方法是HOBO仪器所记录的光强第一个非零值到记录的最后一个非零值之间的时长。
文中所用的DIC(海水溶解无机碳)是根据总碱度与pH值计算而来,计算过程如下:

2 结果
2.1 龙须菜生物量及生长速率的动态变化
胶州湾龙须菜的栽培持续近6个月,整个栽培季共分苗2次,分别在7月1日和28日(见图2)。
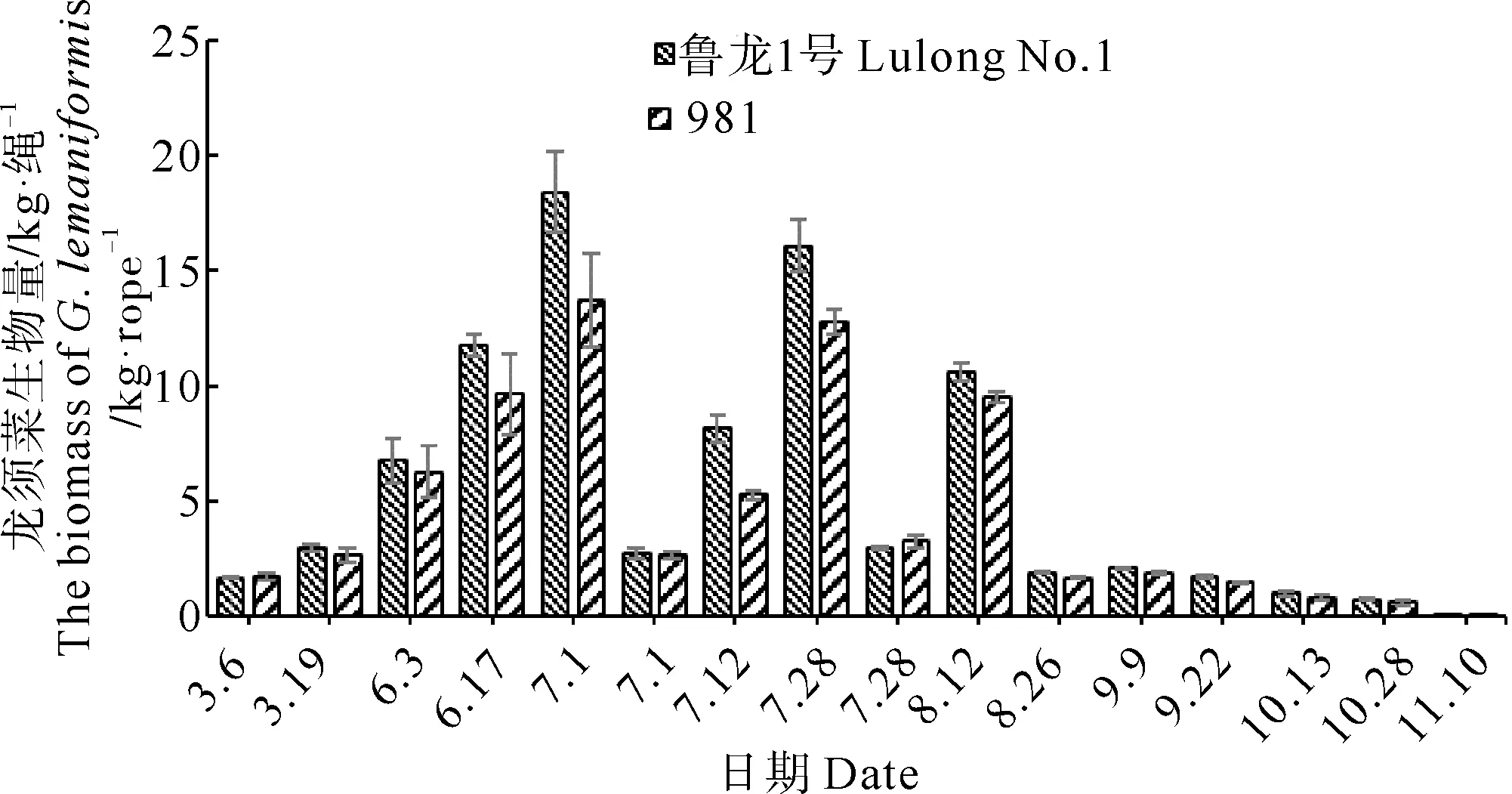
图2 胶州湾栽培季节龙须菜单绳生物量变化
本实验开始至第一次分苗共计8周,在此期间龙须菜生长速率整体较快。夹苗起始量分别为(1.67±0.06) kg FW/绳(鲁龙1号)和(1.72±0.18) kg FW/绳(981),二者无明显差异(p>0.05);至7月1日龙须菜鲁龙1号和981生物量分别达到(18.41±2.75) kg FW/绳和(13.71±3.02) kg FW/绳。收获龙须菜鲁龙1号的生物量显著高于981(p<0.05)。将龙须菜收获后重新夹苗,起始量分别为(2.73±0.25) kg FW/绳(鲁龙1号)和(2.63±0.16) kg FW/绳(981)。自夹苗后,龙须菜生物量累计更加迅速,经4周实地栽培龙须菜生物量达到(16.07±1.10) kg FW/绳(鲁龙1号)和(12.77±0.55) kg FW/绳(981),均显著高于起始夹苗量;收获龙须菜鲁龙1号生物量也明显高于龙须菜981(p<0.05)。7月28日分苗后,夹苗起始量分别为(2.95±0.05) kg FW/绳(鲁龙1号)和(3.23±0.25) kg FW/绳(981)。在此阶段,龙须菜生物量经历短暂的高速累积后迅速下降,之后一直维持在较低的水平。
在胶州湾不同栽培时期中,龙须菜生长速率表现出显著差异(p<0.05)(见图3)。5月18日~8月12日期间,龙须菜长势较好。初期长势总体较为平稳,在5%左右浮动。7月1日~7月12日期间,生长速率达到最高值,其中鲁龙1号的生长速率为(9.12±0.65)%·d-1,981的生长速率为(5.79±0.34)%·d-1。8月26日之后,龙须菜生长速率呈负增长,分别在8月26日和11月10日达到最低点。
不同品种(鲁龙1号和981)在整个阶段的生长速率变化趋势相一致,表现为“同正或同负”。具体到每次采样,鲁龙1号的生长速率较981具有优势;特别在7月份,鲁龙1号的生长速率显著高于981(p<0.05),在8月12日之后,两种品系之间不存在显著性差异(p>0.05)(见图3)。
