野生达乌尔黄鼠与室内F1代的社会性及社会行为比较
2020-08-03杨晨希马小梅王建礼
王 恒 赫 晨 杨晨希,2 马小梅 王建礼,2*
(1.北方民族大学生物科学与工程学院,银川,750021;2.国家民委生态建模与应用重点实验室,银川,750021)
群居动物的社会性反映了某个个体与其他个体交流的倾向特性,有利于促进生殖与发育、认知思维能力以及协同合作能力,提高广义适合度[1-2]。动物的社会行为主要是种群内的个体相遇及相遇后的一系列相互作用。同种个体间的社会互作行为可反映其在种群内的社会地位[3-4],有助于种群内信息交流和共享,有利于逃离捕食者,提高动物觅食的效率,保证幼体得到双亲抚育,并使动物更易接近异性以完成交配,以保证种族的延续和发展[5]。关于啮齿动物个体间的社会行为已有大量研究,如对布氏田鼠(Lasiopodomysbrandtii)[6-7]、小家鼠(Musmusculus)[8]、草原田鼠(Microtusochrogaster)和棕色田鼠(Lasiopodomysmandnarinus)[9-10]等的研究。
动物行为的变化也是生物多样性的一个组成部分[11]。与社会性相关的社会行为是高度社会化的反映,是动物长期适应环境的结果。在以往的研究中发现环境变化会对个体的生理活动、情绪、认知以及行为产生深远的影响。例如,多配制的草甸田鼠(Microtuspennsylvanicus)父本行为极少,但当温度下降,白天变短,雄鼠会进入幼仔巢区并表现出父本行为[12-13];大鼠在丰富饲养环境下表现出更强的社会行为[14-15]。栖息地对社会行为也有重要影响,非洲野驴(Equusafricanus)在干旱和湿地环境中的交配体制和社会组织会呈现变化[16-17]。美国中部伊利诺伊州的草原田鼠种群和东部堪萨斯州的草原田鼠种群相比,具有不同的社会性和社会行为,伊利诺伊州种群具有符合单配制特点的高度社会性和社会行为,如配对关系的形成和双亲抚育[18],而堪萨斯州的种群缺乏社会性,呈现出多配制的特征,包括低水平的身体接触[19],更多的攻击性[20],更少的异亲和双亲行为[21]。研究棕色田鼠也发现,可能受到降水、食物资源等影响,捕自河南灵宝市程村的种群与新郑种群不仅在体重有着显著差异,也表现出焦虑样行为、配偶选择等行为上的差异[22-23]。占据高原草原化草甸的高原鼠兔(Ochotonacurzoniae)和占据裸岩栖息地的北美鼠兔(Ochotonaprinceps),尽管二者都属于鼠兔属,但面对选择压力的行为反应相反[24]。这些发现说明环境对动物行为的复杂作用。