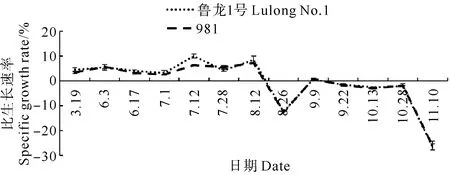
图3 胶州湾栽培季节龙须菜比生长速率变化
2.2 胶州湾栽培海区环境因子动态变化
2.2.1 温度和光照 在胶州湾海区中,海水的温度随栽培时间发生规律性变化,呈先上升后下降的趋势,具有明显的峰值(见图4)。栽培初期,海水温度在15 ℃左右,随后逐步上升。8月份,海水温度达到25 ℃以上,全天水温>26 ℃的天数为19 d,最高水温>28 ℃的天数为21 d,高温持续了近3周的时间,随后开始初步降低。至实验结束时,温度已经下降到15 ℃以下。
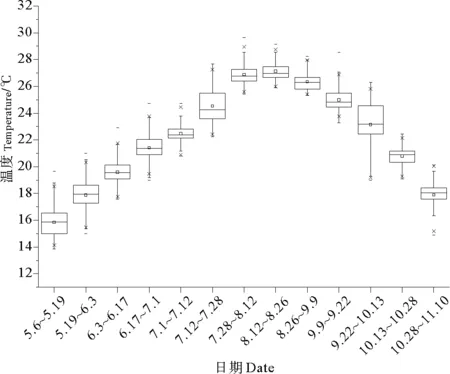
图4 胶州湾栽培季节海水温度变化
胶州湾海区龙须菜栽培层中,日光强度的日变化程度剧烈(见图5)。记录的瞬时最强的光照强度在7月1日上午11:30,为1 388.8 μmol·m-2·s-1;日平均光照强度最高的是7月1日~7月12日,数值为321.6 μmol·m-2·s-1;7月12日~7月28日和10月28日~11月10日的日平均光强较低,分别为22.25和20.07 μmol·m-2·s-1。在整个栽培光照监测中,出现高光照(光强>200 μmol·m-2·s-1)的次数为243次,占总检测数的5.23%。

图5 胶州湾栽培季节海面日平均光强变化
2.2.2 营养盐与海水溶解无机碳 在胶州湾内,对龙须菜可大量利用的碳素和氮素、磷素营养盐进行检测,主要包括:氨态氮(NH4-N)、硝态氮(NO3-N)、磷酸磷(PO4-P)及DIC(海水溶解无机碳),结果如图6所示,在不同的栽培阶段各环境因素表现各不相同。
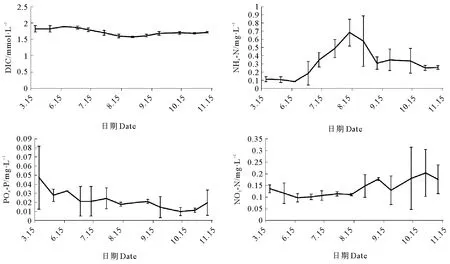
图6 胶州湾栽培季节海水营养盐与溶解碳变化情况
海水中溶解碳(DIC)初期的浓度值为(1.847±0.077) mmol·L-1,经过3个月的时间,胶州湾海水溶解碳的浓度下降至(1.600±0.042) mmol·L-1,与实验初期有显著性差异(p<0.05)。8月份后海水溶解碳浓度缓慢回升,恢复至实验初期的水平,无显著性差异(p>0.05),总体呈先下降后上升的趋势。
NH4-N在0.082~0.799 mg·L-1范围内,呈先上升后下降,最后趋于平稳的趋势。5月至6月期间一直维持在较低的水平,最低点出现在6月份,浓度值为(0.082 4±0.002) mg·L-1;高峰出现在8月份,浓度值为(0.596±0.220) mg·L-1,与初期的低浓度有显著性差异(p<0.05)。随后逐步下降,9月份后较为平稳,未发生显著性变化(p>0.05)。NO3-N浓度没有显著的变化(p>0.05),平均值为(0.133±0.034) mg·L-1;最高浓度出现在10月28日,数值为(0.204±0.100) mg·L-1;最低浓度出现在6月17日,数值为(0.097±0.017) mg·L-1。
在栽培期间,PO4-P浓度整体呈下降趋势。初始浓度为(0.027±0.006 9) mg·L-1;10月12日到达最低峰,为(0.009±0.004) mg·L-1,两者具有显著性差异(p<0.05)。
2.2.3 盐度、pH、光周期 在青岛胶州湾栽培海区,对海水理化性质及影响龙须菜生长的环境因子(光周期)作进一步检测,结果表明(见图7):在不同时期,海水盐度、pH和浊度保持在较为平稳的水平。海水盐度表现的最为平稳,几乎无波动,均值为31.4±1.5;pH呈先上升后下降最后趋于平稳的趋势,高峰出现在6月1日与6月12日,为8.17±0.85,与8月12日之后的各组pH之间具有显著性差异(p<0.05)。浊度围绕1.8 m上下波动较为频繁,均值为(1.80±0.37) m。光周期呈现规律性变化,随栽培时间的推移逐渐缩短,初期变化幅度较小,均值为(13.8±0.6) h。8月份后逐步缩短,8月26日之后各组与实验初期的光周期之间具有了显著性差异(p<0.05)。

图7 胶州湾栽培季节海水盐度、pH、浊度及光周期变化
2.3 龙须菜生长与环境因子关系
Pearson线性相关系数分析表明,在10个监测的环境因子下,硝态氮和光周期与胶州湾栽培龙须菜比生长速率之间具有显著相关性(见表1)。其中,硝态氮与胶州湾栽培的两种龙须菜比生长速率之间的相关性分别为-0.56和-0.549(p<0.05),光周期与两种龙须菜比生长速率之间的相关性分别为0.743和0.729(p<0.01)。

表1 胶州湾栽培龙须菜生长速率与各环境因素的相关系数
在相邻两组之间(见表2),两种龙须菜的生长速率在相邻两组数据之间的变化较大,特别是在龙须菜生长较为迅速的6~7月,与生长显著下降的8月份组间表现为极显著的差异。而环境因素的变化相对较为平缓,除了温度、光周期的个别组外,相邻两组之间的差异并不明显。环境的平稳变化造成了栽培龙须菜比生长速率较大幅度的改变,从一定程度上说明龙须菜生长对环境因素的变化较为敏感。