达乌尔黄鼠(Spermophilusdauricus)为地栖型松鼠科(Sciuridae)动物,具冬眠习性,是研究冬眠生理的重要模式动物。已有学者在其冬眠调控机制、室内繁殖和骨骼肌适应等方面做了相关研究[25-27],但关于达乌尔黄鼠行为学方面的研究较少,已有的研究仅涉及活动节律和范围等方面[28-29]。本实验采用社会互作、社会性偏爱及旷场实验,研究了野生达乌尔黄鼠的两性社会行为和室内F1代的社会行为,拟对不同出生及成长环境下的达乌尔黄鼠社会行为进行比较。
1 材料与方法
1.1 动物
达乌尔黄鼠捕于宁夏盐池县惠安堡,塑料饲养笼(46 cm×31.5 cm×20 cm)饲养,同性别鼠3—4只一笼。室温25 ℃,光暗周期为12L∶12D,喂以鼠饲料(沈阳茂华生物科技有限公司)和胡萝卜,饮水充足。捕获黄鼠中的孕鼠生产的雌性幼仔作为室内出生F1代鼠,孕鼠于4月底至5月初分娩,幼鼠至40日龄左右与母鼠分开,同性鼠3—4只一笼,待60日龄后于7月进行实验。捕获的雌、雄野生鼠同样在7月实验。
1.2 旷场行为
旷场实验可用来评估实验动物的焦虑水平和运动能力[30]。旷场箱的结构参照Fiore和Ratti[30],旷场箱(长50 cm×宽 50 cm×高25 cm)底面分16等分,中间4等分为中央区域,剩余 12等分为周围区域。在旷场箱正上方 1.5 m 处安放4×60 W红色灯泡照明。以中央区域停留时间评估焦虑样行为,穿格总次数评估运动性。行为观察开始时,实验鼠(雄鼠,n=11;雌鼠,n=16;F1代雌鼠,n=8)被置于旷场箱中央区域,使用DV 摄像机(HDR-XR260E,Sony)拍摄5 min。每次实验结束后,使用75%的酒精对旷场箱进行清洗以清除气味。
1.3 社会性偏爱
社会性偏爱实验是评价社会能力的主要方法之一[31]。采用三箱式社交实验箱(长120 cm×宽80 cm×高45 cm,XR-XJ117,上海欣软信息科技有限公司)测试,箱体由聚乙烯材料构成,中间有两段隔板,将箱体分成3室。中央室下方有允许动物自由移动的开口。实验开始前将实验鼠(雄鼠,n=8;雌鼠n=7;F1代雌鼠,n=7)放于中央室,两侧室分别放一个等大束缚笼,一个笼内放置同性陌生鼠,另一个为空笼。实验鼠在中央室适应5 min后,打开通向两侧的小门,用SuperMaze软件(上海欣软信息科技有限公司)跟踪记录10 min内实验鼠对陌生鼠与空笼的探究频次及时间。每次试验后用75%酒精擦拭箱体及束缚笼,去除气味影响。
1.4 社会互作
选择质量与实验鼠相似的同性个体作为刺激鼠,刺激鼠以背部剪毛作为标记。观察箱底部覆以2 cm厚的木屑。测试时将实验鼠(雄鼠,n=11;雌鼠,n=14;F1代雌鼠,n=8)和同性刺激鼠分别置于观察箱两侧,中央用木板隔开,适应5 min后,使用SuperMaze软件记录15 min内实验鼠以下行为的持续时间,行为类型的划分和定义参见先前的文献[32-33]