表2 胶州湾栽培龙须菜生长速率及环境因素组间差异性统计
2.4 栽培龙须菜显微形态观察
实验初期,龙须菜生长状况较好,表明此时环境条件适合龙须菜的生长,龙须菜藻体表皮细胞排列紧密(见图8a和f,箭头)。
8月后,龙须菜生长速率有了显著的下降,此时日平均温度在25 ℃左右,龙须菜面临高温胁迫。在此阶段,藻体发育产生四分孢子囊(见图8b、g,箭头),周围表皮细胞排列开始疏松(见图8c、h,白色箭头),孢子囊发育成熟释放四分包子后,留下大量“孔洞”(见图8c、h,黑色箭头),表皮形态不完整,与苗绳链接处藻体脱落严重。
进入9月后,水温逐渐下降,龙须菜生长速率显著回升,藻体开始恢复生长。藻体表面“孔洞”有愈合的趋向(见图8d、i,箭头),但仍有疏松的部位(见图8a、f和d、i)。
龙须菜在经历了短暂的恢复后,生长又开始下降。一方面,海水中营养盐溶解碳、磷处于较低的水平;另一方面,光周期显著下降,白天的时长已经小于12 h。表皮层与愈合趋势的“孔洞”,沿边缘处扩张,显微镜下“孔洞”透光性增强(见图8e、j,黑色箭头),大量微藻、虫卵等在溃烂处附生(见图8e、j,白色箭头),龙须菜断裂脱落现象严重。此时龙须菜状态极差,与正常状态下的龙须菜表皮形态有明显的差别(见图9)。
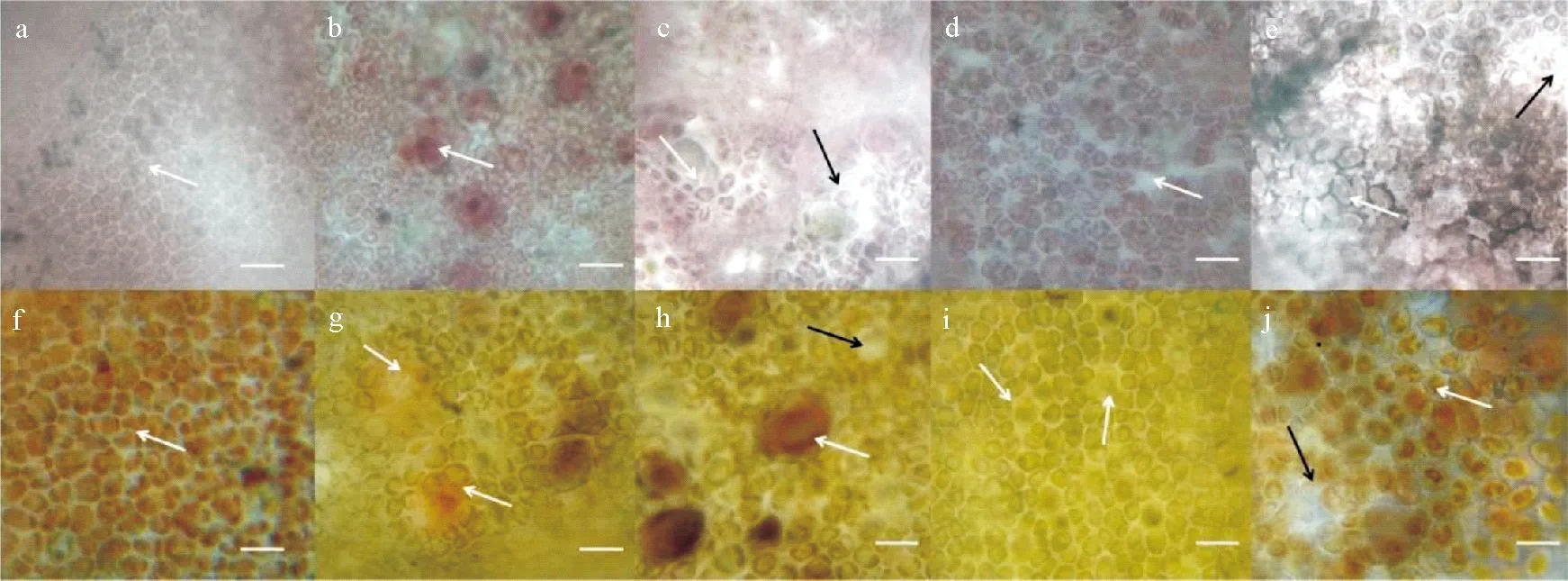
(a和f:6月17日;b和g:8月12日;c和h:8月26日;d和i:9月9日;e和j:10月28日。a~e:品种鲁龙1号;f~j:品种981。标尺=20 μm。 a and f: June 17; b and g: August 12; c and h: August 26; d and i: September 9; e and j: October 28. a~e: cultivar “Lulong no. 1”; f~j: cultivar “981”. Scale bar =20 μm.)
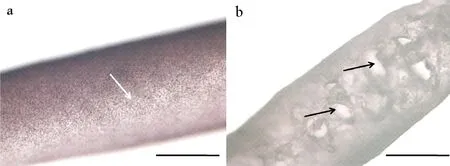
(a:6月17日;b:10月28日;标尺=1 mm。a: June 17; b: October 28; Scale bar=1 mm.)