(1)社会探究(social investigation):嗅闻同伴的身体的任何一个部分(包括肛殖区、头面部及躯体等)。
(2)跟随(following behavior):刺激鼠走动,实验鼠紧随其后。
(3)亲密行为(affiliative behavior):包括聚团在一起以及舔同伴的皮毛或爬在同伴的上面或下面,大部分情况下修饰的动物会把自己的前爪放在同伴的颈部或背部。
(4)攻击行为(aggressive behavior):扑击、撕咬、翻滚和追击。
(5)自饰(self-grooming behavior):用嘴舔舐或用爪有节律的在口部、面部、耳部、腹下、侧肋、肛殖区等处挠动。
(6)非社会行为(nonsocial behavior):静卧、挖掘和爬笼等行为。
1.5 统计分析
使用SPSS 19.0进行统计学分析,组间比较采用独立样本t检验分析。数据以平均数±标准误表示。显著性水平设为P<0.05。
2 结果
2.1 旷场实验
雄性和雌性黄鼠相比,在旷场中央区域的停留时间(t=0.312,P=0.758)与穿格次数(t=1.464,P=0.156)没有显著性差异(图1a-b)。与野生雌性黄鼠相比,室内F1代雌性在旷场中央区的停留时间显著增加(t=2.352,P=0.028),但穿格次数(t=2.024,P=0.055)没有显著差异(图2 a-b)。
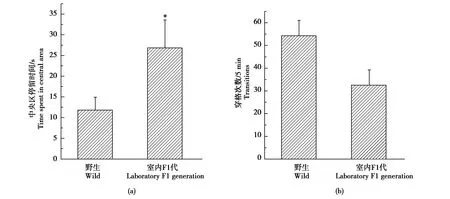
图2 野生雌性和室内F1代雌性达乌尔黄鼠的旷场行为Fig.2 Behaviors of wild-deprived and laboratory F1 generation daurian ground squirrels in an open field
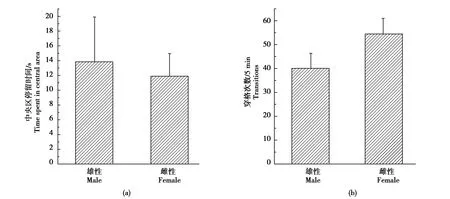
图1 野生雌、雄达乌尔黄鼠的旷场行为Fig.1 Behaviors of wild-deprived male and female daurian ground squirrels in an open field
2.2 社会性实验
野生雄性黄鼠对陌生鼠所处笼子的探究时间(t=0.557,P=0.586)和探究频次(t=0.286,P=0.779)与空笼相比没有显著差异(图3a-b)。野生雌性黄鼠对陌生鼠所处笼子的探究时间(t=1.590,P=0.138)和探究频次(t=2.042,P=0.064)与空笼相比没有显著差异(图3c-d)。室内F1代雌鼠对陌生鼠所处笼子的探究时间(t=3.562,P=0.012)和探究频数(t=4.002,P=0.007)显著高于对空笼的探究(图3e-f)。
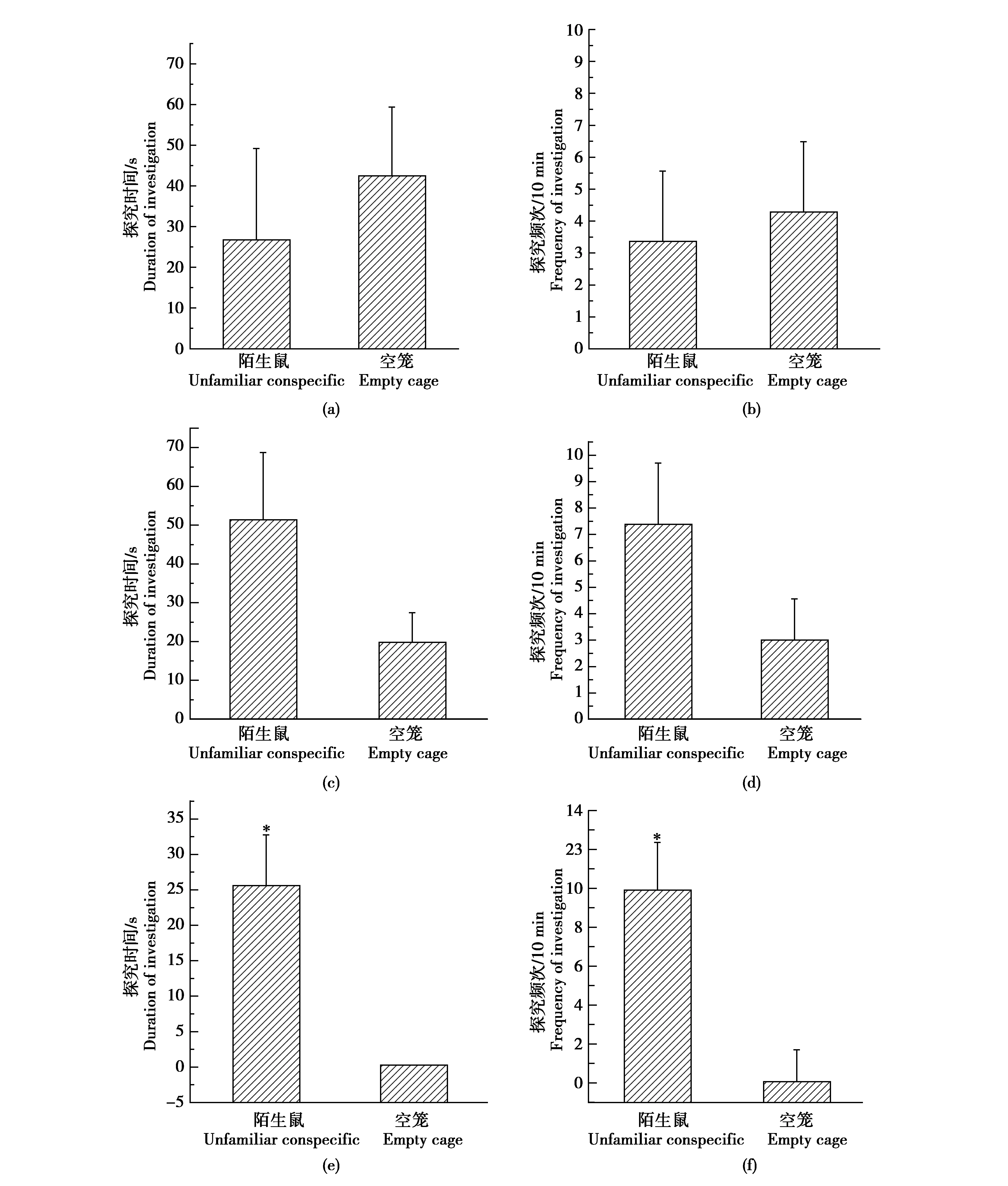
图3 达乌尔黄鼠的社会性偏爱Fig.3 Social preference in daurian ground squirrels
2.3 社会互作实验
野生雌性达乌尔黄鼠与雄性相比,探究行为(t=0.608,P=0.549)、亲密行为(t=0.464,P=0.647)、自饰行为(t=0.875,P=0.391)、跟随行为(t=0.444,P=0.662)和非社会行为(t=0.458P=0.651)的差异均无统计学意义(图4)。与野生雌性黄鼠相比,室内F1代雌性的亲密行为(t=6.042.,P<0.001)和自饰行为(t=2.484P=0.027)显著降低,非社会行为增加(t=4.243,P<0.001);探究行为(t=0.386,P=0.704)和跟随行为(t=0.758,P=0.457)没有显著差异(图5)。
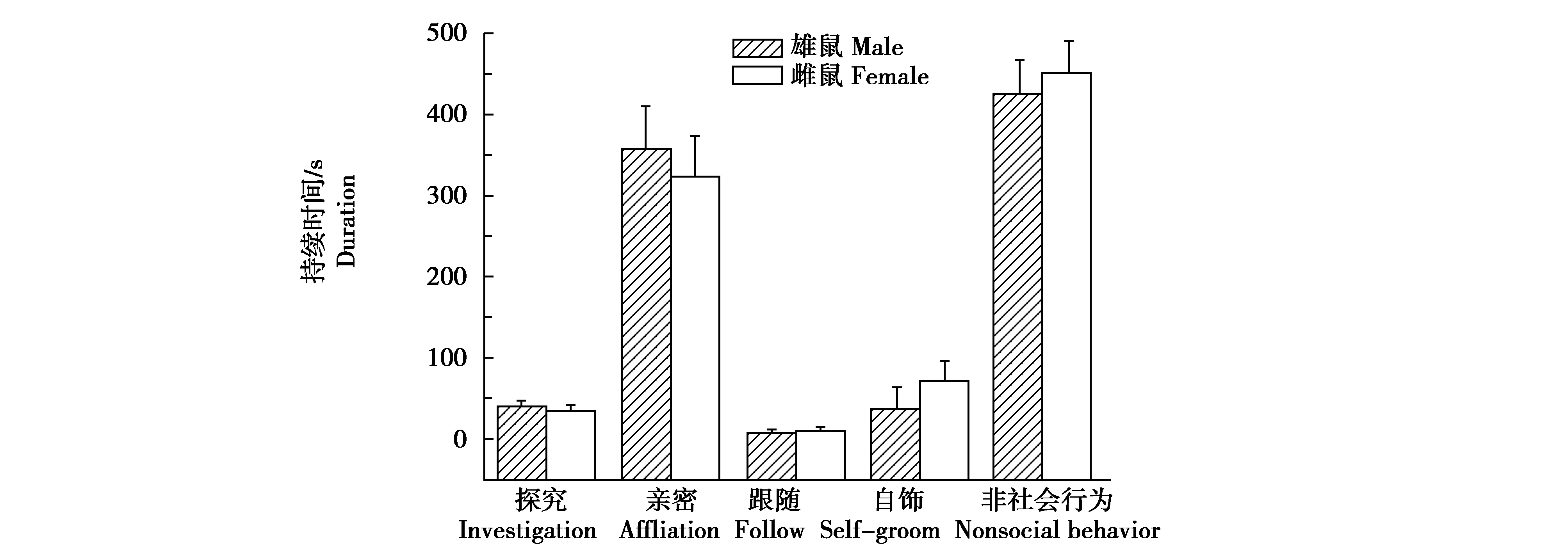
图4 野生雌、雄达乌尔黄鼠的社会互作Fig.4 The duration of social interaction in wild-deprived male and female daurian ground squirrels
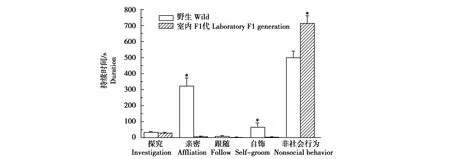
图5 野生和室内F1代雌性达乌尔黄鼠的社会互作比较 *P<0.05Fig.5 The duration of social interaction in wild-deprived and laboratory F1 generation daurian ground squirrel *P<0.05
3 讨论
3.1 达乌尔黄鼠的运动性与焦虑性
焦虑是社会行为的重要组成,旷场实验可用来评估实验动物的焦虑水平和运动能力及在新环境下的自主行为和探究行为[30]。运动能力在新环境中是一个复杂的行为,和很多因素相关,包括动物的焦虑水平、对应激的反应和对新事物的反应[34]。本研究发现,野生雌、雄达乌尔黄鼠的焦虑水平和运动性没有显著差异,这与对其他一些动物的研究结果相似,例如,在研究ICR小鼠中没有发现运动性和焦虑水平的性别差异[35-36];对C57BL/6J 和BALB/cJ小鼠研究也发现性别因素对旷场试验的穿格次数和中央区时间的影响较小[37]。但对棕色田鼠研究发现,无论河南程村种群还是新郑种群,雄性均表现出了较高的焦虑水平,且新郑种群的运动性表现出了性别差异,程村种群却没有[22]。