2.5 胶州湾栽培龙须菜生长模型初建
由于得到的龙须菜鲁龙1号与981两个品系的比生长速率变化趋势一致(见图3),除7月份外其他时间均无显著性差异(p>0.05),故以鲁龙1号的数据为代表进行模型构建。将所有环境因素作为自变量,龙须菜生长速率为因变量构建回归模型(见表3)。由于对该模型整体的F检验没有通过显著性测试(Sig.>0.05),且模型系数的T检验Sig.也均大于0.05,因此初始模型整体不具有统计学意义。
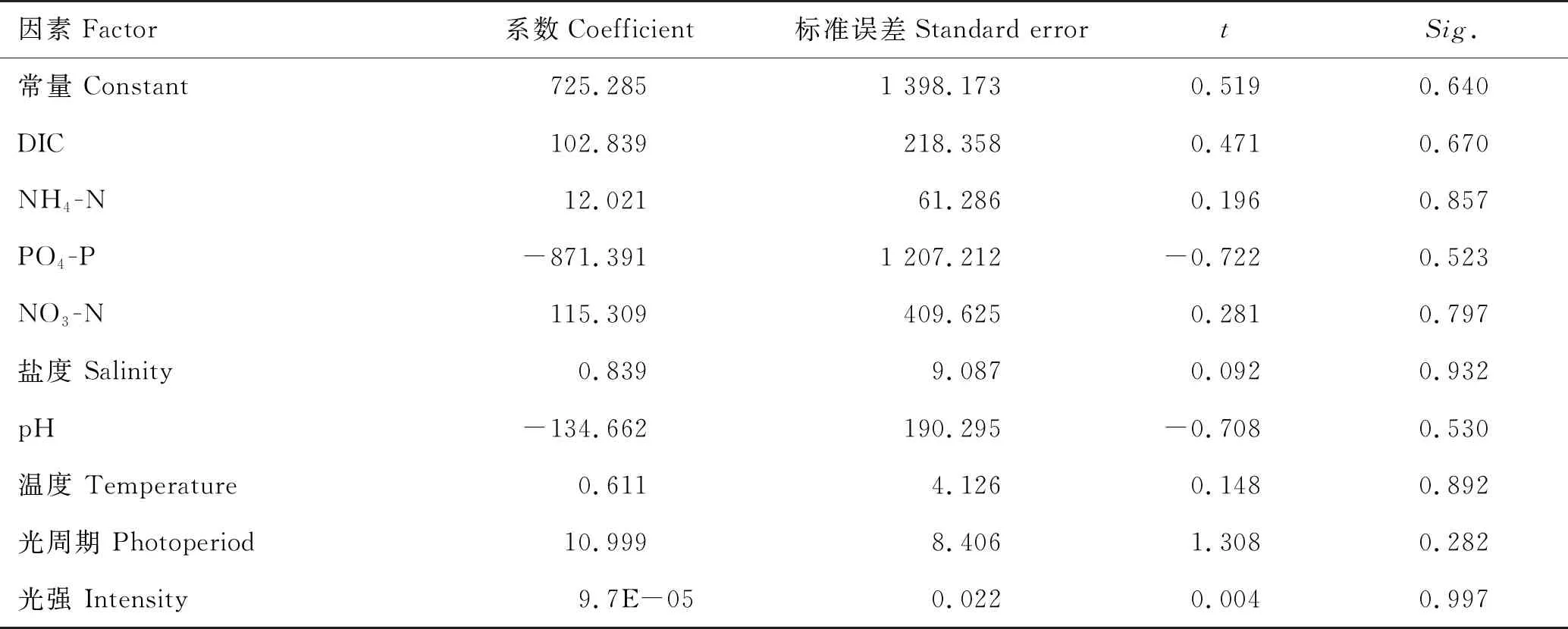
表3 以所有环境因素构建的龙须菜生长初始模型*
以全部因素建立的模型不理想,需要对参数进行筛选。通过对初始模型的权重分析来挑选影响最大的因素再次进行模型的构建。采取Shapley值分析方法定量的确定各项参数对初始模型R2的贡献来提供参考(见表4)。

表4 不同因素对初始模型的重要性贡献
在所有因素中,与龙须菜比生长速率最相关的光周期对初始模型的贡献最高,为第二位PO4-P的2倍,其次为pH、DIC。这四种因素对初始模型的标准化权重系数之和为0.6,即它们的贡献之和占全部的60%。参与初始方程的构建因素有9个,因此平均的标准化权重系数为1/9,而其他的因素的贡献均远低于此平均值。通过权重分析,说明光周期、PO4-P、pH、DIC这四种因素相较于其他因素对比生长速率有着更强的解释能力。
以这4种因素分别构建了单一因素、双因素、三因素及四因素的四个模型来具体比较(见表5)。构建的四个模型经回归模型方差分析检验(Anova)均具有统计学意义(Sig.<0.05)。从模型的系数来看,模型1、模型2和模型3的系数T检验均未通过(Sig.>0.05)。模型4中的常量、光周期的系数T检验Sig.均小于0.05,模型最为理想。
以模型4中的光周期因素为基础,再将其他所有环境因素重新进入模型中进行比较,表现情况如表6所示。除盐度因素外,其他双因素模型的调整R2(见表6)均低于原模型4的调整R2(见表5),且插入变量系数的t检验均未通过(Sig.>0.05),因此,选择模型4作为最终建立的回归模型。