本实验发现,与野生雌性达乌尔黄鼠相比,室内F1代雌性在中央区域停留时间显著高于野生雌性,暗示焦虑水平降低,但二者的运动性没有表现出明显差异,再次说明运动性和焦虑水平没有必然的相关性。研究发现焦虑水平同样受栖息环境影响,河南灵宝市程村的气候干燥、海拔较高,而新郑则气候湿润、海拔较低,两地在降水和食物资源上有差异,程村的棕色田鼠种群比新郑种群表现出更高的焦虑样行为[22]。在野外达乌尔黄鼠的活动范围一般在100 m左右[38]。本实验中F1代黄鼠自出生至成体,一直生活在实验室饲养笼内,活动空间狭小,生活环境缺少变化且安定,没有食物和天敌之虞,因此表现出较低的焦虑水平,而野生鼠出生于野外,在成年时被捕回,虽然在实验室适应了一个多月,但仍表现出较高的焦虑水平。与此结果类似的是,研究发现野生小家鼠与实验室驯化后的小鼠如BALB/c和C57BL/6相比,在旷场中的活动性降低,表现出更强的逃避空旷地方的行为,在陌生环境下,野生小鼠进入潜在危险区变得更加谨慎[39]。
3.2 达乌尔黄鼠的社会性与社会互作
许多啮齿动物包括小鼠具有较强的社群性,实验室内可量化其社会行为,啮齿类的社会偏爱测试已被广泛用于研究小鼠、大鼠和田鼠的社会性倾向[40]。在社会偏爱性实验中,雌、雄野生达乌尔黄鼠对陌生鼠所在笼与空笼的探究时间及频次均未达到显著差异,相比之下,具有高水平社会性的小鼠如C57BL/6J小鼠会花费更多的时间在陌生鼠箱内[40],说明雌、雄达乌尔黄鼠的社会性不高。野外研究发现达乌尔黄鼠除在交尾期偶有雌雄同居外,皆为独居。雌性达乌尔黄鼠分娩后28 d幼鼠开始独立取食,34—36 d,分散打洞,开始分居,至7月则大量分居独栖[38],野外的这种栖居习性也印证了其较低的社会性。在社会互作实验中,雌、雄达乌尔黄鼠的社会探究、亲密行为、跟随及自饰行为均没有表现出性别差异,与社会偏爱性的表现一致。本实验中未发现达乌尔黄鼠有明显的攻击行为,因此未做统计。在ICR小鼠也没有发现社会探究及身体接触的性别差异[35]。对棕色田鼠的同性社会互作研究发现,雄性棕色田鼠比雌性有较多的亲密行为、较少的攻击行为,但社会探究行为没有显著差异[41]。草原田鼠也表现出两性差异,尤其是攻击行为[19]。对C57BL/6和BALB/c小鼠研究发现,雌性较雄性表现出更高的社会探究[37]。这些差异既反映了种的特异性也反映了其社会性程度。社会探究是成年动物典型的社会行为[42],亲密行为中的聚团行为可能更多的是一种保护性行为,自饰行为被认为是一种情绪的暗示[43]。不同形式的互作行为具有不同的生态和发育模式,并被大脑不同的神经化学环路调制[42]。本研究中雄性达乌尔黄鼠的非社会行为在总测试时间中占比达到46.9%,雌性占比为49.9%,二者平均为48.4%。对多配制的雄性东方田鼠(Microtusfortis)室内测试发现,其非社会行为的时间占比也高达40%以上[44]。相比之下,单配制动物具有更复杂的社会互作[45],对单配制的雌、雄棕色田鼠研究发现,其非社会行为的时间占比仅约20%左右[44,46],间接说明达乌尔黄鼠的社会性远较棕色田鼠为低,是一种低社会性动物。