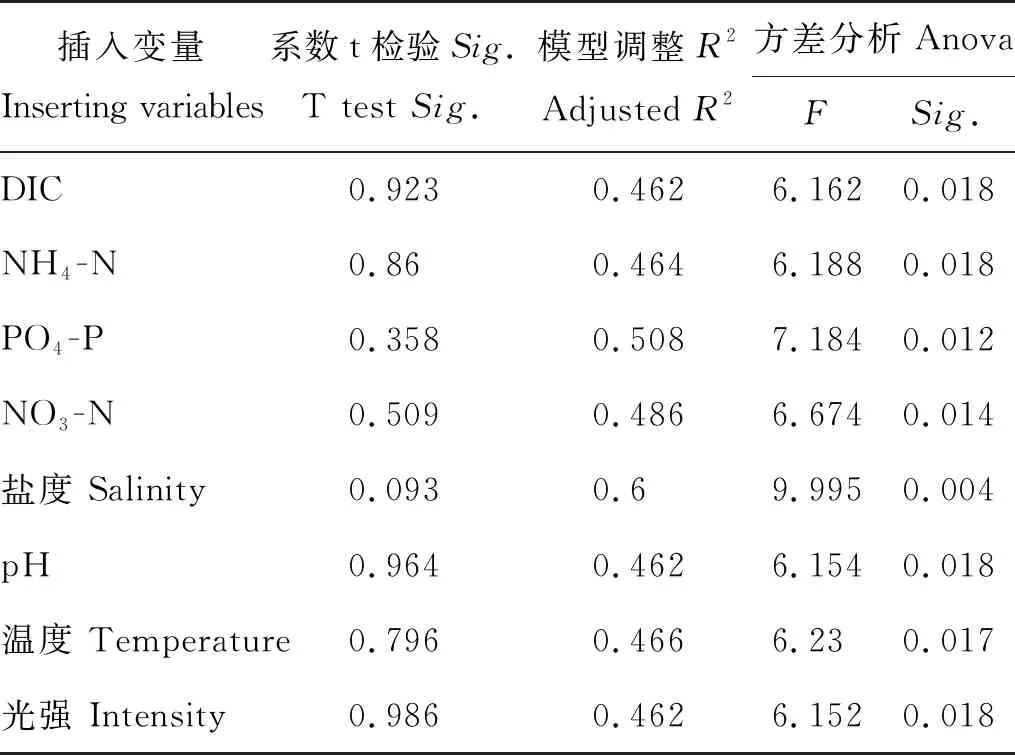
表6 以模型4为基础插入变量的表现
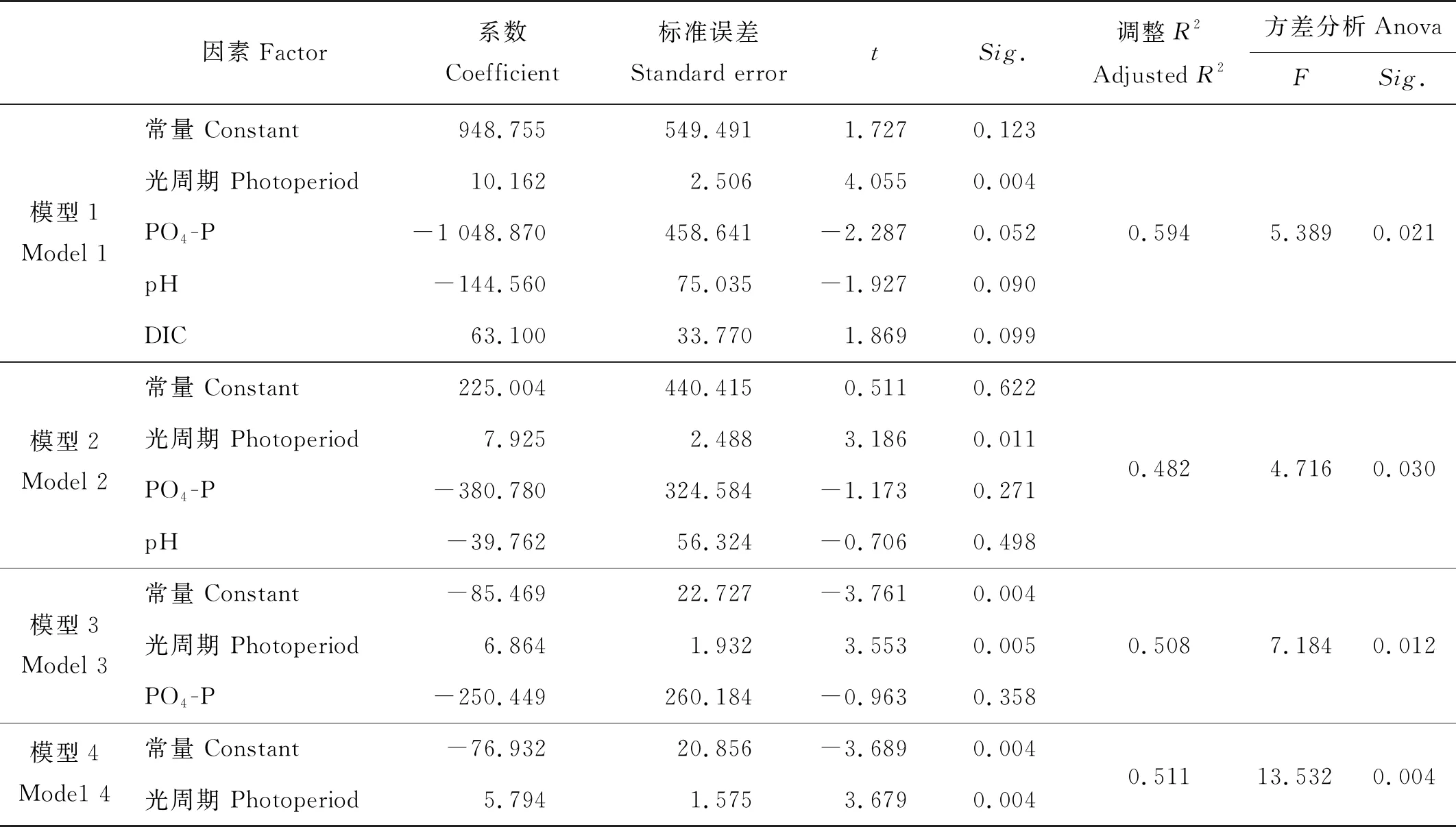
表5 以龙须菜比生长速率为因变量构建的4个模型
综合以上分析,确定最适合用来预测龙须菜比生长速率的回归方程:YSGR=5.794X光周期-76.932(R2=0.511)。将观测到的环境数据带入此方程后,实际观测值与预测结果如图10所示。两组数据整体无显著性差异(p>0.05)。

图10 胶州湾栽培龙须菜生长速率实测值与预测值
3 讨论
3.1 栽培龙须菜的生长速率与环境因素的关系
在海区环境中,光照、温度随季节发生规律性变化。其中,天气、海水的透明度等条件会影响龙须菜在生长过程中受到的实际光照,这使光照条件的变化较为频繁。当藻类暴露在高光强下,大型海藻表现出明显的光抑制或者不可逆的光损伤[14],反之当光照条件不足时,藻类又不能充分地进行光合作用影响生长。实验室条件下,100 μmol·m-2·s-1的光照强度比较适合龙须菜的生长[15]。本次实验中,龙须菜生长速率与光照强度之间呈一定的正相关关系,特别是7月1~12日受到的平均光照强度较强且龙须菜生长较快,9月22日~10月13日受到的平均光照强度较弱且龙须菜生长较慢(见图5),说明自然条件下胶州湾栽培龙须菜未受到光抑制,光照的提升有助于龙须菜的生长。温度对植物的影响表现在低温抑制、高温损伤,相对于植物其它代谢活动,高温对光合作用的损伤最为敏感,尤其是PSII,引起膜脂的过氧化链式反应,促使细胞膜的功能受损,导致光合器官严重破坏[16]。在实验室条件下龙须菜的最适生长温度为15~25 ℃[10]。本实验所得的温度数据变化规律明显,且数据覆盖了15~25 ℃整个区间(见图4)。实验初期,龙须菜生长较为稳定,此时温度由15 ℃逐步上升至25 ℃左右,此时龙须菜的生长并没有随温度的变化而产生明显的规律性变化,说明在15~25 ℃这一温度区间内,龙须菜均能正常生长。在7月28日之后,气温一直维持在25 ℃以上,最初两周内龙须菜生长维持正常,经历了两周高温与高光诱导后,龙须菜开始产生四分孢子(见图8b和g),此时龙须菜因为表皮受到损伤,生长状态较差,同时藻体断裂脱苗严重,导致绳上的生物量大幅下降。较长时间的高温(>27 ℃)与高光的诱导不利于龙须菜的生长,这与Ren等实地栽培的结果一致[17]。
营养盐的浓度与大型藻类的生长之间关系复杂,一方面藻类的生长需要营养盐,较高浓度的营养盐有利于藻类的生理活动;另一方面,藻类的生长也会吸收营养盐,使其浓度降低[18-20]。本次实验中DIC与氨态氮变化较为明显(见图6)。有研究表明龙须菜在高磷水平下,CO2的加富提高了龙须菜的光合作用能力。在实验期间,海水中的溶解碳浓度逐渐下降,在8月26日到底达低点,此时龙须菜生长速率显著降低(p<0.01)。在DIC浓度逐步升高后,龙须菜生长也有一定程度的恢复。有研究表明栽培龙须菜能有效地吸收海水中的氨态氮,使其浓度降低[8];在本次实验中,氨态氮的浓度与龙须菜的生长速率呈显著负相关(p<0.05),这可能与龙须菜对氨态氮的吸收有关。在龙须菜栽培的海区,龙须菜的快速生长有效的利用了海水中的氨氮,使海区的氨氮浓度维持在较低的水平。
海水的酸碱度与盐度较为稳定(见图7),对养殖龙须菜的生长影响有限。海水浊度对龙须菜生长的影响,体现在影响龙须菜受到的光照方面。相关性分析表明,龙须菜生长与光周期呈极显著相关(p<0.01),光周期的影响主要体现在和光照、温度等的交互作用上,Zhou等[22]研究表明在光周期为LD 8:16~14:10 h 范围内,龙须菜最高的生长速率出现在LD 12:12 h,继续缩短光周期不利于藻体生长。本研究中,龙须菜栽培至8月12日之后,光照期缩短至 (11.7±0.6) h,不利于藻体生物量积累,推测原因为光照期的缩短意味着光合作用总时长减少,藻体内形成 ATP 时间减少,消耗ATP 时长增加(黑暗期)。同时,龙须菜受高温高光影响放散四分孢子后藻体表面留有大量的孔洞(见图8),Wang 等[23]研究指出龙须菜孔洞形成后,需要通过光合作用产生大量能量(ATP),用于愈合藻体表面孢子放散后留下的“伤口”,缺少ATP也不利于孔洞的愈合。