在社会性偏爱实验中,室内F1代鼠对陌生鼠所处笼子的探究时间及频次均高于空笼,说明室内F1代鼠有较积极的社会性,这可能与其出生后与双亲或同胞鼠所处时间较长有关。但F1代鼠与野生鼠相比,在社会互作中其亲密行为及自饰行为均降低。在棕色田鼠研究中也发现了F1代鼠与野生鼠的行为差异,程村的雄性棕色田鼠种群较新郑的雄性种群具有更多的攻击和社会性,但这种差异在F1代却没有发现;野生新郑田鼠对陌生鼠有显著偏好,但F1代却呈现出对熟悉鼠的偏好,且表现出对陌生鼠友善行为和更少的攻击行为[23]。这些实验结果表明,室内出生的动物其社会行为特征会发生变化,室内的环境条件可能影响其行为发育模式。
精氨酸加压素(AVP)和催产素(OT)是调节动物社会行为非常保守且重要的神经肽。加压素受体(AVPR1a)和催产素受体(OTR)密度和分布的变化常与社会行为有关,例如,在田鼠属动物侧隔的AVPR1a和OTR以及杏仁核的AVPR1a对社会行为很重要[47]。与达乌尔黄鼠亲缘关系相近的瑞氏黄鼠(Spermophilusrichardsonii)在野外进行社群生活,在繁殖期具有报警鸣叫行为,社会性对于瑞氏黄鼠的生存很重要,属于中度社会性。研究其脑区发现AVPR1a和OTR的分布与调节社会行为和通讯有关,青春期时的雄性和雌性AVPR1a和OTR的分布和密度没有显著性差异[48],但在成体的一些脑区发现有差异[49]。此外,有研究表明调节社会行为的神经环路在社会性和非社会性物种不同[50],胼胝体大小相对于脑的重量也与动物的社会性有关[51]。因此,开展AVP和OT系统以及其胼胝体相对大小的研究可能有助于本研究中对达乌尔黄鼠社会行为表现的解释。
3.3 结语
动物行为是长期进化的产物,是其生活环境、遗传发育和成长经历等多方面相互作用的结果,其中任何一方面的改变都可能引起动物行为的改变。达乌尔黄鼠表现出较低的社会性与其野外种群的生活习性相适应。圈养环境和野生环境相比,活动空间小、种群密度高,取食、寻求庇护所与社会接触的活动以及躲避天敌的机会均减少,因此,圈养动物的生活不同于自然栖息地[39]。同样,实验室内饲养,饲养笼有限的空间也会影响动物行为表达[52-54]。研究发现野外、实验室驯化和实验室繁殖的树鼩(Tupaiabelangeri)在体重、身体组成和消化道形态等方面具有差异,在行为上也出现了一定的分化[55-56]。本实验中即使同在饲养笼饲养,出生于实验室的黄鼠与野外出生的黄鼠相比,一些社会行为特征发生明显改变,这种变化与早期生活环境和社会环境的变化有关。因此,在描述野生动物的行为属性时要考虑到所处环境的影响。