综合以上,可见在实验初期环境因素适合龙须菜的生长。在温度提高到25 ℃两周时间后,诱导龙须菜放散四分孢子,藻体表面受到损伤;逐步缩短的光周期与较低的海水溶解碳浓度使龙须菜的光合作用减弱,产出的能量(ATP)一方面要修复藻体损伤,另一方面又要维持正常的生理活动,ATP严重供给不足。损伤的进一步扩大使得情况更加恶劣,由此进入恶性循环。因此,实验后期生物量的下降的主要原因是由于龙须菜受高温诱导进入繁殖期后,四分孢子的放散使藻体受损出现脱苗现象。
3.2 胶州湾龙须菜生长模型
模型的模拟可在实验室及室外实地环境下开展,在实验室内条件过于稳定,所构模型未必适合海区环境。在海区环境中,光照强度变化最为剧烈,日变化可以从0~1 388.8 μmol·m-2·s-1间波动,不同时间及同一时间不同地点均有明显地变化;温度变化次之,在不同的栽培阶段,也发生明显变化;营养盐等其它海水理化指标在特定时间内维持较为稳定的水平,复杂多变的环境和各环境因素间的交互作用,为模型的构建增加了困难。通过逐步回归的方法构建的生长模型中,胶州湾龙须菜的生长与光周期(光照期)密切相关,以光周期的值来反映胶州湾栽培龙须菜的生长有较好的效果。其他因素虽然在逐步回归中被多重共线性变量的筛选而被剔除,但并不说明对龙须菜生长无影响。精确模型的模拟需要大数据的支持[30],为了完善胶州湾龙须菜生长模型,需要掌握多组龙须菜生长状况与环境因素累年变化的数据。
本次实验获得的胶州湾环境数据的时间跨度大,平行性较好,且无异常数据,实验海域选在常规栽培海域,对于胶州湾海区具有一定的代表性。且胶州湾海区作为黄海沿岸重要的养殖区,实验反映了夏季北方龙须菜养殖环境因素的具体变化情况。但是由于龙须菜在中国南方北方均有栽培,各地区的环境因素存在一定的差异,所以单一地区的数据构建的模型具有较大的局限性。本模型针对胶州湾海域,能够较好预测鲁龙1号、981这两个优良龙须菜品种的生长状况,也为不同海域龙须菜栽培动力学模型构建提供了借鉴。
